甘蓝无蜡粉亮绿突变体材料LD10遗传规律及分子标记研究
2014-03-15
(中国农业科学院蔬菜花卉研究所,北京 100081)
甘蓝无蜡粉亮绿突变体材料LD10遗传规律及分子标记研究
刘东明 杨丽梅*孙培田 李景涛唐 俊 刘泽洲 方智远 刘玉梅 庄 木 张扬勇
(中国农业科学院蔬菜花卉研究所,北京 100081)
为了明确结球甘蓝无蜡粉亮绿突变体材料LD10的特征特性及遗传规律,测定了LD10的主要农艺性状和营养物质含量;对LD10和普通材料21-3的叶面超微结构进行了观察比较;以LD10和21-3为亲本构建6世代群体,对LD10无蜡粉亮绿性状遗传规律进行分析。结果表明:无蜡粉亮绿突变体材料LD10叶面蜡粉呈小颗粒状结构,缺失严重,而普通材料21-3叶面蜡粉呈长棒状结构,含量丰富,二者具有明显差异;LD10的无蜡粉亮绿性状由1个隐性基因(cgl-4)控制,其与SSR标记LT-SSR16紧密连锁,遗传距离为4.7 cM。用LD10与另一单基因隐性遗传的无蜡粉亮绿突变体材料10Q-961杂交获得F1群体,F1群体单株全部表现为叶面有蜡粉,推断LD10与10Q-961的无蜡粉亮绿基因位于不同的遗传位点。
结球甘蓝;无蜡粉亮绿;特征特性;遗传规律;分子标记
结球甘蓝(Brassica oleracea L. var. capitata L.)起源于地中海沿岸,属十字花科芸薹属,具有营养物质含量丰富、适应性广、易栽培、耐运输等特点,我国年栽培面积约90万hm2(方智远,2008;杨丽梅 等,2011),在蔬菜周年供应和出口贸易中具有重要地位。普通结球甘蓝外叶、外层球叶及茎表皮均覆有一层蜡粉,叶片表现为绿色、灰绿色或深绿色。无蜡粉亮绿甘蓝是普通甘蓝的突变体,在其整个生长阶段植株体表无蜡粉覆盖,颜色亮绿,与普通甘蓝存在着显著区别。初莲香和王余文(1993)研究发现无蜡粉亮绿甘蓝具有叶片脆嫩,糖、VC及干物质等营养成分含量丰富的特点;牟香丽等(2013)通过扫描电镜观察发现甘蓝无蜡粉突变体材料叶面蜡粉含量极少,且蜡粉发育不完全;此外,无蜡粉亮绿甘蓝对小菜蛾还具有一定的抗性(Lin et al.,1984;Eigenbrode & Shelton,1990;Eigenbrode et al.,1991;Stoner,1992)。因此,无蜡粉亮绿突变体材料的研究对选育商品性状美观、营养品质优良的甘蓝新品种具有十分重要的意义。
Anstey和Moore(1954)、North和Priestley(1962)、赵建锋等(2006)、曹阳等(2008)、冯辉等(2010)和李景涛等(2012)分别研究了青花菜、抱子甘蓝、菜薹、普通白菜、青梗菜和结球甘蓝无蜡粉亮绿性状的遗传方式,均证明其符合单基因隐性遗传规律。Mo等(1992)和Pu等(2013)分别报道了编号为W-6和6-1025的甘蓝型油菜无蜡粉性状受1对完全显性基因控制。周熙荣等(1995)报道甘蓝型油菜无蜡粉亮绿性状呈隐性遗传,受2对基因控制。从前人的研究结果可以看出,芸薹属作物无蜡粉性状遗传规律较为复杂,现已报道的大多数突变体无蜡粉性状受单个隐性基因控制,单基因显性或多基因遗传的遗传方式较少。
关于植物表皮蜡质基因的研究,目前已从拟南芥(Bernard & Joubes,2013)、水稻(Mao et al.,2012;Zhou et al.,2013)、玉米( Dietrich et al.,2005;Sturaro et al.,2005)、大麦(Richardson et al.,2007)、小麦(Muria et al.,1999)、小盐芥(赵宝,2007)等植物上克隆了一些蜡质相关基因,为阐明植物外表皮蜡质合成、运输及调控机制奠定了基础。李景涛等(2012)、牟香丽等(2013)、Pu等(2013)、Zhang等(2013)对结球甘蓝、甘蓝型油菜、大白菜等芸薹属植物无蜡粉亮绿性状分子标记及基因定位进行了研究,获得一些与无蜡粉性状控制基因紧密连锁的分子标记。但是这些标记与甘蓝无蜡粉性状控制基因间的遗传距离仍较远,需要进一步寻找与无蜡粉基因连锁更加紧密的分子标记,为无蜡粉基因的精确定位和克隆奠定基础。
中国农业科学院蔬菜花卉研究所甘蓝课题研究组已搜集获得6份无蜡粉亮绿甘蓝材料,编号分别为10Q-385、10Q-961、10Q-974、LD10、21-3油和大绿,其中10Q-385和21-3油表现为扁球型,其余4份材料均为圆球型;10Q-385的熟性为晚熟,其余5份材料为早熟或中熟。本试验以圆球型中熟材料LD10为试材,对其主要农艺性状和营养物质含量进行了测定,通过扫描电镜观察比较了无蜡粉亮绿突变体材料LD10和普通材料21-3的叶表面超微结构,从农艺性状和微观层面进一步了解了无蜡粉亮绿突变体材料LD10的特征特性;通过构建6世代群体和遗传规律分析,明确了LD10无蜡粉亮绿性状的遗传规律;通过引物筛选,获得1个与LD10无蜡粉亮绿性状紧密连锁的SSR标记,为无蜡粉亮绿基因的进一步定位与克隆提供了可利用的分子标记,从而加快结球甘蓝无蜡粉亮绿性状种质的研究和利用。
1 材料与方法
1.1 试验材料
试验于2011~2013年在本所南口农场及甘蓝课题研究组实验室进行。以LD10和21-3为亲本构建6世代群体;用10Q-961和LD10杂交获得另一个F1群体。其中LD10由本所甘蓝课题研究组于2010年搜集得到,表现为植株体表无蜡粉,圆球形,开展度中等,外叶数13片左右,中熟;21-3为本所甘蓝课题研究组自有高代自交系材料,植株体表有蜡粉,扁球形,开展度较大,外叶数14片左右,中熟;10Q-961由北京华耐种子有限公司提供,植株体表无蜡粉,圆球形,开展度中等,外叶较直立,外叶数13片左右,生长势较强,其遗传规律为单基因隐性遗传(李景涛 等,2012)。
1.2 试验方法
1.2.1 群体构建及农艺性状调查 2011年7月初播种LD10和21-3的种子;2012年3~6月进行温室人工授粉,并获得F1群体种子;2012年7月初播种F1、双亲及10Q-961的种子,8月下旬定植;于成熟期对LD10植株农艺性状和VC、总糖等营养物质含量进行调查、测定,调查、测定项目包括单球质量、球径、中心柱长、球形、熟性和干物质、总糖、蛋白、VC、粗纤维含量,其中农艺性状的测量值取21株单株的平均值,营养物质含量的测定采用取3个成熟叶球后混样测定的方法。2013年3~6月进行温室人工授粉,获得F2、BC1、BC2群体种子;以LD10和10Q-961杂交获得另一个F1群体种子,用于判断两材料无蜡粉性状控制基因间的关系。2013年7月初播种各群体种子,于3~4叶期调查、记录各群体中有、无蜡粉植株数据,采用SAS统计分析软件进行卡平方测验。
1.2.2 扫描电镜观察 于结球期取无蜡粉亮绿突变体材料LD10和普通材料21-3最内层外叶,将新鲜叶片冲洗干净,避开叶脉切成5 mm×10 mm的长方形小块,2%锇酸熏蒸24 h,自然风干24 h,然后将样品用银胶固定在样品台上,喷金,置于日立S-570扫描电镜下进行观察,放大倍数为3 000倍。
1.2.3 DNA池构建 采用CTAB法(谢景梅 等,2006)提取幼嫩叶片DNA,利用1.0%琼脂糖凝胶电泳检测其完整性和纯度,用紫外分光光度计检测DNA质量和浓度。采用BSA法(白凤虎 等,2006),在F2群体中分别随机选取8株无蜡粉亮绿单株和8株有蜡粉单株,将其DNA分别等量混合,构建无蜡粉单株DNA混池和有蜡粉单株DNA混池。
1.2.4 SSR分子标记分析 利用混池对引物进行筛选,挑选在两池间表现出差异性条带的引物,对差异性引物再用双亲、F1和F2群体单株进行验证。PCR反应体系为10 μL:模板2.0 μL、Taq DNA聚合酶(5 U·μL-1)0.2 μL、10×PCR buffer(25 mmol·L-1MgCl2)1.0 μL、dNTPs(2.5 mmol·L-1)0.8 μL、Forward Primer(5 pmol·L-1)0.4 μL、Reverse Primer(5 pmol·L-1)0.4 μL、ddH2O5.2 μL。反应程序为:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,循环35次;72 ℃延伸7 min,扩增产物4 ℃保存。8%聚丙烯酰胺凝胶电泳分离扩增产物,设恒定电压100 W,银染法染色(许绍斌 等,2002)。
1.2.5 数据记录与连锁分析 将筛选出的SSR标记在双亲及F2群体中进行验证。F2群体中与无蜡粉亮绿亲本LD10带型一致的记为a,与有蜡粉亲本21-3带型一致的记为b,与F1带型一致的记为h,未扩增出条带或条带模糊不清的记为-。利用Joinmap 4.0软件进行连锁分析,绘制连锁图。
2 结果与分析
2.1 LD10叶表皮微观结构
如图1所示,在放大3 000倍视野下,无蜡粉亮绿突变体材料LD10叶面光滑,下表皮几乎没有蜡粉,上表皮只有极少量蜡粉覆盖,且蜡粉呈小颗粒状结构;普通材料21-3叶面蜡粉含量丰富、分布致密,上、下表皮均有大量蜡粉覆盖,且蜡粉呈长棒状结构,两者之间具有明显差异。
2.2 LD10特征特性
农艺性状调查结果显示,无蜡粉亮绿突变体材料LD10生育期80 d左右,叶色亮绿,圆球形,横径14.82 cm,纵径12.65 cm,开展度中等(47.86 cm),中心柱长6.51 cm,外叶数13片左右,单球质量0.55 kg,表现出较好的商品性。
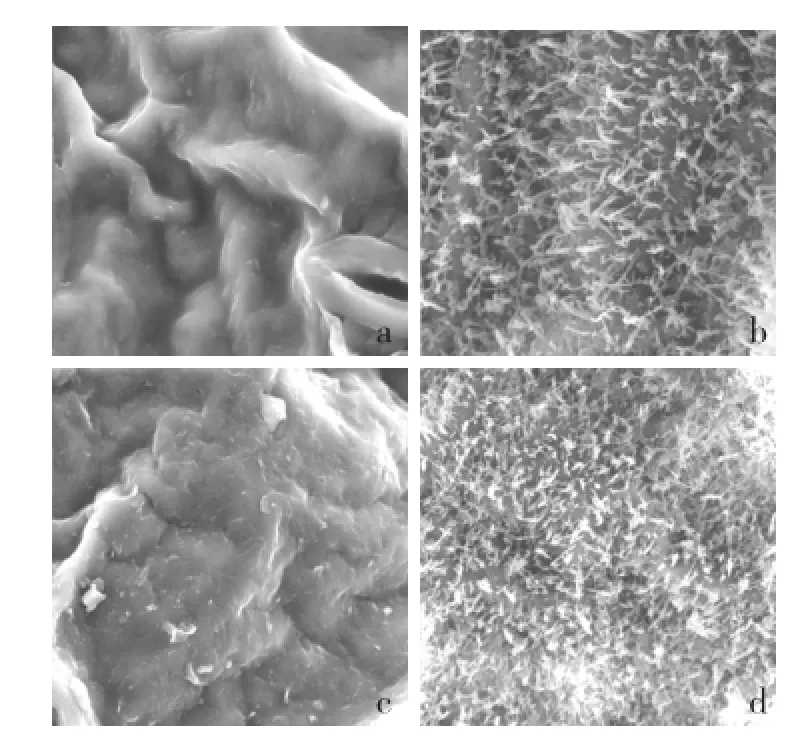
图1 LD10和21-3叶表皮微观结构 (×3 000)
营养物质含量测定结果表明,无蜡粉亮绿突变体材料LD10干物质含量7.82%、总糖3.39%、VC 524.00 mg·kg-1、蛋白1.87%、粗纤维0.64%,营养物质含量丰富,对选育商品性状美观、营养品质优良的甘蓝新品种具有重要意义。
2.3 无蜡粉亮绿性状遗传规律分析

图2 LD10、21-3及其F1叶片微观结构 (×3 000)

图3 LD10、21-3及其F1田间性状
以LD10和21-3为亲本构建6世代群体,F1群体16株单株全部表现为叶面有蜡粉(图2、3);F2群体有蜡粉植株667株,无蜡粉植株231株,分离比例为2.89∶1,经卡平方测验符合3∶1(χ2=0.831<=3.841)的分离比例;以有蜡粉的21-3为回交亲本获得的回交群体中,180株单株全部表现为叶面有蜡粉;而与无蜡粉的亲本LD10回交获得的回交群体中,有蜡粉植株161株,无蜡粉植株157株,分离比例为1.03∶1,经卡平方测验符合1∶1(χ2=0.057<=3.841)的分离比例(表1)。表明结球甘蓝LD10的无蜡粉亮绿性状由隐性单基因控制。
本所甘蓝课题研究组之前已研究报道了另一份无蜡粉亮绿突变体材料10Q-961,并确定其遗传方式符合单基因隐性遗传规律(李景涛 等,2012)。为了研究LD10与10Q-961无蜡粉亮绿性状控制基因之间的关系,用LD10与10Q-961杂交获得另一个F1群体,调查发现该群体内23株单株均表现为叶面有蜡粉(图4、5)。表明这两份材料虽同为无蜡粉亮绿突变体,但控制无蜡粉亮绿性状的基因可能位于不同的遗传位点。
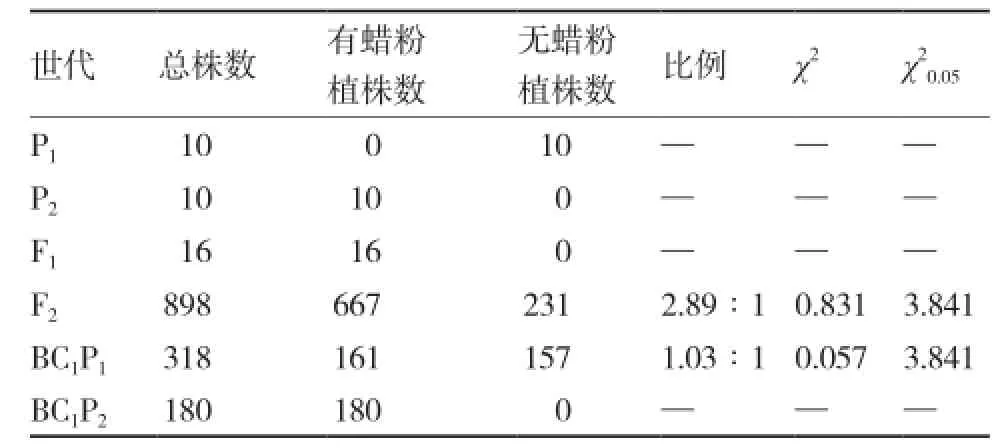
表1 LD10、21-3及其后代无蜡粉亮绿性状卡平方测验结果

图4 LD10、10Q-961及其F1叶片微观结构 (×3 000)

图5 LD10、10Q-961及其F1田间性状
2.4 引物筛选及连锁距离的确定
利用构建的无蜡粉单株DNA池和有蜡粉单株DNA池对4 753对引物进行筛选,其中Scaffolds SSR引物1 916对,根据甘蓝全基因组测序组装(王万兴,2013);SSR引物864对,根据甘蓝全基因组测序信息设计合成;EST-SSR引物1 013对,利用NCBI数据库中62 567条甘蓝类作物EST序列开发设计合成(陈琛 等,2011);Indel引物960对,根据两份骨干材料96-100和01-20重测序信息设计合成。在两池间表现出多态性的引物共168对,多态性比率为3.53%。结合叶面蜡粉有无的表型性状,利用亲本和F2群体进行复筛,结果表明1对引物(LT-SSR16)与甘蓝无蜡粉亮绿性状存在连锁关系,以该引物对F2群体898株单株进行验证(图6),利用Joinmap 4.0软件进行连锁分析,确定该标记与结球甘蓝LD10无蜡粉亮绿性状基因cgl-4的连锁遗传距离为4.7 cM。
3 结论与讨论

图6 引物LT-SSR16在F2群体部分单株中的扩增结果
本试验结果表明,结球甘蓝无蜡粉亮绿突变体材料LD10具有叶色亮绿、营养物质含量丰富等重要的商品性状和品质性状,扫描电镜观察发现LD10叶面只有极少量蜡粉覆盖,且蜡粉呈小颗粒状结构;而普通材料21-3叶面蜡粉含量丰富,蜡粉呈长棒状结构,两者差异明显,这与李景涛等(2012)、牟丽香等(2013)的研究结果一致。
芸薹属植物无蜡粉亮绿性状遗传规律较为复杂,不同材料之间差异较大,据报道多数突变体材料的无蜡粉亮绿性状为单基因隐性遗传,显性遗传或多基因遗传方式较少。本试验以结球甘蓝无蜡粉亮绿突变体材料LD10和普通有蜡粉材料21-3为亲本构建6世代群体,经卡平方检验各群体分离比例符合孟德尔隐性单基因遗传规律,由此判断LD10的无蜡粉亮绿性状受1对隐性基因控制,遗传方式与本所甘蓝课题研究组之前报道的无蜡粉亮绿突变体材料10Q-961相同(李景涛 等,2012),但这两份材料杂交后获得的F1群体单株全部表现为叶面有蜡粉,说明这两份材料的无蜡粉亮绿性状控制基因位于不同的遗传位点。本所甘蓝课题研究组共搜集得到6份无蜡粉亮绿甘蓝材料,经研究初步推断21-3油和大绿的无蜡粉性状由隐性基因控制,而10Q-385和10Q-974的无蜡粉性状由显性基因控制。作物育种的成效很大程度上取决于种质资源的数量,因此进一步搜集、研究更多具有无蜡粉亮绿性状的甘蓝突变体材料对选育商品性状美观、营养品质优良的甘蓝新品种具有十分重要的意义。
本试验筛选获得1个与LD10无蜡粉亮绿性状紧密连锁的分子标记,确定其与该性状控制基因cgl-4的遗传距离为4.7 cM,为进一步筛选与该性状连锁更加紧密的分子标记奠定了基础。在今后的工作中,可以通过合成并进一步筛选引物,获得与无蜡粉性状控制基因连锁更加紧密的分子标记,为结球甘蓝无蜡粉基因的精确定位和克隆奠定基础。
白凤虎,李德芳,陈安国,唐慧娟.2006.基于BSA分析法的分子标记基因定位技术在农作物中的应用.中国麻业科学,28(6):282-288.
曹阳,沈向群,李峰.2008.小白菜花茎无蜡粉性状遗传分析.中国蔬菜,(3):20-22.
陈琛,张兴桃,程斐,庄木,刘玉梅,杨丽梅,张扬勇,方智远.2011.秋甘蓝品种的SSR指纹图谱的构建.园艺学报,38(1):159-164.
初莲香,王余文.1993.无蜡粉亮叶结球甘蓝的发现及其利用.辽宁农业科学,(3):49-50.
方智远.2008.我国甘蓝产销变化与育种对策.中国蔬菜,(2):1-2.
冯辉,杨晓飞,辛彬,赵鹏涛,王玉刚.2010.无蜡粉型青梗菜核基因雄性不育系转育研究.沈阳农业大学学报,41(2):142-146.
李景涛,杨丽梅,方智远,刘玉梅,庄木,张扬勇,孙培田.2012.结球甘蓝10Q-961无蜡粉亮绿性状遗传规律初探.中国蔬菜,(12):37-41.
牟香丽,王超,王帅.2013.甘蓝无蜡粉突变体叶表皮蜡质超微结构观察.中国蔬菜,(4):32-37.
王万兴.2013.结球甘蓝高密度遗传连锁图谱的构建与主要农艺性状的QTL定位〔博士论文〕.北京:中国农业科学院.
谢景梅,刘晓兰,曲存民,卢坤,吕俊,李加纳.2002.甘蓝型油菜成熟籽粒DNA快速提取方法探讨.作物杂志,(1):17-21.
许绍斌,陶玉芬,杨昭庆,褚嘉.2002.简单快速的DNA银染和胶保存方法.遗传,24(3):335-336.
杨丽梅,方智远,刘玉梅,庄木,张扬勇,孙培田.2011.“十一五”我国甘蓝遗传育种研究进展.中国蔬菜,(2):1-10.
赵宝.2007.三种生态型盐芥(Thellungiella halophila)表皮蜡质及其合成相关的KCS家族各基因表达分析研究〔硕士论文〕.济南:山东师范大学.
赵建锋,沈向群,张海楼,刘镜,蒋守义,刘同.2006.菜薹无蜡粉性状遗传规律初探.园艺学报,33(3):538.
周熙荣,周志疆,李树林.1995.甘蓝型油菜无蜡粉性状的遗传性.上海农业学报,11(3):87-89.
Anstey T H,Moore J F.1954.Inheritance of glossy foliage and cream petal.Heredity,45:39-41.
Bernard A,Joubes J.2013.Arabidopsis cuticular waxes:advances in synthesis,export and regulation.Progress in Lipid Research,52:110-129.
Dietrich C R,Perera M A,Yandeau-Nelson M D,Meeley R B,Nikolau B J,Schnable P S.2005.Characterization of two GL8 paralogs reveals that the 3-ketoacyl reductase component of fatty acid elongase is essential for maize(Zea mays L.)development.The Plant Journal,42:844-861.
Eigenbrode S D,Shelton A M.1990.Behavior of neonate diamondback moth larvae(Lepidoptera:Plutellidae)on glossy-leaved resistant genotypes of Brassica oleracea L.Environmental Entomology,19(5):1566-1571.
Eigenbrode S D,Espelie K E,Shelton A M.1991.Behavior of neonate diamondback moth larvae〔Plutella xylostella(L.)〕on leaves and on extracted leaf waxes of resistant and susceptible cabbages.Journal of Chemical Ecology,17(8):1691-1704.
Lin J,Dickson M H,Eckenrode C J.1984.Resistance of Brassica lines to the diamondback moth(Lepidoptera:Yponomeutidae)in the field and inheritance resistance.Journal of Economic Entomology,77:1293-1296.
Mao B G,Cheng Z J,Lei C L,Xu F H,Gao S W,Ren Y L,Wang J L,Zhang X,Wang J,Wu F Q,Guo X P,Liu X L,Wu C Y,Wang H Y,Wan J M.2012.Wax crystal-sparse leaf2,a rice homologue of WAX2/GL1,is involved in synthesis of leaf cuticular wax.Planta,235:39-52.
Mo J G,Li W Q,Yu Q,Bodnaryk R P.1995.Inheritance of the waxless character of Brassica napus Nilla glossy.Canadian Journal of Plant Science,75:893-894.
Muria J,Taira T,Ohta D.1999.Isolation and characterization of the three Waxy genes encoding the granule-bound starch synthase in hexa-ploid wheat.Gene,234(1):71-79.
North C,Priestley W G.1962.A glossy-leaved mutant of Brussels sprout.Horticultural Research,1:95-99.
Pu Y Y,Gao J,Guo Y L,Liu T T,Zhu L X,Xu P,Yi B,Wen J,Tu J X.2013.A novel dominant glossy mutation causes suppression of wax biosynthesis pathway and deficiency of cuticular wax in Brassica napus.BMC Plant Biology,215(13):1-14.
Richardson A,Boscari A,Schreiber L,Kerstiens G,Jarvis M,Herzyk P,Fricke W.2007.Cloning and expression analysis of candidate genes involved in wax deposition along the growing barley(Hordeum vulgare)leaf.Planta,226(6):1459-1473.
Stoner K A.1992.Density of imported cabbageworms,cabbage aphids,and flea beetles on glossy and trichome-bearing lines of Brassica oleracea L.Journal of Economic Entomology,85:1023-1030.
Sturaro M,Hartings H,Schmelzer E,Velasco R,Salamini F,Motto M.2005.Cloning and characterization of GLOSSY1,a maize gene involved in cuticle membrane and wax production.Plant Physiology,138(1):478-489.
Zhang X,Liu Z Y,Wang P,Wang Q S,Yang S,Feng H.2013.Fine mapping of BrWax1,a gene controlling cuticular wax biosynthesis in Chinese cabbage(Brassica rapa L. ssp. pekinensis).Mol Breeding,32(4):867-874.
Zhou L Y,Ni E D,Yang J W,Zhou H,Liang H,Li J,Jiang D G,Wang Z H,Liu Z L,Zhuang C X.2013.Rice OsGL1-6 is involved in leaf cuticular wax accumulation and drought resistance.PLoS One,5(8):1-12.
Studies on Inheritance and Molecular Marker in Cabbage Glossy Wax-less Mutant LD10
LIU Dong-ming,YANG Li-mei*,TANG Jun,LIU Ze-zhou,FANG Zhi-yuan,LIU Yu-mei,ZHUANG Mu,ZHANG Yang-yong,SUN Pei-tian,LI Jing-tao
(Institute of Vegetables and Flowers,Chinese Academy of Agricultural Sciences,Beijing 100081,China)
To clarify the characteristics and inheritance of cabbage(Brassica oleracea L. var. capitata L.)glossy wax-less mutant LD10,the major agronomic characteristics and nutrient contents of LD10 were tested,and ultra-structures of leaves in mutant material LD10 and normal material 21-3 were measured,observed and comparatively analyzed. LD10 and 21-3 were taken as parent to construct 6 generations population,and inheritance analysis onthe glossy wax-less characteristics of LD10 was conducted. The results showed that there was little wax on the leaves of LD10,while the wax on the leaves of 21-3 was abundent,and the granular wax in LD10 was of long stick shape,significantly different from the wax in 21-3. The glossy wax-less trait in cabbage LD10 was controlled by one single recessive gene named cgl-4. One molecular marker(LT-SSR16)linked to cgl-4 with genetic distances 4.7 cM. Another F1population obtained through crossing LD10 and 10Q-961 showed that the genes leading to glossy wax-less trait were at different locations in the two glossy wax-less materials.
Cabbage;Glossy wax-less;Characteristics;Inheritance;Molecular marker
刘东明,男,硕士研究生,专业方向:蔬菜遗传育种,E-mail:liudongming5668@126.com
*通讯作者(Corresponding author):杨丽梅,女,研究员,博士生导师,专业方向:蔬菜遗传育种,E-mail:yanglimei@caas.cn
2014-04-08;接受日期:2014-05-23
国家大宗蔬菜产业技术体系项目(nycytx-35-gw01),国家“863”计划项目(2012AA100101),“十二五”国家科技支撑计划项目(2012BAD02B01),农业部园艺作物生物学与种质创制重点实验室项目
