白令海北部陆坡全新世以来的生物标志物记录及其古环境意义
2014-03-08张海峰王汝建陈荣华高爱国李丽王慧
张海峰 王汝建 陈荣华 高爱国 李丽 王慧
(1同济大学海洋地质国家重点实验室,上海200092;2国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州310012;3国家海洋局第二海洋研究所,浙江杭州310012;4厦门大学海洋与地球学院,福建厦门361005)
0 前言
白令海位于太平洋的最北端,是连接北冰洋和太平洋的唯一通道,白令海及北太平洋与北冰洋之间的水团交换过程对于理解物质和热量平衡及气候变化具有十分重要的意义[1]。前人研究表明,白令海是一个受风、海流和海冰综合控制的高生产力生态区系,其显著特征就是具有很高的表层生产力,尤其是在白令海盆东北和西北陆坡至陆架边缘的上部水体中存在一个高生产力带,被称为“白令海绿带”(Bering Sea Green Belt,BSGB)[2-4]。而其北部为宽阔的陆架,东、西两侧发源于阿拉斯加和西伯利亚的几条河流搬运来大量陆源物质,记录下丰富的海陆气候变化信息。前人研究发现晚更新世以来白令海的海洋环境与全球气候变化关系密切[5-9],其沉积物记录的古海洋学信息可以提供与米兰柯维奇轨道周期相关的北半球冰盖演化及高频率的Heinrich和D/O(Dansgaard/Oeschger)事件的记录,使其成为国际古海洋学和古气候学研究的热点之一[3-4,10-12]。
随着分析测试技术的发展,生物标志物(biomarker)分析方法已经广泛应用于海洋地质学以及古环境和古气候演变的研究中[13-17]。海洋沉积物中的生物标志物有两种组分,其中,甾醇、烯酮类组分是研究海洋初级生产力和表层生物结构的一种实用而有效的指标[14-16]。研究表明,沉积物中这些生物标志物的含量可以反映特定藻类的生产力,例如,菜子甾醇和岩藻甾醇可指示硅藻[18-19],甲藻甾醇可以反映甲藻[20],直链烯酮可以指示颗石藻[21-22],n-C30-diol则代表黄绿藻[23-24],这些藻类是目前大洋初级生产力的主要代表,其产生的类脂化合物具有不溶于水、不易挥发、化学性质稳定、抗生物降解能力强的特点,在沉降和成岩过程中依然保存有碳骨架,在许多地质环境中都可长期保存,因此可用于海洋浮游植物生产力和群落结构的重建,为研究地质历史时期的古生产力提供了重要的信息[25-29]。而烷烃类和醇类组分来自于陆地植被和海洋浮游藻类,但其中的长链正构烷烃和正构醇主要来自陆地高等植物的叶蜡表皮部分,可以通过河流和风搬运到海洋中。研究表明[30-31],高等植物叶蜡的长链正构烷烃主要分布在 n-C21—n-C35,具有明显的“奇高偶低”的奇碳数优势(即主要合成含奇数个碳原子的烷烃),通常是 n-C29,n-C31或 n-C33最高;而菌藻类的正构烷烃主要分布在低碳数n-C11—n-C25,以n-C17或n-C19占优势,没有明显的奇偶优势[31],前人也据此将n-C25—n-C33之间奇碳数的长链奇数正构烷烃含量的总和(表示为 ΣOdd(n-C25—n-C33),简称烷烃总量),作为研究陆源物质输入的替代性指标[30-32]。此外,还有一类沉水植物比较特殊,如狐尾藻和眼子菜等,其烷烃主要分布在n-C21—n-C25之间,主碳峰则是,明显区别于陆生植物[34]。
另外,长链正构烷烃的分子组组合特征,如碳优势指数(CPI)、平均链长指数(ACL)、烷烃指数(AI)以及n-C31/n-C27比值等,也常被用来进一步研究海洋沉积物中陆源物质的来源,恢复源区的植被特征和环境气候演变[16,32]。其中,CPI可指示烷烃的成熟度和来源,因为高等植物烷烃显著的奇偶优势在经历成岩、降解等作用后会降低,由此有别于化石烷烃[32,35-36];ACL,AI以及 n-C31/n-C27指标的建立则是基于现代植被的观测结果,即草本植物和木本植物都产生 n-C27,n-C29和 n-C31,但前者常以 n-C31为主峰,后者主要以n-C27或n-C29为主峰,因此这几个参数升降可代表草本植物相对贡献的增减,反映源区植被类型的变化[35-36]。
然而,生物标志物分析方法在白令海海洋地质学和古环境、古气候学研究工作中的实际应用却并不普遍[37-40],至于将该方法应用于该海区上层海洋环境和植物群落结构的变化以及陆源输送量的研究则更为缺乏。本文是在前人有限的工作基础上,利用AMS14C测年方法建立的高分辨率年龄框架,对在白令海北部陆坡采集到的沉积物柱状样进行了生物标志物分析,恢复该区全新世以来初级生产力和浮游植物群落结构的变化,探讨该区陆源输入量的变化及其源区植被和气候演化,以期进一步丰富白令海全新世以来高分辨率的古海洋学、古气候学研究。
1 现代海洋环境
现代白令海的海流系统主要为气旋式环流,海盆中央存在一个较大规模的逆时针环流系统(白令海涡流,BSG)[41]。北太平洋的水体从阿留申岛弧上各水道涌入白令海后汇入主表层流,其中最重要的即是白令陆坡流(BSC),该海流常年存在于白令海盆与东北部陆架之间陆坡处,沿着大陆架边缘自东南向西北流动,发育成为主表层环流的东部边界[1,42],其在空间范围上与 BSGB几乎重合[2-4]。对该区初级生产力、叶绿素含量以及物理海洋学和海水化学的现代观测表明[3,43],BSC的存在刺激了原地初级生产力的增强,导致在这样一个相对较窄但界限明显的区域范围内,浮游生物勃发、生物量增高。同时,由于太阳辐射量的变化以及海冰的进退,海水温盐也发生变化,导致BSGB的初级、次级生产力都具有季节性变化的特点。虽然BSGB生产力较高,但研究表明,大量的有机物并没有从白令海输出到开放大洋,而是就地堆积在陆架上,被底栖生态系统所利用[43]。
季节性海冰不仅对白令海上层海洋环境产生重要影响,还控制着具有极高生产力的春季浮游植物勃发的时间和地点[44],这种生物勃发足以提供全年的有机物沉降量,是整个大陆架生态系统的物质基础[43,45-46],尤其是当海冰向南扩张到大陆架坡折处,在海冰边缘区域存在一个与海冰和BSC更密切相关的春季生物勃发[47]。现代白令海由于受到较暖的太平洋水输入的影响,其阿留申海盆并没有海冰形成[48],海冰边缘通常在4月份某些时候就到达其最南的位置,平均覆盖范围从东到布里斯托尔湾(Bristol Bay)北端、西至陆架边缘普里比洛夫群岛(Pribilof Islands)区域的陆架上(图 1)[49-50]。
陆上入海河流不仅向白令海输送丰富的淡水,还搬运来大量的陆源物质。有研究指出,阿拉斯加和西伯利亚地区的诸多河流对白令海淡水输送的年平均速率至少在 104 m3·s-1[51],其中最重要的是育空河(Yukon River)[51](图 1),该河流水系发达、支流众多、水量充沛,是注入白令海长度最长、流域最广、输入贡献量最大的河流,也是其生态环境的基础[49,52-53],为白令海东部提供了最大的淡水输入、沉积物和溶解物质[54],其次是卡斯科奎姆河(Kuskokwin River)和阿纳德尔河(Anadyr River)[51,55]。有研究指出,白令海陆源物质碎屑的主要源区是阿拉斯加地区,由育空河和卡斯科奎姆河搬运至沉积区,其次是西伯利亚东部地区,主要通过阿纳德尔河输送[49](图1)。另外,还有一些小型河流也注入白令海,诸如堪察加河(Kamchatka River)[51]以及 Kvichak河、Nushagak河等[56]。

2 材料与方法
2.1 材料来源
本文研究的材料来源于1999年中国首次北极科学考察在白令海用重力活塞取样器钻取的B2-9站位沉积物柱状样样品[57]。该站位位于59°17′32″N、178°41′50″W,水深为2 200 m,处在白令海北部陆坡区(图1),柱状样长度为231 cm。柱状样的岩性单一,为一套深灰色的硅质生物软泥。采样间隔为2 cm,共获得115个样品。
2.2 研究方法
采集后的样品经提取和分离后进行了有机质定性和定量分析[32]。所提取和分析的有机质主要包括:长链正构烷烃类、长链正构醇类、烯酮类和甾醇类。有机质提取和分离方法为:将样品冷冻干燥,充分研磨后称取2—4 g装入10 mL的TFL管中,加入甲醇∶二氯甲烷(V∶V=1∶3)混合溶液萃取有机质;重复萃取4—5次;将上层萃取液收集到玻璃瓶中,在氮吹仪下吹干后加入含6%KOH的甲醇溶液进行碱水解;室温下放置约12 h后用正己烷萃取4—5次,收集的上层萃取清液经氮吹浓缩后进行硅胶柱层析分离;用正己烷淋洗收集烷烃组分,再用二氯甲烷(内加5%甲醇)淋洗收集醇类、烯酮组分,分别经吹干、浓缩后转移至细胞瓶中;烷烃组分定量加入30μL异辛烷后直接用气相色谱分析,醇类、烯酮组分则需进行衍生化反应(加入BSTFA)后再进行气相色谱分析。
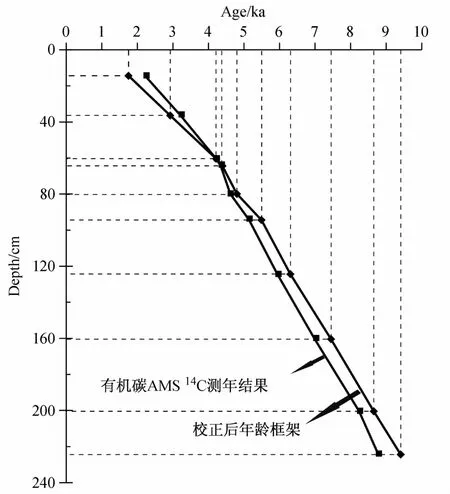
图2 白令海北部陆坡B2-9站位年龄框架Fig.2.The age model of Site B2-9 from northern Bering Sea Slope
气相色谱分析采用Finnigan Trace GC Ultra气相色谱仪测试,色谱条件:HP-1J色谱柱,柱长60 m,内径 0.32μm,涂层 0.17μm。FID检测器温度280℃,进样口温度:280℃。分5段升温程序,采用无分馏进样,载气为 He,流速 1.0 mL·min-1。生物标志物的定性主要采用内标和目标峰的位置(保留时间)来定性,定量采用n-C24D50,n-C19H39OH内标法,通过目标峰值与内标峰值来计算。利用内标物质进行检验,该方法回收率>85%,4次平行实验结果的标准偏差<1%。以上分析和测试工作均在同济大学海洋地质国家重点实验室完成。
2.3 年龄框架
本文的年龄框架是根据10个样品的有机碳AMS14C测年数据建立的,校正程序为Calib 6.0程序,校正曲线选用mixed Marine&NH Atmosphere混合曲线,经多次校正对比,认为海源有机碳平均含量选取75%最合理。校正后得到顶部年龄约980 a,底部年龄9 590 a(图2),平均取样分辨率达到75年/样(据王汝建等,未发表资料)。
3 研究结果
3.1 浮游植物群落的变化
本文利用B2-9站位沉积物中甾醇、烯酮类生物标志物的含量反映特定藻类的生产力(菜子甾醇-硅藻、甲藻甾醇-甲藻、长链烯酮-颗石藻、n C30-diol-黄绿藻),而以硅藻、甲藻、颗石藻和黄绿藻为代表的浮游藻类是目前大洋初级生产力的主要贡献者;同时本文参考前人的研究结果[14-24],以这四种藻类反映浮游植物群落结构的变化,并将其含量总和记为浮游植物总量,作为初级生产力的替代性指标。
数据显示,近1万年以来,B2-9站位沉积物中4种浮游藻类一致呈现出“S”型的变化趋势(图3,ad),虽然颗石藻的这种变化趋势稍弱,但整体上仍呈现出升高-降低-再升高的趋势(图3,d),浮游植物总量也相应呈现出十分明显的“S”型变化趋势(图3,e)。同时,这种变化趋势也具有明显的阶段性特点(图 3,a-e):9.6—6.9 ka BP,4种藻类的含量和浮游植物总量都处于高值期;6.9—3.5 ka BP,浮游植物生产量都有所降低,波动幅度较小;4.4—3.5 ka BP期间达到全新世以来的最低水平;3.5—1.0 ka BP,浮游植物的生产量逐渐增加,至1.0 ka BP几乎达到全新世以来的最高值。值得特别注意的是,在早全新世的生产力高值期,4种藻类的生产力以及浮游植物总量都经历了两次显著的降低过程,分别出现在9.4—9.1 ka BP和 8.0—7.7 ka BP期间。

图3 白令海北部陆坡B2-9站位全新世以来浮游植物群落结构及其变化.其中,a—d分别表示硅藻、甲藻、黄绿藻、颗石藻含量的变化;e为浮游植物总量的变化,反映初级生产力的变化;f为硅藻和甲藻占浮游植物总量的比例,反映两者对初级生产力的贡献;水平虚线标示变化的时间界限,竖向虚线标示50%贡献率,实线显示数据的S型变化趋势,灰色阴影标示早全新世快速气候变化事件Fig.3.Phytoplankton community structure and its changes from Site B2-9 of northern Bering Sea Slope during the Holocene.a—d,the changes of diatoms,dinoflagellates,eustigmatophyceae,coccolithophores,respectively.e,the change of total phytoplankton,indicate the evolution of primary productivity.f,percentage of diatoms and dinoflagellates in total phytoplankton,show their contribution to primary productivity.Horizontal dashed lines indicate the time boundary of changes.Vertical dashed line is level of50%.Solid lines show the trend of changes,gray rectangles show the rapid climate change during early Holocene
从浮游藻类的组合结构来看,几乎每个样品中硅藻的含量都是最高的,4.4—3.5 ka BP期间降幅最明显(图3,a);其次是甲藻,其生产量与硅藻在同一个数量级,个别样品中甚至高于硅藻(图3,b);黄绿藻和颗石藻的生产量都比硅藻和甲藻低1个数量级(图3,c、d)。为进一步研究浮游植物之间的共生组合关系,分别计算4种藻类生产量占浮游植物总量的百分比,发现硅藻和甲藻具有十分明显的“此消彼长”的竞争关系(图3,f)。同时,硅藻对浮游植物总量的贡献率存在阶段性的变化,以5.6 ka BP为界,此前低于50%,此后几乎都高于50%,相对而言,甲藻的贡献率整体降低(图3,f)。
3.2 长链正构烷烃的变化
白令海B2-9站位沉积物样品中检测出的长链正构烷烃的碳数主要分布在n-C21至n-C35之间,奇数碳优势明显,但与其他海区相比[16,32,58],其主要碳数分布特征略有不同。首先,几乎所有样品的主碳峰都是n-C27而非 n-C29或n-C31,相比之下,虽然n-C25、n-C29和n-C31也都是较高的碳峰,但都低于n-C27,这与卢冰等[38-39]的研究结果是一致的;其次,在其他海区沉积物中含量较高的 n-C33在本区较低[16,32],与此相反的是 n-C23的含量较高,几乎在每个样品中都高于n-C33的含量。本文采用ΣOdd(n-C25—n-C33)计算烷烃总量,作为陆源物质输入的替代性指标,并单独对 n-C23加以讨论。另外,通过CPI,ACL,n-C31/n-C27以及 AI这几个参数进一步分析海洋沉积物中烷烃的来源,重建其源区的植被特征和气候与环境的演变。相关参数的定义公式及在本文中的指示意义见表1。

表1 烷烃各参数公式及其环境指示意义Table 1.Alkane formula index and their environmental implications in this paper
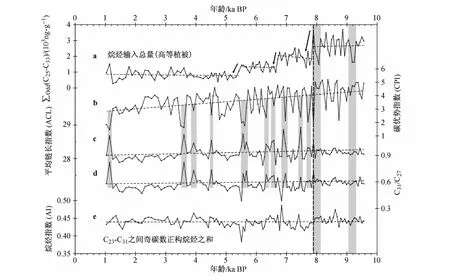
图4 白令海北部陆坡B2-9站位全新世以来烷烃总量及其分子组合特征的变化.其中,a—e分别表示烷烃输入总量、碳优势指数、平均链长指数、C31/C27比值和烷烃指数的变化;实线箭头和水平虚线显示数据变化趋势,竖向虚线和灰色长阴影区标示各参数对早全新世两次快速气候变化事件的响应,灰色短阴影区标示CPI、ACL及C31/C27变化的对应关系Fig.4.Total n-alkane and molecular parameters changes from Site B2-9 of northern Bering Sea Slope during the Holocene.a—e,the changes of total n-alkane,carbon preference index(CPI),average chain length(ACL),ratio of C31/C27 and alkane index(AI),respectively.Solid arrows and horizontal lines show the trends of data,vertical dashed lines and long gray rectangles indicate their response to the rapid climate change during early Holocene,short gray rectangles show the synchronization of CPI,ACL and C31/C27 changes
白令海B2-9站位近1万年以来烷烃总量的变化范围在 275.3—3 694.8 ng·g-1之间,平均值均1 681.2 ng·g-1,其中最高值出现在约 8.9 ka BP,最低值出现在约1.2 ka BP,这种显著的差别与烷烃总量整体降低的趋势是一致的(图4,a)。同时,烷烃总量的变化也具有明显的阶段性:第一阶段,9.6—7.8 ka BP,烷烃总量波动较快,变化幅度也较大,但总体上较为稳定,平均值较高,达2 657.3 ng·g-1;第二阶段,7.8—6.7 ka BP,烷烃总量变化的频率和波动幅度都有所降低,呈略有增加的趋势,但整体水平都低于第一阶段,平均值为1 948.7 ng·g-1;第三阶段,6.7—5.4 ka BP,烷烃总量进一步降低,平均值为1 304.6 ng·g-1,变化趋势与第二阶段相似,但更为平稳;第四阶段,5.4—1.0 ka BP,烷烃总量总体上变化较为平稳,平均值为4个阶段中的最低值,但也达到814.2 ng·g-1,仍然属于全新世陆源烷烃输入水平较高的海区,明显高于其他海区,如南大洋南塔斯曼海[16]、中国南海北部[32]。
与烷烃总量呈阶梯状降低的变化模式相较,CPI等指标的变化略有不同。CPI值整体呈下降趋势,但无明显的阶段性变化,其值在1.47—5.19之间,平均值为3.63,且绝大部分都大于3,只有极少几个值小于 2(图 4,b)。ACL、AI和 n-C31/n-C27的变化相对都比较稳定,其中,ACL的变化范围在27.81—28.93之间,平均值 28.18,除个别时期外,ACL的变化都相对稳定(图 4,c);n-C31/n-C27变化范围介于0.39—0.82之间,平均值为 0.58,除个别时期外,其变化与 ACL相似(图4,d);AI的变化范围在0.38—0.49之间,平均值为0.44,除个别时期外,其变化趋势最为稳定,波动幅度最小(图4,e)。
如前所述,B2-9站位沉积物中检出的长链正构烷烃有两个显著的特点:第一,n-C27的含量最高,其变化范围介于90.5—1 187.3 ng·g-1之间,平均含量达494.2 ng·g-1,其变化趋势也与烷烃总量十分一致(图5,a、b),相关性 R2=0.98,对烷烃总量的贡献率介于26%—44%之间,贡献最大且总体上较为稳定(图5,c);第二,几乎每个样品中的 n-C23含量都比 n-C33含量高,前者含量介于57.3—627.3 ng·g-1之间,平均值为 234.8 ng·g-1(图 5,d),明显高于后者95.6 ng·g-1的平均值。n-C23含量的阶段性变化的特点也很明显,在8.2—7.8 ka BP期间经历了降低后略升高,但并未再达到之前的水平,而在7.3—7.1 ka BP间,n-C23含量达到全新世的最高值。
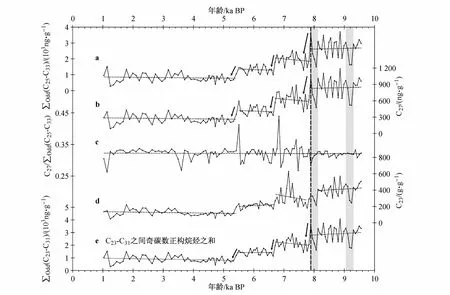
图5 白令海北部陆坡B2-9站位全新世以来烷烃总量和单体碳同位素的变化.其中,a为烷烃输入总量的变化;b、d分别为单体碳同位素n-C27、n-C23的变化,c为n-C27对烷烃输入总量的贡献率,e为n-C23至n-C31之间奇碳数正构烷烃之和,以与烷烃输入总量进行对比;实线箭头和水平虚线显示数据变化趋势,竖向虚线和灰色阴影区标示早全新世期间烷烃总量和单体碳同位素两次显著的快速变化事件Fig.5.Total n-alkane andmono-carbon isotope changes from Site B2-9 of northern Bering Sea Slope during the Holocene.a,the changes of total n-alkane.b and d,show the changes of n-C27 and n-C23.c,show the contribution of n-C27 to total n-alkane.e,sum of odd n-alkane from n-C23 to n-C31,to compare with total n-alkane.Solid arrows and horizontal lines show the trends of data,vertical dashed lines and gray rectangles indicate the rapid changes of total n-alkane and mono-carbon isotope during early Holocene
4 讨论
4.1 初级生产力与浮游植物群落结构的变化
前文已详述了B2-9站位1万年以来浮游植物的变化特点,这种几乎一致的“S”型的变化趋势表明,全新世以来白令海北部陆坡区浮游植物群落的结构较为稳定,主要藻类成员对于上层海洋环境的变化具有相似的响应过程。在B2-9站位所处的BSGB高初级生产力的背景下,对初级生产力和浮游植物群落结构的变化模式进行深入分析具有重要意义。
季节性海冰作为白令海的一个重要的特征,对其上层海洋环境的影响是显而易见的。有研究指出,11 ka BP海冰已经退却到白令海东南部Umnak Plateau的北部[59]。进入早全新世,随着海平面的快速上升,白令海峡重新打开[60]。到8 ka BP,白令陆桥的海岸线达到或接近现代的界限[61],白令海已经达到现代的海洋学条件[62],与北太平洋之间的所有通道都被打开,海冰已经有好几千年没有在白令海盆中形成[59]。Katsuki等[63]的研究指出,全新世白令海海冰的分布主要受控于阿留申低压的位置,海冰的进退据此分为3个阶段:早全新世(7 ka BP以前),受位于白令海西部的阿留申低压及位于阿拉斯加湾的小型低压系统的控制,海冰扩张;中全新世(7—3 ka BP期间),阿拉斯加湾上空的低压系统减弱,引起整个白令陆架上的海冰消退;晚全新世(3 ka BP以来),由于阿留申低压发育引起的西南盛行风导致白令陆架上的海冰覆盖面积进一步减小。这些研究结果说明早全新世(7 ka BP之前)白令海海冰在陆架上扩张,但并未到达白令海盆区域,海冰的南部边缘就在白令陆架坡折至陆坡一线,7 ka BP以来就一直分布于白令海东部和北部陆架浅水区域[59,64]。根据 Caissie等人[59]的研究结果,11.3 ka BP以来,白令海东南部硅藻组合从以海冰种为主转变为以北太平洋的暖水种为主;与此同时,表层海水温度(SST)较此前上升了3℃,白令海南部终年无冰,进入全新世后,SST也经历了升高-降低-升高的变化模式。Max等人[64]的相关研究也指出,进入早全新世,白令海西部经历了SST上升的过程,自9 ka BP以来则逐渐降低,但在约3.5 ka BP以来,SST开始升高。
据此,可以解释全新世以来B2-9站位沉积物中浮游植物含量和初级生产力的变化:早全新世(9.6—6.9 ka BP),海冰最远扩张到陆架-陆坡边缘一线,与此同时,白令陆坡流的上升带来了下层海水中的营养物质,引起了硅藻、甲藻等的春季勃发。这为初级生产力的升高做出了巨大贡献,而喜海冰的属种有可能在其中起到了主要作用。到了夏季,较高的SST和太阳辐射量(图6,b)促进了颗石藻等的勃发,进一步提高了初级生产力;中全新世(6.9—3.5 ka BP),由于阿留申低压系统向白令海西南方向迁移、阿拉斯加湾上空的低压系统减弱甚至消失[63],导致海冰消退至白令陆架上,白令陆坡区早春季节与海冰相关的生物勃发减弱甚至消失,导致浮游植物的生产量和初级生产力都逐渐降低。同时,由于区域温度的降低[59,63],尤其是约 4.7 ka BP左右开始的所谓新冰期冷事件[65],导致海洋表层海水变冷,上层海洋环境恶化,甚至有可能影响富营养盐的太平洋暖水的输入,进一步阻碍了浮游藻类的繁殖;晚全新世(3.5—1.0 ka BP),阿留申低压系统继续发育,盛行西南风导致海冰被限制在陆架之上,陆坡区未再受到海冰的影响,营养盐的供给成为控制初级生产力的主要因素,白令陆坡流源源不断地把富营养盐的海水输送到北部陆坡区,为浮游植物带来大量的营养物质;SST和太阳辐射量都呈现出上升的趋势,表层海洋环境趋于温暖,促进了浮游植物的生产;同时在沿白令海陆坡转折带,潮汐混合作用和横向环流导致海水分层较差,300—800 m水深的营养盐得以到达表层水体,使表层生产力增加、甚至可以达到热带的高生产力水平[2]。
值得注意的是,浮游藻类含量和浮游植物总量所指示的初级生产力在早全新世都经历了两次幅度较大的降低过程,分别出现在9.4—9.1 ka BP和8.0—7.7 ka BP期间(图3,a-e,图 6,a)。与格陵兰NGRIP冰芯中δ18O记录的对比后发现:这两次初级生产力降低的过程之前,冰芯中的δ18O也记录两次快速降温事件,分别出现在9.4—9.2 ka BP和8.4—8.1 ka BP期间(图6,c),但前者比后者分别滞后约100 a和约300 a(图6,a、c)。这表明白令海是一个良好的、全球气候变化的响应区域,甚至是一个放大器。这种滞后现象可能受控于气候变化信号的传播,同时由于海水的热容比较高,对气候变冷的响应幅度变小、响应过程略有滞后,而浮游藻类突然降低是其对表层海洋环境变化的敏感响应。

图6 全新世以来白令海北部陆坡B2-9站位浮游植物总量与60°N-8月太阳辐射量以及NGRIP冰芯δ18 O的对比.其中,a为浮游植物总量的变化;b为60°N-8月太阳辐射量的变化,数据来自文献[66];c为格陵兰NGRIP冰芯中δ18 O的变化,数据来自文献[67],并对数据进行了7点平滑处理;灰色阴影标示早全新世快速气候变化事件Fig.6.Contrast between total phytoplankton from Site B2-9 of northern Bering Sea Slope and insolation of 60°N-August,δ18 O of ice core from NGRIP during the Holocene.a,the change of total phytoplankton.b,insolation of60°N-August,the data obtained from[66].c,7-point smoothedδ18O of ice core from NGRIP during Holocene,the data obtained from[67].Gray rectangles show the rapid climate change during early Holocene
浮游藻类几乎一致的变化表明,全新世以来白令海北部陆坡区表层浮游植物群落结构较为稳定,这可能与营养盐的浓度或分布结构有关。其次,可以直观地看出硅藻是白令海区初级生产力的最主要贡献者,甲藻次之,而颗石藻和黄绿藻则低1个数量级。这可能反映硅藻、甲藻是白令海春季勃发的主要成员,而颗石藻、黄绿藻的勃发则是在夏末季节[59,64],表 层 水 体 中 的 营 养 盐 浓 度 低 甚 至 被 耗尽[4],导致浮游藻类生产量显著降低。此外,硅藻勃发吸收大气中的CO2,形成有机质,沉降到海底并被埋藏在沉积物中,而颗石藻在吸收CO2的同时其钙质的壳体又会被溶解而释放出CO2,因此,较高的硅藻生产力和较低的颗石藻生产力表明白令海是较强的CO2汇区。
另外,硅藻和甲藻之间明显的竞争关系表明两者的生活环境较为相似,对营养物质的摄取具有竞争性(图3,f),由于白令海水体中硅质的含量很高[10-11],硅藻在竞争中占优势,同时,可能由于某些海冰种对较冷的生活环境也更加适应,因此其生产量居于主要地位。这与在本区开展的研究工作和现代观测结果是一致的[68-69]。5.4 ka BP前后两者对浮游植物总量贡献率的变化是对上层海洋环境变化的响应:烷烃总量在5.4—5.0 ka BP下降至进入全新世以来的最低水平,表明陆源物质输入经历一次较大的降低过程,表层海水中陆源有机质的供应量降低,在一定程度上限制了甲藻的生产,而海水中硅质含量受影响较小,足以保障硅藻的生命活动,因此出现了硅藻贡献增加而甲藻贡献降低的现象。
不过,也有学者指出,由于沉积物中生物标志物的含量低,容易受到陆源物质输入“稀释”作用的影响,因此严格上讲不宜用来指示初级生产力的变化[25,70]。但通过与烷烃总量变化趋势的对比发现,早全新世的烷烃总量最高,同期浮游植物的生产量也处于高值,并没有降低;晚全新世两者相反的变化趋势也并不具有内在的相关性,因此,陆源物质输入的“稀释”作用对生物标志物相对含量的影响较小,生物标志物的相关参数仍然可以作为浮游植物群落结构和初级生产力的替代性指标。
4.2 高等植被输入与源区的气候变化
一般来说,海洋沉积物中保存下来的烷烃总量受控于两个因素:高等植物源区烷烃的生产量及其向海洋中输送的环境条件,这两者都主要取决于气候和环境的变化。陆源物质输送到海洋中主要有两种方式[71]:河流直接输入[72-73]以及经空气和风尘的搬运输入[74-75]。这也是陆地高等植物生产的长链正构烷烃输入海洋、沉降、并保存在沉积物中的主要方式[32]。贺娟等人[32]在南海北部的研究中指出:最有利于烷烃输送和保存的环境条件是温暖潮湿环境和低海平面时期。白令海地区虽处亚北极高纬地区,但其东西两侧紧邻广袤的陆源植被区,其接受烷烃输入、保存的条件与南海北部相似。温暖潮湿的环境有利于源区植被生长,可以生产大量的烷烃;而低海平面不仅增加了陆源植被生产源区的分布面积,也缩短了河口到陆坡沉积区的距离,有利于烷烃输送到此。另外,在偏北的东/西风盛行的年份,通过大气的传播,也有利于把阿拉斯加和东西伯利亚广大地区的烷烃输送到白令海。
由于B2-9站位所处的白令海北部陆坡区位置较为特殊,既会受到白令海陆坡流的影响,也受到育空河、卡斯科奎姆河以及阿纳德尔河淡水输入的影响,甚至还受到西伯利亚高压和阿留申低压系统耦合消长的影响[59,65]。末次冰期以来,海冰进退的锋面也基本上沿陆坡边缘一线分布[64],进入全新世以后,海冰则沿此线逐渐消退至陆架内部[63]。尽管在早全新世冬季海冰前缘分布在陆坡边缘一线,可能封锁了育空河等河流向白令海输送的表层通道,但是冬季可能并非烷烃生产和输出的主要时期;到了夏季海冰全面消退,烷烃得以通过河流和大气输送,因此,海冰对陆源烷烃的输入可能有一定的限制作用。中晚全新世,海冰消退至白令陆架内部,最远推进距离也很少能够到达陆坡边缘[63],海冰对于陆源烷烃的输入几乎可以忽略。
全新世海平面变化是陆源物质输入的一个非常重要的影响因素。11 ka BP,全球海平面比现在低50—60 m,之后逐渐上升,到6 ka BP达到现代海平面的高度,之后基本保持不变[76-79]。海平面持续上升的结果是导致海岸线向内陆方向收缩,白令海东西两侧沿岸地区出露的大片陆架被海水淹没,导致陆架上的陆生植物死亡,植物残体在海平面上升过程中会被洋流输送到陆坡区,甚至海盆当中。同时,海平面上升也导致育空河等河流入海口向内陆退移,增加了陆源物质从河口到陆坡区的搬运距离,进而致使随河流输送到陆坡区的陆源物质减少,导致9.6—5.4 ka BP烷烃输入量阶段性的下降(图4,a)。自5.4 ka BP以来,海平面趋于稳定,陆源烷烃输入总量减少。Hopkins指出[80],6—5 ka BP期间白令海地区又发生了一次海侵,B2-9站位沉积物记录了这次海侵过程,烷烃总量在约200 a的时间里下降了近1/2,这也是全新世烷烃总量的最后一次阶段性的降低,此后烷烃总量保持一个较为稳定的水平(图4,a)。
持续性的海平面上升对烷烃输入的影响是渐变的,因此对于约6.7 ka BP和7.8 ka BP前后烷烃总量的阶段性降低过程来说,可能主要受控于源区气候与环境的变化。Katsuki等人[63]指出,早全新世由于阿拉斯加湾上空低压系统的影响,阿拉斯加南部地区总体上是温暖、湿润的气候。但7.0 ka BP以来,阿拉斯加湾上空的低压系统消散,导致阿拉斯加南部地区的气候由暖湿变为湿冷,这样的气候转变可能导致了植被分布范围的收缩,同时由于大气压力系统的改变,风向也发生变化,导致陆源烷烃的输送主要依靠河流,而风力搬运减弱,致使研究区6.7 ka BP以来烷烃总量的阶段性下降(图4,a)。与此相较,烷烃总量在7.8 ka BP的阶段性降低的特点及其控制因素又稍有不同:由图4a可以看出,约8.0 ka BP前后的200年里,烷烃总量处于第一阶段的低值期,甚至低于第二阶段烷烃总量的平均水平,但在约7.9 ka BP年重新恢复到最高值,此后进入第一、二阶段的阶梯式下降过程。这种变化可能是对8.2 ka BP冷事件的响应,这次气候变冷事件虽然持续时间只有约400 a,但其降温幅度较大,甚至可达YD事件的一半[81-82]。在早全新世较为温暖湿润的气候条件背景下,这种突然的剧烈降温事件对陆生植被的破坏是十分严重的。Hu等人[83]的研究也表明,在11—8.0 ka BP的最后时期,阿拉斯加西南部地区植被群落的丰度都降低了,而这对白令海陆源烷烃输入量的影响是不可逆转的,直接导致烷烃总量从第一阶段下降到第二阶段。结合C27含量以及 CPI和 ACL的变化(图 4,b、c,图 5,b)可以发现,化石烷烃和n-C27的含量在约7.9 ka BP时期都增加了,但是n-C27对烷烃总量的贡献量却下降了(图5,c)。因此,在7.9 ka BP烷烃总量的回升有可能是化石烷烃贡献量的增加造成的,而不是陆源植被烷烃的增加。而9.3—9.1 ka BP烷烃总量的降低明显响应于格陵兰冰芯中记录的9.3 ka BP冷事件。
碳优势指数(CPI)平均值为3.63,且在大部分样品中都>3,极少数<2,表明沉积物中长链正构烷烃基本上都来自于陆源植被,而不是高成熟度的化石烷烃。CPI整体持续下降的趋势表明,陆源植物烷烃对烷烃总量的贡献逐渐降低。这可能是由于全新世以来海平面的持续上升,逐渐淹没了陆源植被区,陆源植物烷烃对烷烃总量的贡献逐渐减少,而CPI的持续下降可能意味着化石烷烃输入量的逐渐增加。结合n-C27对烷烃总量的贡献(图5,c),ACL、n-C31/n-C27以及AI的变化特点表明,全新世以来源区植被结构较为稳定,并以木本植物占据优势。中晚全新世几个ACL峰值与CPI的低值具有较好的对应性(图4,b-d),可能并不是陆源植被中草本植物的比例增加,而是木本植物的生产力相对降低,烷烃供应量相对减少,化石烷烃的贡献相对增加。不过,CPI等参数在早全新世的两次快速降温期间却并未出现相应的大幅度变化,这可能与海陆气候之间信号传递和沉积物早期成岩作用有关。
全新世以来,n-C27含量对烷烃总量的平均贡献率达到32%,其变化趋势几乎与后者完全一致(图5,a-c),甚至可以直接替代后者作为烷烃输入总量的指标。这可能是因为源区植被中木本植物对气候变化的响应。有学者认为白令海地区的气候最适宜期大致开始于8 ka BP或10 ka BP以前,杨属植物的界线比现在广,早全新世海侵以来,阿拉斯加东岸的森林在6—5 ka BP扩展到现在的位置[84-85]。大约6 ka BP以来,黑云杉到达阿拉斯加山脉以北的广大地区,并随之成为森林生态系统中的优势树种,这种变化是森林生态系统对晚全新世较冷和较湿的气候环境的综合响应[83]。而 n-C27含量对9.3 ka BP和8.2 ka BP的快速降温事件的响应都十分明显,表明木本植物的生长受到了寒冷气候的限制。
在B2-9站位沉积物中,高含量的n-C23与n-C27及其他们与烷烃总量的相关性都较高,若计算n-C23—n-C31之间奇碳数正构烷烃的总和∑Odd(n-C23—n-C31),可以看出其变化趋势与∑Odd(n-C25—n-C33)十分相似(图5,a、e)。这些都表明 n-C23与 n-C27等其他长链正构烷烃受到某种相同因素的影响。然而,n-C23主要产自沉水植物[33],在本研究区其主要可能来自于北半球沿海分布最广泛的一类海草(其典型代表是大叶藻)[86],是一种较为特殊的高等植物,可以在海洋中进行沉水生活,并能在海水中完成开花、结种和萌发这一生命史[87]。因此,为了保持正构烷烃总量计算与CPI等参数计算上的一致性,减小其他不确定因素的影响,本文仍采用∑Odd(n-C25—n-C33)作为烷烃总量的计算公式,而将n-C23排除在外。不过,n-C23的变化趋势在一定程度上也可以反映源区的气候变化。Hu等人[83]对阿拉斯加西南部地区Farewell湖沉积物中孢粉的研究发现,该区的植被结构与阿拉斯加中、北部地区的植被分布颇为相似[88-89],都显示 11—8.0 ka BP夏季温度比现代高[89-91],温暖干燥的夏季气候以及北半球高纬地区太阳辐射量达到最大值引起了湖水变深、水生生产力增加[83],这促进了n-C23生产量的增加。同时,其含量在约9.3 ka BP和8.1 ka BP快速下降(图5,d),是对早全新世两次冷事件的响应(图6,c),可以推测气温降低直接导致海洋沉水植物生长衰退;但此后与烷烃总量和n-C27不同的是,n-C23含量在7.2—6.7 ka BP达到了早全新世的水平。这可能主要是由湖泊、海洋生产力增加造成的,也反映了n-C23与陆生植被生产的烷烃在指示意义上的差异。
5 结论
本文通过白令海北部陆坡B2-9站沉积物样品的AMS14C测年和生物标志物分析,获取了近1万年来高分辨率的浮游植物群落和初级生产力记录,以及陆源烷烃的输入及其源区植被与气候和环境等特征,得出如下认识:
(1)白令海北部陆坡区全新世以来的表层浮游植物和初级生产力都经历了“高-低-高”的变化过程,具有明显的阶段性,9.6—6.9 ka BP初级生产力的升高,6.9—3.5 ka BP初级生产力下降,3.5—1.0 ka BP初级生产力再次升高。这种变化模式受控于陆架坡折处海冰的分布、上层海洋营养盐供应和早全新世9.3 ka BP和8.2 ka BP快速降温事件的影响。
(2)白令海北部陆坡区全新世以来的浮游植物群落的结构基本上是稳定的,硅藻是初级生产力的主要贡献者,甲藻次之,而颗石藻和黄绿藻比前两者低了1个数量级。这是由于表层海洋环境不同导致浮游藻类生产力不同;同时硅藻和甲藻存在明显的竞争关系,但由于海水中硅质供给充足,硅藻在竞争中明显占据优势,成为白令海有机碳汇的主要贡献者,与现场实测结果吻合。
(3)白令海北部陆坡区全新世以来的烷烃总量变化经历了3次阶梯状的下降过程,分别出现在7.8,6.7和5.4 ka BP,烷烃总量变化呈现出 4个相对稳定的阶段,烷烃的输入主要受控于早全新世海平面上升以及周边陆地植被源区的气候与环境变化;CPI指数表明烷烃主要来自于陆源高等植物,并呈现出下降的趋势,而ACL等相关参数表明烷烃源区的植被结构较为稳定,以木本植物占据优势。
(4)白令海北部陆坡区全新世以来的单体碳同位素研究发现,正构烷烃具有两个显著的特征,主碳峰是n-C27,并且n-C23含量较高。n-C27含量对烷烃总量的贡献最大,平均可达1/3,甚至可以代替烷烃总量作为陆源烷烃输入的指标。这可能与陆地繁盛的木本植物及其分布有关;含量较高的n-C23则可能主要来源于北半球沿海广泛分布的海生沉水植物,其次可能是陆地湖泊水生植物。
致谢 该项工作是由国家财政部资助,国家海洋局极地办公室组织实施的“中国首次北极科学考察项目(CHINARE-1999)”的一部分,参加此项工作的单位有中国极地研究中心、国家海洋局第一研究所、国家海洋局第二研究所、国家海洋局第三研究所等。感谢中国首次北极科学考察队的全体科考队员和“雪龙”号全体船员为沉积物样品的采集所付出的艰辛努力。样品由中国极地研究中心沉积物库提供。在生物标志物实验分析测试工作中得到了同济大学李文宝博士的支持和帮助,在文章撰写过程中同济大学贺娟老师、党皓文、董良等同学以及国家海洋局第二海洋研究所卢冰老师提出了宝贵意见,在此一并表示感谢。
1 Stabeno P J,Schumacher JD,Ohtani K.The physical oceanography of the Bering Sea//Loughlin T R,Ohtani K.Dynamics of the Bering Sea.Fairbanks:University of Alaska Sea Grant,1999:1—28.
2 Springer A M,McRoy C P,FlintM V.The Bering Sea Green Belt:shelf-edge processes and ecosystem production.Fisheries Oceanography,1996,5(3-4):205—223.
3 Wang R J,XiaoW S,LiQ Y,et al.Polycystine radiolarians in surface sediments from the Bering Sea Green Beltarea and their ecological implication for paleoenvironmental reconstructions.Marine Micropaleontology,2006,59(3-4):135—152.
4 陈立奇,高众勇,王伟强,等.白令海盆p CO2分布特征及其对北极碳汇的影响.中国科学(D辑),2003,33(8):781—790.
5 Gorbarenko SA.Stable isotope and lithologic evidence of late-glacial and Holocene oceanography of the northwestern Pacific and itsmarginal seas.Quaternary Research,1996,46(3):230—250.
6 Gorbarenko SA,Basovb IA,Chekhovskayac M P,etal.Orbital andmillennium scale environmental changes in the southern Bering Sea during the last glacial-Holocene:Geochemical and paleontological evidence.Deep-Sea ResearchⅡ,2005,52(16-18):2174—2185.
7 Cook M S,Keigwin L D,Sancetta C A.The deglacial history of surface and intermediatewater of the Bering Sea.Deep-Sea ResearchⅡ,2005,52(16-18):2163—2173.
8 Okazaki Y,Takahashi K,AsahiH,etal.Productivity changes in the Bering Sea during the late Quaternary.Deep-Sea ResearchⅡ,2005,52(16-18):2150—2162.
9 Brunelle B G,Sigman D M,Cook M S,etal.Evidence from diatom-bound nitrogen isotopes for subarctic Pacific stratification during the last ice age and a link to North Pacific denitrification changes.Paleoceanography,2007,22(1):PA1215,doi:10.1029/2005PA001205.
10 Takahashi K.The Bering and Okhotsk Seas:modern and past paleoceanographic changes and gateway impact.Journal of Asian Science,1998,16(1):49—58.
11 Takahashi K,Fujitani N,Yanada M.Long-term monitoring of particle fluxes in the Bering Sea and the central subarctic Pacific Ocean,1990—2000.Progress in Oceanography,2002,55(1-2):95—112.
12 Gorbarenko SA,Wang P,Wang R,et al.Orbital and suborbital environmental changes in the southern Bering Sea during the last50 kyr.Palaeogeography,Palaeoclimatology,Palaeoecology,2010,286(1-2):97—106.
13 李丽,汪品先.大洋“生物泵”——海洋浮游植物生物标志物.海洋地质与第四纪地质,2004,24(4):73—79.
14 Zhao M,Huang C Y,Wang C C,et al.A millennial-scalesea-surface temperature record from the South China Sea(8°N)over the last150 kyr:Monsoon and sea-level influence.Palaeogeography,Palaeoclimatology,Palaeoecology,2006,236(1-2):39—55.
15 贺娟.南海北部(MD05-290柱样)26万年生物标志物记录的古环境变迁.博士学位论文,上海:同济大学,2008.
16 李文宝.南大洋南塔斯曼海2Ma以来的古海洋学记录及高、低纬海域的对比.博士学位论文,上海:同济大学,2010.
17 Martínez-Garcia A,Rosell-MeléA,GeibertW,et al.Links between iron supply,marine productivity,sea surface temperature,and CO2over the last1.1 Ma.Paleoceanography,2009,24(1),doi:10.1029/2008PA001657.
18 Volkman JK.A review of sterolmarkers formarine and terrigenous organic matter.Organic Geochemistry,1986,9(2):83—99.
19 Meyers P A.Organic geochemical proxies of paleoceanographic,paleolimnologic,and paleoclimatic processes.Organic Geochemistry,1997,27(5-6):213—250.
20 Volkman JK,Barrett SM,Dunstan G A,et al.Geochemical significance of the occurrence of dinosterol and other4-methyl sterols in amarine diatom.Organic Geochemistry,1993,20(1):7—15.
21 Marlowe IT,Brassell SC,Eglinton G,etal.Long chain unsaturated ketones and esters in living algae andmarine sediments.Organic Geochemistry,1984,6:135—141.
22 Marlowe IT,Green JC,Neal A C,et al.Long chain(n-C37-C39)alkenones in the Prymnesiophyceae:Distribution of alkenones and other lipids and their taxonomic significance.British Phycological Journal,1984,19(3):203—216.
23 Volkman JK,Barrett SM,Dunstan G A,etal.C30-C32alkyl diols and unsaturated alcohols inmicroalgae of the class Eustigmatophyceae.Organic Geochemistry,1992,18(1):131—138.
24 Volkman JK,Barrett SM,Blackburn S I.Eustigmatophytemicroalgae are potential sources of C29sterols,C22-C28n-alcohols and C28-C32n-alkyl diols in freshwater environments.Organic Geochemistry,1999,30(5):307—318.
25 南青云,李铁刚,陈金霞,等.南冲绳海槽7000年BP以来基于长链不饱和烯酮指标的古海洋生产力变化及其与气候的关系.第四纪研究,2008,28(3):482—490.
26 Eglinton T I,Eglinton G.Molecular proxies for paleoclimatology.Earth and Planetary Science Letters,2008,275(1-2):1—16.
27 黄小慧,王汝建,翦知湣,等.全新世冲绳海槽北部表层海水温度和初级生产力对黑潮变迁的响应.地球科学进展,2009,24(6):653—661.
28 丁玲,邢磊,赵美训.生物标志物重建浮游植物生产力及群落结构研究进展.地球科学进展,2010,25(9):981—989.
29 于培松,张海生,扈传昱,等.利用沉积生物标志物分析南极普里兹湾浮游植物群落结构变化.极地研究,2012,24(2):143—150.
30 Eglinton G,Hamilton R J.Leaf epicuticularwaxes.Science,1967,156(3780):1322—1335.
31 Meyers P A,Ishiwatari R.Lacustrine organic geochemistry an overview of indicators of organicmatter sourcesand diagenesis in lake sediments.Organic Geochemistry,1993,20(7):867—900.
32 贺娟,赵美训,李丽,等.南海北部MD05-2904沉积柱状样26万年以来表层海水温度及陆源生物标志物记录.科学通报,2008,53(11):1324—1331.
33 Ficken K J,LiB,Swain D L,etal.An n-alkane proxy for the sedimentary inputof submerged/floating freshwater aquaticmacrophytes.Organic Geochemistry,2000,31(7-8):745—749.
34 Meyers PA.Applications of organic geochemistry to paleolimnological reconstructions:a summary of examples from the Laurentian Great Lakes.Organic Geochemistry,2003,34(2):261—289.
35 MaffeiM.Chemotaxonomic significance of leafwax alkanes in the Gramineae.Biochemical Systematics and Ecology,1996,24(1):53—64.
36 Zhang Z,Zhao M,Eglinton G,etal.Leafwax lipidsas paleovegetational and paleoenvironmental proxies for the Chinese Loess Plateau over the last 170 kyr.Quaternary Science Reviews,2006,25(5-6):575—594.
37 卢冰,陈荣华,王自磐,等.长链烯酮及值在北极海洋古温度的应用研究.海洋学报,2001,23(5):49—57.
38 卢冰,陈荣华,王自磐,等.亚北极白令海近百年海洋环境变化——来自分子化石的证据.中国科学(D辑),2004,34(4):367—374.
39 卢冰,周怀阳,陈荣华,等.北极现代沉积物中正构烷烃的分子组合特征及其与不同纬度的海域对比.极地研究,2004,16(4):281—294.
40 张海生,潘建明,陈建芳,等.楚科奇海和白令海沉积物中的生物标志物及其生态环境响应.海洋地质与第四纪地质,2007,27(2):42—49.
41 FujiiM,Yamanaka Y,NojiriY,etal.Comparison of seasonal characteristics in biogeochemistry among the subarctic North Pacific stations described with a NEMURO——based marine ecosystem model.Ecological Modeling,2007,202(1-2):52—67.
42 Katsuki K,Takahashi K,Jordan RW,etal.Surface circulation changes based on fossil diatoms in the Bering Sea and thewestern subarctic Pacific.Kaiyo Monthly,2003,35:394—400.
43 Walsh J J,McRoy C P.Ecosystem analysis in the southeastern Bering Sea.Continental Shelf Research,1986,5(1-2):259—288.
44 Stabeno P J,Bond N A,Kachel N B,etal.On the temporal variability of the physical environmentover the south-eastern Bering Sea.Fisheries O-ceanography,2001,10(1):81—98.
45 Stockwell D A,Whitledge T E,Zeeman S I,etal.Anomalous conditions in the south-eastern Bering Sea,1997:Nutrients,phytoplankton and zooplankton.Fisheries Oceanography,2007,10(1):99—116.
46 Lovvorn JR,Cooper LW,Brooks M L,et al.Organicmatter pathways to zooplankton and benthos under pack ice in late winter and open water in late summer in the north-central Bering Sea.Marine Ecology Progress Series,2005,291:135—150.
47 Alexander V,Niebauer H J.Oceanography of the eastern Bering Sea ice-edge zone in spring.Limnology and Oceanography,1981,26(6):1111—1125.
48 Overland JE,Pease C H.Cyclone climatology of the Bering Sea and its relation to sea ice extent.Monthly Weather Review,1982,110(1):5—13.
49 Nagashima K,Asahara Y,Takeuchi F,etal.Contribution of detritalmaterials from the Yukon River to the continental shelf sediments of the Bering Sea based on the electron spin resonance signal intensity and crystallinity of quartz.Deep-Sea ResearchⅡ,2012,61-64:145—154.
50 Niebauer H J,Schell DM.Physical environmentof the Bering Sea population//Burns JJ,Montague JJ,Cowles C J.The Bowhead Whale.The Society for Marine Mammalogy,Special Publication No.2.Seattle,Washington,1993:23—43.
51 Roden G I.On river discharge into the northeastern Pacific Ocean and the Bering Sea.Journal of Geophysical Research,1967,72(22):5613—5629.
52 Brabets TP,Wang BW,Meade R H.Environmental and hydrologic overview of the Yukon River Basin,Alaska and Canada.U.S.Geological Survey Water-Resources Investigations Report,2000:99—4204.
53 Inman D L,Nordstrom C E.On the tectonic andmorphologic classification of coasts.The Journal of Geology,1971,79(1):1—21.
54 Lisitsysn A P.Recent Sedimentation in the Bering Sea.Washington,DC:National Science Foundation,The Israel Program for Scientific Translations(translated from Russian),1969.
55 Hood DW.The Bering Sea//Ketchum B H.Estuaries and Enclosed Seas.New York:Elsevier,1983:337—373.
56 Sancetta C,Heusser L,Labeyrie L,etal.Wisconsin-Holocene paleoenvironmentof the Bering Sea:Evidence from diatoms,pollen,oxygen isotopes and clayminerals.Marine Geology,1984,62(1):55—68.
57 陈立奇.中国首次北极科学考察报告.北京:海洋出版社,2000:98—100.
58 Ratnayake N P,Suzuki N,Okada M,et al.The variations of stable carbon isotope ratio of land plant-derived n-alkanes in deep-sea sediments from the Bering Sea and the North Pacific Ocean during the last250,000 years.Chemical Geology,2006,228(4):197—208.
59 Caissie B E,Brigham-Grette J,Lawrence K T,et al.Last Glacial Maximum to Holocene sea surface conditions at Umnak Plateau,Bering Sea,as inferred from diatom,alkenone,and stable isotope records.Paleoceanography,2010,25(1),PA1206,doi:10.1029/2008PA001671
60 Fleming K,Johnston P,Zwartz D,etal.Refining the eustatic sea-level curve since the LastGlacialMaximum using far-and intermediate-field sites.Earth and Planetary Science Letters,1998,163(1-4):327—342.
61 Lambeck K,Yokoyama Y,Purcell T.Into and outof the LastGlacialMaximum:Sea-level changes during oxygen isotope stages3 and 2.Quaternary Science Reviews,2002,21(1-3):343—360.
62 Sancetta C.Use of semiquantitativemicrofossil data for paleoceanography.Geology,1979,7(2):88—92.
63 Katsuki K,Khim B K,Itaki T,et al.Land-sea linkage of Holocene paleoclimate on the Southern Bering Continental Shelf.The Holocene,2009,19(5):747—756.
64 Max L,Riethdorf JR,Tiedemann R,et al.Sea surface temperature variability and sea-ice extent in the subarctic northwest Pacific during the past 15 000 years.Paleoceanography,2012,27,PA3213,doi:10.1029/2012PA002292.
65 Crockford S J,Frederick SG.Sea ice expansion in the Bering Sea during the Neoglacial:evidence from archaeozoology.The Holocene,2007,17(6):699—706.
66 Laskar J,Robutel P,Joutel F,etal.A long term numericalsolution for the insolation quantitiesof the Earth.Astronomy&Astrophysics,2004,428(1):261—285,doi:10.1051/0004-6361:20041335.
67 Vinther BM,Clausen H B,Johnsen SJ,etal.A synchronized dating of three Greenland ice cores throughout the Holocene.Journal of Geophysical Research,2006,111,D13102,doi:10.1029/2005JD006921.
68 陈荣华,盂翊,华棣,等.楚科奇海与白令海表层沉积物中的钙质和硅质微体化石研究.海洋地质与第四纪地质,2001,21(4):25—30.69 杨清良,林更铭,林茂,等.楚科奇海和白令海浮游植物的种类组成与分布.极地研究,2002,14(2):113—125.
70 邢磊,赵美训,张海龙,等.冲绳海槽中部过去15ka来浮游植物生产力和种群结构变化的生物标志物重建.科学通报,2008,53(12):1448—1455.
71 胡建芳,彭平安,贾国东,等.三万年来南沙海区古环境重建:生物标志物定量与单体碳同位素研究.沉积学报,2003,21(2):211—218.
72 Bird M I,Summons R E,Gagan M K,et al.Terrestrial vegetation change inferred from n-alkanesδ13C analysis in the marine environment.Geochimica et Cosmochimica Acta,1995,59(13):2853—2857.
73 Pelejero C,Kienast M,Wang L,et al.The flooding of Sundaland during the last deglaciation:imprints in hemipelagic sediments from the southern South China Sea.Earth and Planetary Science Letters,1999,171(4):661—671.
74 Gagosian R B,Peltzer E T,Zafiriou O C.Atmospheric transport of continentally derived lipids to the tropical North Pacific.Nature,1981,291(5813):312—314.
75 Huang Y,Collister JW,Chester R,etal.Molecular andδ13Cmappingofeolian inputof organic compounds intomarine sediments in the Northeastern Atlantic//Фygard K.Organic Geochemistry.Oslo:Falch Hurtigtrykk,1993:523—528.
76 黄小慧,王汝建,翦知湣.全新世冲绳海槽北部陆源输入物变化及其古气候意义.海洋地质与第四纪地质,2009,29(5):73—82.
77 Bard E,Hamelin B,Arold M,et al.Deglacial sea-level record from Tahiti corals and the timing of globalmeltwater discharge.Nature,1996,382(6588):241—244.
78 Chen Y G,Liu T K.Sea level changes in the last several thousand years,Penghu Islands,Taiwan Strait.Quaternary Research,1996,45(3):254—262.
79 Waelbroeck C,Labeyrie L,Michel E,etal.Sea-level and deep water temperature changes derived from benthic foraminifera isotopic records.Quaternary Science Reviews,2002,21(1-3):295—305.
80 Hopkins D M.Quaternary Marine Transgressions in Alaska.The Bering Land Bridge.Stanford:Stanford University Press,1967:47—90.
81 Alley R B,Mayewski P A,Sowers T,et al.Holocene climatic instability:A prominent widespread event 8.2 ka ago.Geology,1997,25(6):483—486.
82 Mayewski PA,Meeker L D,Twickler M S,et al.Major features and forcing of high-latitude northern hemisphere atmospheric circulation using a 110 000-year-long glaciochemical series.Journal of Geophysical,1997,102(12):26345—26365.
83 Hu F S,Brubaker L B,Anderson PM.Boreal ecosystem development in the northwestern Alaska range since11,000 yr BP.Quaternary Research,1996,45(2):188—201.
84 Lozhkin A V,Anderson PM,EisnerW P,et al.Late Quaternary Lacustrine pollen records from the southwestern Beringia.Quaternary Research,1993,39(3):314—324.
85 刘焱光,石学法,吕海龙.日本海、鄂霍次克海和白令海的古海洋学研究进展.海洋科学进展,2004,22(4):519—530.
86 叶春江,赵可夫.高等植物大叶藻研究进展及其对海洋沉水生活的适应.植物学通报,2002,19(2):184—193.
87 Nejrup L B,Pedersen M F.Effects of salinity and water temperature on the ecological performance of Zosteramarina.Aquatic Botany,2008,88(3):239—246.
88 Anderson PM,Reanier R E,Brubaker L B.Late Quaternary vegetational history of the Black River region in northeastern Alaska.Canadian Journal of Earth Sciences,1988,25(1):84—94.
89 Anderson PM,Brubaker L B.Vegetation history of north-central Alaska:amapped summary of late-Quaternary pollen data.Quaternary Science Reviews,1994,13(1):71—92.
90 Ritchie JC,Cwynar L C,Spear RW.Evidence from north-west Canada for an early Holocene Milankovitch thermalmaximum.Nature,1983,305(5930):126—128.
91 Barnosky CW,Anderson PM,Bartlein P J.The northwestern USduring deglaciation:vegetational history and paleoclimatic implications.North A-merica and Adjacent Oceans during the Last Deglaciation,1987,3:289—321.
