中国区域滨海湿地固碳能力研究现状与提升
2014-03-20唐博龙江平章伟艳谢永清乔吉果翟万林
唐博,龙江平,2,章伟艳,谢永清,乔吉果,翟万林
(1.国家海洋局第二海洋研究所,浙江 杭州 310012;2.浙江大学,浙江 杭州 310006;3.浙江省水文地质工程地质大队,浙江 宁波 315012;4.天津华北地质勘查局海洋地质勘查中心,天津 300170;5.国家海洋技术中心,天津 300112)
社会经济的发展,使大气中CO2的排放量增加,大气温度升高,海平面上升,给生态环境和沿海经济造成很大危害。有研究表明,100 多年以来,全球大气CO2含量增加了25 % (王云英,2012)。德国波茨坦气候影响研究所的Stefan Rahmsdorf 认为,到2200年,海平面估计将上升1.5~3.5 m。
湿地是全球最大的碳库之一,可以将空气中的CO2固定和封存。全球湿地面积仅占陆地面积的4 %~6%,但它的碳储量占陆地生物碳素的35%,超过农业、森林生态系统碳储量之和,具有很强的固碳能力,固碳潜力较其他类型的生态系统最高(段晓男等,2006)。
滨海湿地是指发育在海岸带附近并且受海陆交互作用影响的湿地,按照国际湿地公约定义,滨海湿地的下限为海平面以下6 m 处(习惯上常把下限定在大型海藻的生长区外缘),上限为大潮线之上与内河流域相连的淡水或半咸水湖沼以及海水上溯未能抵达的入海河的河段(关道明,2012)。滨海湿地是湿地的三大类型(滨海、河流、湖泊) 之一,是生产力最高的天然生态系统。滨海湿地占地面积比较小,但是在固碳方面发挥着重要作用,是许多国家的研究热点。
1 国外滨海湿地的固碳能力研究
湿地生态系统在美国和欧洲研究较多,1971年《湿地公约》颁布后引起了更大的关注,主要是对湿地鸟禽保护、盐沼湿地以及河口湿地的研究,以及研究湿地野生保护、生态作用等方面。近年来,由于全球气温上升,湿地碳循环与湿地固碳受到了广泛关注。
河口三角洲处在水动力较强的沿海区域,大量的悬浮泥沙会携带大量物质,并会在絮凝作用下沉积下来。Brix 等(2001) 对欧洲芦苇湿地的碳固定和碳释放做了研究,发现芦苇湿地在短期内(数十年) 是碳源,但是,在长期内(大于100年) 则会成为碳库。
泥炭地对气候水平衡变化的响应是预测全球碳循环的潜力的重要因素。Belyea 等(2004) 指出泥炭地的形成和固碳率与气候水文、泥炭地的表层结构有很大关系,分析他们之间的关系对于预测全球碳循环潜在的反馈具有重要作用。Bernal 等(2012) 认为湿地类型和植被群落的不同与生长差异会导致湿地的固碳能力有很大变化,清楚的认识湿地类型和湿地植被群落对于研究湿地的固碳能力具有重要意义。Choi 等(2004) 用同位素测年法测量盐沼湿地有较高的碳含量,发现短期内湿地固碳量高于长期的,这可能与有机物长期处在厌氧环境有关,还可能因为近年来气候变化,海平面上升导致土壤层抬高、碳氮含量增加导致初级生产力提高有关。结果表明盐沼湿地是减少温室效应的碳库。Chmura 等(2003) 调查了大西洋、太平洋的东面和西面沿岸,以及印度洋、地中海、墨西哥沿岸的盐碱沼泽湿地和红树林湿地的固碳量,并且研究了他们的碳密度和碳储存随着气候以及地理位置的空间变化。土壤碳埋藏会随着气候,植物种类、区域环境条件变化而变化。温度越高,土壤碳埋藏越多。植物种类会影响植物生长速率和分解速率。潮汐、泥沙含量、海拔的影响是综合因素:海拔高的地方,潮汐大,含沙量高,土壤碳埋藏较少,有些海拔低的地方由于易于沉积物沉降碳埋藏率较高。Howe 等(2009) 研究表明澳大利亚Hunter 河口受到潮流干扰的湿地比没有受到潮流干扰的湿地土壤固碳率高,含碳量低,这与湿地的垂向加积和土壤密度有关。Eric 等(2004) 用同位素测年法测定了美国加利福尼亚南部滨海泻湖和湿地混合区沉积物的固碳率,在过去5 000年里,固碳速率为0.033 ± 0.002 9 kg C /m2/yr。滨海湿地固碳能力研究结果见表1。
综上,研究滨海湿地固碳能力的研究主要从其固定碳和释放碳的角度出发,其次,水文环境也是研究的重点。不同时间尺度内固碳能力也不同,尤其是近年来环境污染严重,与工业革命之前相比会有很大变化,所以,探究不同时间尺度的固碳量和固碳率可以有效揭示影响湿地固碳能力的因素。滨海湿地的固碳潜力很大,研究滨海湿地的固碳能力及其提升技术对于改善生态环境和社会经济的发展有重要意义。
2 我国滨海湿地的固碳能力研究
我国海岸线绵长,滨海湿地总面积约6 万km2,占我国湿地总面积的15.4 % (安鑫龙 等,2009;赵扬,2008)。涉及沿海的11 个省(区、市)。以杭州湾为界,以北由环渤海滨海和江苏滨海湿地组成,以南主要有杭州湾、泉州湾、珠江口河口湾和北部湾湿地等。我国滨海湿地类型可分为浅海水域、滩涂、滨海沼泽、河口水域、河口三角洲以及人工湿地(养殖池塘、盐田、水库) (关道明,2012)。
滨海湿地水陆交界的自然环境为碳储存提供了良好的条件,植物固碳能力比较高。滨海湿地上的植物固定的碳部分储存在土壤中,称为土壤的碳汇。土壤是滨海湿地最大的碳库。目前我国有多位学者对于滨海湿地固碳和土壤碳储量进行了研究,显示了滨海湿地很强的固碳能力。

表1 国外滨海湿地固碳能力研究
我国对滨海湿地植被的固碳能力研究较多,涉及温带到热带地区,跨度较大。
辽河三角洲湿地植物主要是芦苇,稻田,碱蓬滩涂,固碳能力平均值为1.77 kg/m2/yr,固碳能力比较高,是黄河三角洲滨海湿地植被平均固碳能力的5 倍,甚至与同等条件下的温带落叶阔叶林的固碳能力相当(索安宁等,2010)。
长江口崇明东滩的主要植物是海三棱藨草和芦苇。两种植物固碳能力之和为1.47~3.52 kg/m2/yr,是全国陆地植被平均固碳能力的3~7.18 倍,是全球植被平均固碳能力的3.59~8.59 倍(梅雪英等,2007a,2007b)。研究了崇明东滩植被群落的演替和碳储固碳的变化,结果表明植被由海三棱藨草群落向芦苇群落演替的过程中,固碳能力增强了。同时提出非生物组分对植被固碳有影响。还分析了海三棱藨草碳储量、固碳能力、固碳潜力,其固碳潜力是很大的(梅雪英等,2007a,2007b)。
黄河三角洲滨海湿地植被平均固碳能力为0.35 kg/m2/yr(张绪良 等,2012),土壤有机碳含量为0.75~8.35 g/kg(董洪芳等,2013)。天然的湿生湿地植被固碳能力高于全国陆地植被平均固碳能力,但是盐生湿地和水生湿地植被由于干旱和土壤盐渍化,植物固碳能力低于全国陆地植被平均固碳能力,总体上固碳能力不强(张绪良等,2012)。
天津滨海湿地土壤属于盐碱地,有机质容易降解,植物覆盖度低,土壤碳储量不高,郝翠等分析天津滨海湿地土壤平均有机碳密度与全国平均有机碳密度(9.6 kg/m2) 相比小近1.5~4 倍。土壤储碳量由大到小的顺序为林地、草地、滩涂、浅水域、裸土(郝翠等,2011)。
杭州湾是南北滨海湿地的分界,杭州湾南岸主要是淤涨型滩涂,含盐量较高。植物主要种类有芦苇、互花米草和海三棱藨草。邵学新等测得杭州湾土壤有机碳平均含量为6.45±1.70 g/kg,低于天津滨海湿地土壤有机碳含量,固碳能力较弱,这可能与围垦造地有较大关系(邵学新等,2011)。
红树林是滨海湿地典型的植物类别,生长于陆海交界的潮间带,发达的根系可以固岸护堤,防止水土流失,还可以净化海水和空气,对保护沿岸的生态环境有重要作用。我国红树林主要分布在广西、广东、海南、台湾、福建、香港、澳门和浙江南部沿岸,广西、广东、海南覆盖面积最广,约占我国红树林面积的97%,红树林种类也较多。红树林有很强的固碳能力,对于减少全球碳含量,降低温室效应有不可替代的作用。如许方宏等测得我国面积最大的红树林保护区广东湛江红树林自然保护区地表100 cm 桐花树群落、木榄+秋茄群落、白骨壤群落土壤碳库平均为45.68 kg/m2(许方宏等,2012)。据报道,该保护区高桥核心区900 公顷红树林每年可吸收23 100 t 二氧化碳,固定碳627 t(刘垠,2012);曹庆先等(2011) 实验获得北部湾5 种红树林植物群落地上平均生物量为2.257 kg/m2。朱可峰(2007) 获得广州南沙新垦湿地人工红树林无瓣海桑生物量为11.46 kg/m2,同期种植的木榄生物量为5.55 kg/m2,木榄平均固碳速率是0.404 kg/m2/yr,约是全球陆地植被平均固碳能力的1 倍。无瓣海桑平均固碳能力为0.964 kg/m2/yr,是全球陆地植被平均固碳能力的2.4 倍。表2 列出了我国典型滨海湿地的固碳能力情况。
我国各地滨海湿地的固碳能力存在较大的空间差异,北方地区固碳能力高于全球植被平均固碳能力的3~9 倍,南方地区固碳能力低于全球滨海湿地的固碳能力。这主要受植被生长状况和植被覆盖度的影响。其次气候对其固碳能力也有很大影响,北方天气寒冷,生物降解程度低,固定的碳大部分储存在土壤中,固碳能力高,南方天气炎热,生物降解程度较高,净固碳量较低。综合整个中国区域,我国滨海湿地的固碳能力较高。另外,植被的生长状况是影响其固碳能力的直接因素,如生长多年的天然红树林有机碳密度是全国平均有机碳密度的几十倍,但是人工红树林的有机碳密度低于全国平均有机碳密度。
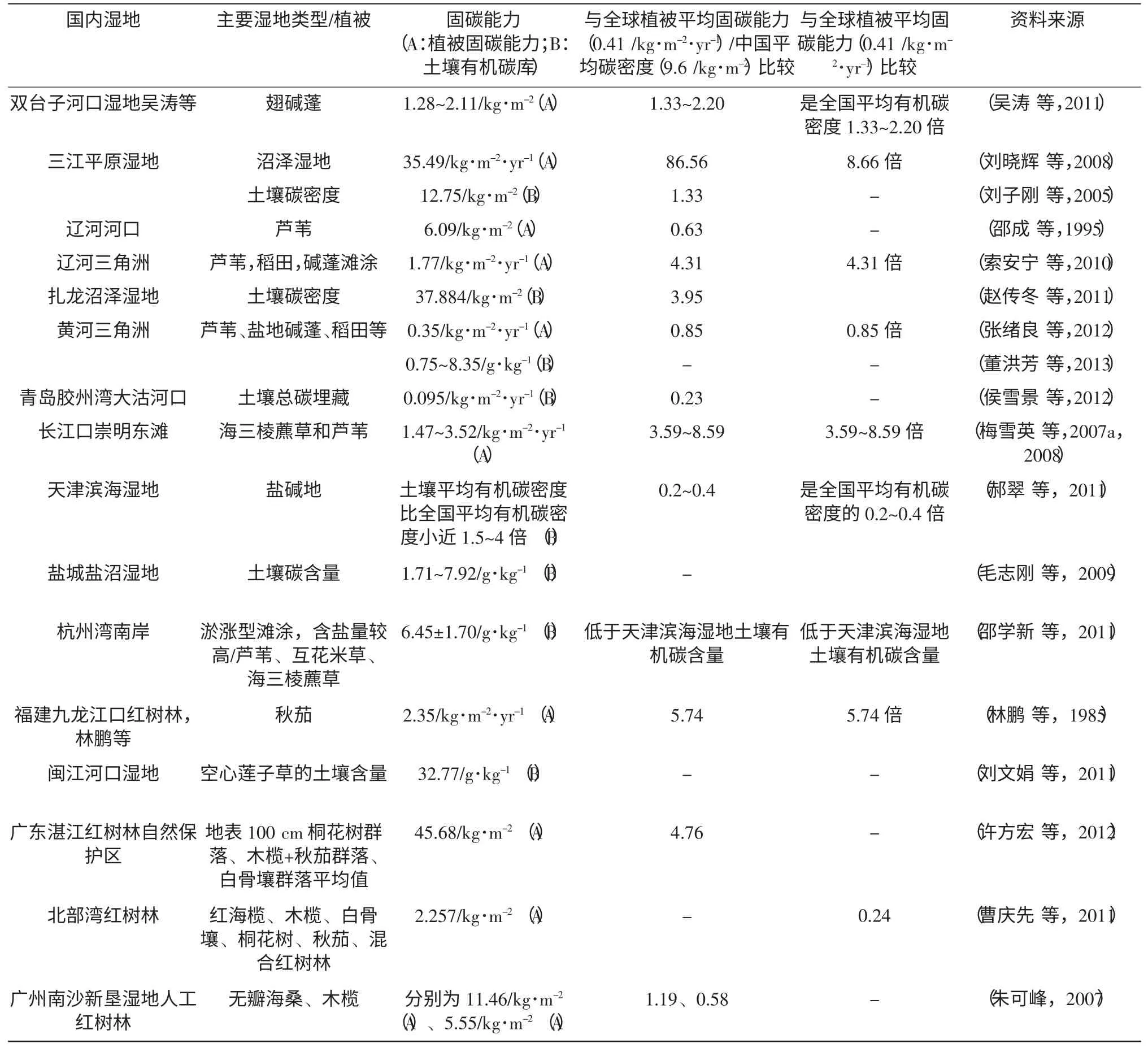
表2 我国典型滨海湿地的固碳能力
3 滨海湿地固碳能力的遥感监测
滨海湿地分布在沿海的海陆交界地带,覆盖面积大小不同,分布分散,地基稀软,不易进行现场调查。遥感技术的出现为滨海湿地调查提供了方便,它可以大面积、实时、全天候的对滨海湿地进行监测,节省人力物力,填补了滨海湿地调查的一大空白。
遥感监测是利用人造卫星、飞机或其他飞行器获得滨海湿地上的地物电磁辐射信息,判别所测地物的特征。卫星发射的光谱根据波段长度的大小可分为可见光、红外线、紫外线、微波。不同波段对地物的响应不同,所以要有针对性的选择光谱波段进行探测,现代技术多为多谱段探测卫星,可以同时获得更多的地物信息。飞机或其他飞行器可以近距离的观测所测地物,能够更准确的辨别地物信息,弥补卫星遥感远距离,低分辨率带来的信息缺乏与信息模糊的不足。通过遥感获得的图像不能直接运用,还要经过校正、信息提取与分类、精度检验等一系列处理,以达到识别地物信息的目的。
但是,遥感监测并不能完全代替现场调查,在图像处理过程中需要结合资料收集和实地调查的结果对遥感图像进行识别。现场调查可以了解湿地环境地貌、植物群落与植被覆盖状况、土壤和水环境要素、保护与利用状况、破坏状况等,这对遥感图像识别有很大帮助。
在20 世纪70年代末,美国政府要求调查录入全国滨海湿地的分布,以期对滨海湿地的生态价值做出评估(Hardisky et al,1986),由此遥感监测在滨海湿地的运用应运而生,遥感监测开始用于滨海湿地的自然环境、分布范围或者滨海湿地植被分布以及滨海湿地的演变等方面(Ramsey,1995;Rao et al,1999;Victor et al,2001)。
随着遥感技术的发展,遥感技术在滨海湿地固碳方面发挥了很大的作用,已被应用于滨海湿地水质、植物生物量、固碳量、固碳能力等多个方面的研究。如利用SRTM 数据建立红树林的平均高度与生物量之间的关系,获得红树林沼泽地国家公园生物量的地理分布图(Simard et al,2006)。利用水体反射的光谱特征和水质参数之间的关系监测水质,为湿地保护提供重要信息(杜为静等,2010);建立植被指数与生物量之间的关系,获得湿地植被的生物量(黎夏等,2006);运用雷达后向散射系数建立湿地植被的生物量估算模型(黎夏 等,2006);通过对比不同时间植被生物覆盖度分析植被的时空变化情况,为湿地保护对策措施的实行提供依据(赵峰等,2011);利用遥感确认湿地的空间分布与面积大小(田素荣等,2007;李学杰等,2010);通过分析不同传感器波谱的特征选取波段对湿地上的要素进行解译和分类,其精度可达到湿地环评的制图要求(曹杨等,2010)。
4 滨海湿地固碳能力评估
4.1 生物量
滨海湿地固碳能力的评估方法可以借助滨海湿地上生长的植物生物量来衡量,植物光合作用固定CO2并放出O2,其方程式(鲁莉,2008) 如下:

由光合作用反应方程式可知植物每产生1 g 干物质需要1.63 g 的CO2。所以由植物生物量可以求得植物固定的CO2量,从而获得植物的固碳能力。
计算植物生物量一般取固定面积的植被,计算地上部分和地下部分或者计算整颗植株的干重量或者鲜重,也可以根据已有的方法进行估算(张鹏超等,2010)。如贾小荣等由林分密度及平均木的器官含水量估算林分的干重(贾小容等,2006);杨同辉等针对植物的不同种类运用了与其相适应的方法计算干重(杨同辉等,2010)。计算生物量法是最直接,最准确,目前普遍运用的方法。
利用建立方程和模型拟合的方法计算植物生物量。刘晓辉等(2008) 在计算沼泽湿地的植物地上生物量的时候采用CHIKUGO 模型近似取植物的净初级生产量为植物生物量。李轩然等(2006) 利用实测值法(即计算植物器官干重)、估计值建模法(建立方程) 和平均标准木法(拟合方程) 3 种方法计算湿地松林生物量并对计算结果进行比较。
利用遥感来反演植物生物量,即建立遥感参数与植被生物量之间的关系测得植被生物量。目前主要有光学遥感和雷达遥感两种方法,核心是拟合植被指数或雷达散射系数与植被生物量之间的关系。黎夏等(2006)通过雷达遥感估算了红树林植被的生物量,并得出雷达散射系数比植被指数在估算植被生物量中精度高的结论。沈国状等(2009) 基于ENVISAT ASAR 数据通过神经网络方法拟合生物量与后向散射系数间的关系获得植被生物量。王庆等(2010) 将光学遥感和雷达遥感结合起来测植被生物量,弥补了两种方法的不足,有效提高了生物反演精度。
4.2 碳通量
沉积物碳通量是指生态系统单位时间单位面积通过沉积断面的碳含量。沉积物碳通量可以表征沉积物的储碳速率和储碳能力。通过断面净碳通量越多,生态系统固碳能力越强(邹绿柳等,2009)。测量碳通量的方法主要有箱式法和涡度相关法。
箱式法测量的空间是密封的,空气不能流通,排放通量值有偏差。涡度相关法能够测量平流通量,并能长期监测生态系统碳通量,是目前测量生态系统碳通量的主要方法(宋霞等,2003;耿绍波等,2010)。但是涡度相关法受下垫面地形和气象的影响,会产生一定误差,但可以监测长期的沉积物碳汇变化,给观测湿地碳循环和碳沉积提供很大帮助。
4.3 土壤碳含量和碳密度
滨海湿地土壤是碳储存的空间,可以称为碳库。土壤碳含量和碳密度在一定程度上可以反映滨海湿地的固碳能力。
土壤碳含量指单位质量的沉积物中所含的碳含量,计算公式(董洪芳等,2013) 为:

SOC 为土壤有机碳含量(g/kg),FOC 为土壤物理组分有机碳含量(g/kg),P 为土壤有机碳物理组分分配比例。
土壤碳含量还可以用实验室分析法直接测量,分别测得土壤有机碳含量(g) 和土壤质量(kg),两者相除得到(邵学新等,2011)。
土壤碳密度指单位面积一定深度的沉积物层中的碳储量(Hardisky et al,1986),计算公式(蔡毅 等,2013) 为:

式中:Ci为土壤碳含量(g/kg),Di为土壤容重(g/cm3),Ei为土壤层厚度(cm)。土壤碳密度单位为kg/m2或t/hm2。土壤碳含量、容重、厚度用仪器测量或实验分析。其他计算土壤碳密度的公式可以在此公式基础上根据实际情况进行变换。
我国多位学者对我国的碳含量和碳密度进行了评估,具体评估结果见表2。
4.4 碳埋藏速率
大气碳流量通过光合作用被固定,并通过分解作用释放到大气中(Kristin et al,2006),这其中固定碳与分解碳的差即为碳埋藏。碳固定速率越快,分解速率越低且湿地面积越大滨海湿地的固碳潜力越大。碳固定速率在现场由仪器测得;碳分解过程包括碳流失和碳释放两个过程,有氧环境下释放CO2,厌氧环境下释放CH4。碳流失率可以实地观测,使用“现场围隔比较法”等方法测量,碳释放率在现场通过仪器测得。由此可获得湿地碳埋藏的速率(段晓男等,2008):

CSR 为碳固定速率mg/(cm2·a),CLR 为碳流失速率mg/(cm2·a),CER 为CO2释放速率mg/(cm2·a)。
滨海湿地的碳埋藏在沉积物中,埋藏速率与沉积物容重、沉积物碳含量、沉积物沉积速率成正比,有以下公式(段晓男等,2008):

P 为湿地沉积物容重(g/cm3);SOC 为湿地沉积物碳含量(g/kg);R 为湿地沉积物沉积速率(mm/a)。沉积物的沉积速率一般用同位素定年法测量,指利用天然放射性同位素的衰变规律计算出沉积物的年龄,并由此计算出沉积速率。常用的同位素有14C、210P 和137Cr 等。
我国滨海湿地中红树林湿地固碳速率最快,其次是沿海盐沼和泥炭地(段晓男等,2008)。这与红树林湿地植被特性和沉积物分解速率慢有关,其次,可能是因为红树林多生长在温热带地区,生产力相对较高;沿海盐沼固碳速率快是因为沿海沼泽湿地有浅层水覆盖,处于缺氧环境,碳分解慢,还会形成泥炭,导致碳分解速率更慢。由于缺氧环境植被的根系多露在水面,根系裸露削弱了波浪海潮等的流速,挟带而来的沉积物沉积下来,加快了沉积速率;泥炭地主要分布在寒冷地区,植被碎屑被埋藏在土壤中,在低温、少雨、缺氧的环境下分解非常缓慢,使生产力不高的泥炭地不断积累碳。
4.5 结论
同一地区运用不同的方法可能会造成结果差异,以能准确方便获得测量分析数据为主要依据,结合研究区的特点选择合适的评价方法。实验室分析生物量法测量简便,但是由于现实中不能把所有的树砍下来拿回实验室分析,导致分析结果有片面性,研究大区域植被的生物量精度不高。涡度相关法可以获得长期监测数据,需要实地测量数据相配合,设备建设难度大,成本高,测量数据的稳定性和准确性较低(宋洪涛等,2011)。沉积物是储存碳的场所,湿地植被将气体CO2封存在沉积物中,完成固碳过程,滨海湿地的沉积物碳密度和碳埋藏速率是针对沉积物做的结果性研究,,此过程主要受气候、植被、水体、沉积物影响,在分析滨海湿地沉积物碳密度和碳埋藏速率时要考虑它们产生的影响。
5 滨海湿地固碳能力的提升技术
滨海湿地由于人类干扰,受到很大破坏,环境恶化,面积减少。20 世纪80年代以来我国滨海湿地面积丧失了近50%。经济发展,人类生活干扰,填海造地工程、造塘养殖等使滨海湿地面积大大减少。近年来气候的改变值得人们反思,人们也已经认识到保护滨海湿地的重要性和必要性。研究如何保护滨海湿地,对环境保护和人类生活有重要意义。
滨海湿地的固碳能力的提升包括生物物种选择与结合种植,生境改造技术与种植技术和管理技术(提高存活率和固碳效率)。
为了提高生物对生态资源的利用,采用乔木、灌木和草本植物混交群落的高效固碳种植方法。另外,固定面积土地的植物不是越多越好,植物数量太大,会造成营养供应不足,空间和阳光的竞争激烈等问题。所以,要合理选择生物物种,科学结合生物种植。植被密度要合理,太稀疏固碳率不高,太密不通透,放出的CO2多于固定的CO2,起到反效果。如叶功富等(2008) 研究表明木麻黄-厚荚相思混交林的土壤层含碳率和碳贮量大于10年生木麻黄纯林的,可见混交林有利于碳的积累和保存。
滨海湿地的生境是指生态环境,生态环境对滨海湿地生物有很大影响,反之,生物也会影响生态环境。不同种类的生物对环境的适应性不同,选择能适应当地环境的植物进行栽培种植可以提高存活率。同时,人类活动是破坏滨海环境的主要因素,保护滨海生态环境,可以有效降低植物死亡率,尤其是旅游区这些敏感地区。修复滨海湿地成为了不可避免的话题,涌出了多种修复技术方法。
窦勇等(2012) 结合国内外技术方法总结分析了利用微生物修复技术和植物修复技术修复滨海湿地。微生物修复技术是利用微生物对特定污染物的分解作用净化滨海湿地。目前,利用微生物分解石油烃类和重金属等技术比较成熟。植物修复主要是利用种类植物对元素的吸收特性,针对区域污染状况,选择对污染物吸收强的植物进行栽培,以改善当地的生态环境。
提升滨海湿地固碳能力改善土壤环境是很重要的措施。目前有人尝试在土壤中加入土壤改良剂来改良土壤性质,提高土壤营养含量,提高土壤固碳能力。尤其是生物改良剂是研究热点。生物改良剂是在土壤中加入菌类、蚯蚓等生物来改善土壤的物理性质和化学性质(陈义群等,2008)。
提高植物存活率和固碳效率跟种植技术和管理技术密不可分,滨海湿地生态环境特殊,地质稀软,潮汐冲刷,给种植和管理人员带来不便,需要专业技术进行种植和管理。如杨淑慧等(2011) 研究发现不同刈割期和还田处理对土壤碳排放有很大影响。
滨海湿地土壤即是碳库,在受到干扰时还会成为碳源,湿地储存着大量的碳,如果作为碳源释放危害很大,所以,提高滨海湿地的固碳能力,首先要保护湿地不被破坏,防止湿地变为碳源。滨海湿地的破坏主要是围垦造地、生活污水、工业废水排放、养殖业等带来的,围垦造地是彻底的,不能逆转的,并会进一步破坏周边环境。政府要加大立法,加强政策法规的实施,相关部门要严格遵守法律,并加强宣传作用,增强人们保护湿地的意识。与此同时,要扩大湿地面积,20 世纪80年代至今,我国湿地面积减少近50%。红树林湿地面积在20 世纪50年代近5 万公顷,到2002年全国红树林面积仅剩近3 万公顷(张忠华等,2006;王燕,2008)。政府对湿地的保护和面积扩展能力是有限的,利用经济利益,让当地居民保护和扩展湿地面积往往能取得较好的效果。目前,美国等一些发达国家实施谁污染谁治理的政策,对于排放污染物的集团或个人罚款,并用于环境治理(Zedler,2000),我国可以借鉴国外的做法,加强法律规范和处罚强度。
另外,有些地区选择引进外来物种来改善湿地的固碳能力。引进外来物种可能会增强已有种群的优势,改善当地土壤和水文环境,能合理地抑制有机碳无机化种群的数量。但是,对于外来物种往往会因为环境的改变而成为优势种,对本地物种造成威胁,甚至消失,比如西葫芦,芦苇等。所以,这种方法要在实验证据充足的条件下使用。如护花米草入侵福建漳江口地区红树林,薇甘菊入侵深圳红树林,在这两个地区红树林都因为外来植被的入侵资源被占用,生长环境被破坏(毛子龙 等,2011)。
6 总结
由于人们对海岸的依赖与破坏,滨海湿地退化现象严重。滨海湿地由于其特殊的自然环境有很强的固碳能力,以小面积高固碳率成为降低全球温室效应的重要生态系统。滨海湿地固碳作用研究起步较晚,在现场调查和遥感监测方面还存在很多不足,如现场仪器的建设难、精度低、人工测量的环境阻碍等;遥感监测图像处理方法带来的分类误差等。所以,需要研究更好的仪器设备适应滨海湿地多变的自然环境。遥感卫星技术也需要人们继续探索提高。
我国属于沿海国家,岸线绵长,滨海湿地面积较大,并且主要集中在南方的温热带地区。我国总体上北方由于处在低温状态下,土壤中的碳很难分解,碳可以不断积累,所以碳库比南方高;长江口由于自然环境优良,植被生长状况良好,植被固碳能力较我国其他地区高;红树林有特殊的生长环境,受潮流影响明显,且土壤处于缺氧环境,碳排放率很低,形成很好的碳库,且植被固碳能力与长江口植被固碳能力相当,但人工湿地年龄小,固碳能力和碳库都较低(表2)。可见,自然环境,植被类型,气候是影响滨海湿地固碳能力的主要因素。
滨海湿地不仅固碳能力高,固碳潜力也很大,提高固碳潜力要重点研究植被物种选择和种植、提升沉积物碳埋藏和降低碳排放。管理技术和保护工作也很重要,研究表明被破坏的湿地恢复是很漫长的过程,可能需要近100年的时间(Burden et al,2013)。如何提高滨海湿地的固碳能力,增加湿地固碳,减少湿地碳排放将成为降低温室效应的重点问题。
Belyea L R, Malmer N, 2004. Carbon sequestration in peatland: patterns and mechanisms of response to climate change.Change Biology,10:1043-1052.
Bernal B, William J M, 2012. Comparing carbon sequestration in temperate freshwater wetland communities. Global Change Biology,18:1636-1647.
Brix H,Brian K S,Lorenzen B,2001.Are Phragmites dominated wetlands a net source or net sink of greenhouse gases. Aquatic Botany, 69:313-324.
Burden A,Garbutt R A,Evans C D,et al,2013.Carbon sequestration and biogeochemical cycling in a saltmarsh subject to coastal managed realignment.Estuarine,Coastal and Shelf Science,120:12-20.
Cannell M G R,Milne R,Hargreaves,et al,1999.National inventories of terrestrial carbon sources and sinks: the UK Experience. Climate Change,42:505-530.
Chmura G L, Anisfeld S C, Cahoon D R, et al, 2003. Global carbon sequestration in tidal, saline wetland soils. Global Biogeochemical Cycles,17(4):22(1)-22(12).
Choi Y, Wang Y, 2004. Dynamics of carbon sequestration in a coastal wetland using radiocarbon measurements. Lobal Biogeochemical Cycles,Vol.18,GB4016,2004:1-12.
Day J W, Conner W H, Ley-Lou F, et al, 1987. The productivity and composition of mangrove forests, Laguna de Terminos, Mexico.Aquatic Botany,27:267-284.
Eric C B, Jeffrey A H, 2004. A 5000 year record of carbon sequestration from a coastal lagoon and wetland complex, Southern California,USA.Catena,57:221-232.
Hardisky M A, Gross M F, Klemas V, 1986. Remote Sensing of Coastal Wetlands.BioScience,36(7):453-460.
Hensel P F,Day Jr J W,Pont D,1999.Wetland vertical accretion and soil elevation change in the Rhone River Delta, France: The importance of riverine flooding.J.Coastal Res,15:668-681.
Howe A J, Rodríguez J F, Saco P M, 2009. Surface evolution and carbon sequestration in disturbed and undisturbed wetland soils of the Hunter estuary, southeast Australia. Estuarine, Coastal and Shelf Science 84:75-83.
Juman R A, 2005. The structure and productivity of the Thalassia testudinumcommunity in Bon Accord Lagoon, Tobago. Revista de Biologia Tropical,53:219-227.
Kristin R,Newton P,Simon E,et al,2006.2006年IPCC 国家温室气体清单指南(V4).日本:日本全球环境战略研究所.
Ramsey E W, 1995. Monitoring flooding in coastal wetlands by using radar imagery and ground-based measurements.International Journal of Remote Sensing,16,Issue 13:2495-2502.
Rao B R M, Dwivedi R S, Kushwaha S P S, et al, 1999. Monitoring the spatial extent of coastal wetlands using ERS-1 SAR data.Int.J.Remote sensing,20(13):2509-2517.
Sharon M L E,Evelyn E G,Daniel L C,et al,2006.Spatial and temporal patterns of aboveground net primary productivity (ANPP)along two freshwater-estuarine transects in the Florida Coastal Everglades.Hydrobiologia,569:459-474.
Simard M, Zhang K, Victor H R M, et al, 2006. Mapping Height and Biomass of Mangrove Forests in Everglades National Park with SRTM Elevation Data. Photogrammetric Engineering & Remote Sensing,72(3):299-311.
Victor V, Klemas S, 2001. Remote Sensing of Landscape-Level Coastal Environmental Indicators. Environmental Management, Vol. 27, Issue 1:47-57.
Vymazal J,1995.Algae and Elemental Cycling in Wetlands. Lewis Publishers,Boca Raton,FL.
Zedler J B, 2000. Progress in wetland restoration ecology. Tree, 15(10):402-407.
安鑫龙,齐遵利,李雪梅,等,2009.中国海岸带研究Ⅲ--滨海湿地研究.安徽农业科学,37(4):1712-1713.
蔡毅,景艳波,钟志诚,等,2013.深圳湾红树林土壤碳同位素研究.价值工程,(19):317-320.
曹庆先,徐大平,鞠洪波,2011.北部湾沿海5 种红树林群落生物量的遥感估算.广西科学,18(3):289-293.
曹杨,尚士友,杨景荣,等,2010.多源遥感数据在湖泊湿地信息提取中的应用.人民黄河,32(2):5-7,9.
陈义群,董元华,2008.土壤改良剂的研究与应用进展.生态环境,17(3):1282-1289.
董洪芳,于君宝,管博,等,2013.黄河三角洲碱蓬湿地土壤有机碳及其组分分布特征.环境科学,34(1):287-292.
窦勇,唐学玺,王悠,2012.滨海湿地生态修复研究进展.海洋环境科学,31(4):616-620.
杜为静,李淑敏,李红,等,2010.汉石桥湿地水质参数光谱分析与遥感反演.光谱学与光谱分析,30(3):757-761.
段晓男,王效科,逯非,等,2008.中国湿地生态系统固碳现状和潜力.生态学报,28(2):463-469.
段晓男,王效科,尹弢,等,2006.湿地生态系统固碳潜力研究进展.生态环境,15(5):1091-1095.
耿绍波,鲁绍伟,饶良懿,2010.基于涡度相关技术测算地表碳通量研究进展.世界林业研究,23(3):24-28.
关道明,2012.中国滨海湿地.北京:海洋出版社:3.
郝翠,李洪远,李姝娟,等,2011.天津滨海湿地土壤有机碳储量及其影响因素分析.环境科学研究,24(11):1276-1282.
侯雪景,印萍,丁旋,等,2012.青岛胶州湾大沽河口滨海湿地的碳埋藏能力.海洋地质前沿,28(11):17-26.
贾小容,曾曙才,苏志尧,2006.广州白云山9 种林分光合固碳放氧价值核算.广东林业科技,22(2):19-21.
黎夏,叶嘉安,王树功,等,2006.红树林湿地植被生物量的雷达遥感估算.遥感学报,10(3):387-396.
李轩然,刘琪璟,胡理乐,等,2006.不同方法计算湿地松林生物量的比较.生态学杂志,25(12):1594-1598.
李学杰,万荣胜,林进清,2010.应用遥感方法分析北部湾滨海湿地的分布.南海地质研究:28-36.
林鹏,卢昌义,林光辉,等,1985.九龙江口红树林研究——Ⅰ.秋茄群落的生物量和生产力.厦门大学学报(自然科学版),24(4):508-514.
刘文娟,曾陆金,王维奇,等,2011.闽江河口湿地空心莲子草土壤碳库研究.7(1):51-55.
刘晓辉,吕宪国,2008.三江平原湿地生态系统固碳功能及其价值评估.6(2):212-217.
刘垠,2012.红树林湿地减缓气候变暖进程.科技日报,12-04(003).
刘子刚,张坤民,2005.黑龙江省三江平原湿地土壤碳储量变化.清华大学学报(自然科学版),45(6):788-791.
鲁莉, 2008. 森林固定大气CO2 实物量方法的比较. 甘肃科技, 24(15):53-55.
毛志刚,王国祥,刘金娥,等,2009.盐城海滨湿地盐沼植被对土壤碳氮分布特征的影响.应用生态学报,20(2):293-297.
毛子龙,赖梅东,赵振业,等,2011.薇甘菊入侵对深圳湾红树林生态系统碳储量的影响.生态环境学报,20(12):1813-1818.
梅雪英,张修峰,2007a.崇明东滩湿地自然植被演替过程中储碳及固碳功能变化.应用生态学报,18(4):933-936.
梅雪英,张修峰,2007b.长江口湿地海三棱蔗草(Scirpus mariqueter)的储碳、固碳功能研究--以崇明东滩为例.农业环境科学学报,26(1):360-363.
梅雪英,张修峰,2008.长江典型湿地植被储碳、固碳功能研究--以崇明东滩芦苇带为例.应用生态学报,16(2):269-272.
邵成,陈中林,董厚德,等,1995.辽河河口湿地芦苇的生长及生物量研究.辽宁大学学报,22(1):81-94.
邵学新,杨文英,吴摇明,等,2011.杭州湾滨海湿地土壤有机碳含量及其分布格局.应用生态学报,22(3):658-664.
沈国状,廖静娟,郭华东,等,2009.基于Envisat Asar 数据的鄱阳湖湿地生物量反演研究.高技术通讯,19(6):644-649.
宋洪涛,崔丽娟,栾军伟,等,2011.湿地固碳功能与潜力.世界林业研究,24(6):6-11.
宋霞,刘允芬,徐小锋,2003.箱法和涡度相关法测碳通量的比较研究.江西科学,21(3):206-210.
索安宁,赵冬至,张丰收,2010.我国北方河口湿地植被储碳、固碳功能研究——以辽河三角洲盘锦地区为例. 海洋学研究, 28(3):67-71.
田素荣,孙永军,李友纲,等,2007.多时相遥感技术在湿地调查中的应用.国土资源遥感,(4):81-84.
王庆,廖静娟,2010. 基于Landsat TM 和Envisat Asar 数据的鄱阳湖湿地植被生物量的反演.地球信息科学学报,12(2):282-291.
王燕,2008.红树林保护的意义和建议.森林工程,24(4):19-20,57.
王云英,2012.浅析温室效应对环境的影响及对策.科技创新导报,(1):125.
吴涛, 赵冬至, 康建成, 等, 2011. 辽东湾双台子河口湿地翅碱蓬(Suaeda salsa)生物量遥感反演研究.生态环境学报,20(1):24-29.
许方宏,张进平,张倩媚,等,2012.广东湛江高桥三个天然红树林的土壤碳库.价值工程,(15):5-6.
杨淑慧,任文玲,仲启铖,等,2011.不同管理措施对长江口围垦湿地芦苇群落土壤呼吸的影响.浙江农业学报,23(6):1203-1209.
杨同辉,宋坤,达良俊,等,2010.中国东部木荷--米櫧林的生物量和地上初级生产力.中国科学:生命科学,40(7):610-619.
叶功富,郭瑞红,卢昌义,等,2008.木麻黄与厚荚相思混交林的土壤碳贮量动态.海峡科学,(10):21-23.
张鹏超,张一平,杨国平,等,2010.哀牢山亚热带常绿阔叶林乔木碳储量及固碳增量.生态学杂志,29(6):1047-1053.
张绪良,张朝晖,徐宗军,等,2012.黄河三角洲滨海湿地植被的碳储量和固碳能力.安全与环境学报,12(6):145-149.
张绪良,张朝晖,徐宗军,等,2012.黄河三角洲滨海湿地植被的碳储量和固碳能力.安全与环境学报,12(6):145-149.
张忠华,胡刚,梁士楚,2006.我国红树林的分布现状、保护及生态价值.生物学通报,41(4):9-11.
赵传冬,刘国栋,杨柯,等,2011.黑龙江省扎龙湿地及其周边地区土壤碳储量估算与1986年以来的变化趋势研究.地学前缘(中国地质大学(北京);北京大学),18(6):27-33.
赵峰,张怀清,刘华,等,2011.福建漳江口红树林湿地保护区遥感监测及保护分析.西北林学院学报,26(1):160-165.
赵扬,2008.滨海湿地生态系统服务功能价值评估--以汕头市为例.广州:中山大学:1-3.
朱可峰,2007.广州南沙新垦湿地人工红树林生物量及碳储量研究.广东:华南农业大学:44-50.
邹绿柳,孙谷畴,赵平,等,2009.马占相思人工林净二氧化碳交换和碳同位素通量.应用生态学报,(11):2594-2602.
