α-硫辛酸和乙酰左旋肉碱改善炎症细胞因子介导胰岛细胞功能障碍的作用
2014-03-08张召锋顾娇娇蔡夏夏北京大学公共卫生学院营养与食品卫生学系食品安全毒理学研究与评价北京市重点实验室北京100191
张召锋,顾娇娇,鲍 雷,蔡夏夏,李 勇*(北京大学公共卫生学院营养与食品卫生学系,食品安全毒理学研究与评价北京市重点实验室,北京 100191)
α-硫辛酸和乙酰左旋肉碱改善炎症细胞因子介导胰岛细胞功能障碍的作用
张召锋,顾娇娇,鲍 雷,蔡夏夏,李 勇*
(北京大学公共卫生学院营养与食品卫生学系,食品安全毒理学研究与评价北京市重点实验室,北京 100191)
目的:探讨α-硫辛酸(α-lipoic acid,LA)和乙酰左旋肉碱(acetyl-L-carnitine,ALC)改善炎症细胞因子介导胰岛细胞功能障碍的效果并探讨机制。方法:大鼠胰岛素瘤RIN-m5f细胞用肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、白细胞介素-1β(interleukin-1β, IL-1β)和γ-干扰素(interferon-γ,IFN-γ)联合作用(TII)48 h造成损伤模型。LA和ALC干预48 h后噻唑蓝法检测胰岛细胞的活力情况,荧光细胞技术法检测细胞的形态学,流式细胞仪检测细胞活性氧(reactive oxygen species,ROS)表达水平,放射免疫法检测胰岛素分泌情况,Western blotting检测细胞凋亡相关和胰岛素分泌相关蛋白表达情况。结果:TII作用48 h可使RIN-m5f细胞活力明显下降,凋亡增加;并降低基础状态下和高糖刺激状态下胰岛素分泌水平;增加ROS水平,增加一氧化氮合酶(nitric oxide synthase,NOS)活性,增加一氧化氮(nitrogen monoxide,NO)水平,促进NF-κB向细胞核转位;TII还可增加RIN-m5f细胞内促凋亡蛋白Bax、Caspase-3表达,抑制抗凋亡蛋白I-κB、Bcl-2表达,增加线粒体细胞色素c释放。而LA、ALC可改善TII诱导的RIN-m5f细胞凋亡,提高基础状态和高糖刺激状态下胰岛素分泌水平;抑制NF-κB向细胞核转位、降低细胞NO水平;降低Bax、Caspase-3表达,增加抗凋亡蛋白I-κB、Bcl-2表达,抑制线粒体细胞色素c释放;LA与ALC联合作用效果强于单独作用。结论:炎症细胞因子作用48 h可通过ROS-cytochrome c-NF-κB-NOS-NO通路最终引起胰岛β细胞的凋亡,进而影响胰岛素分泌;LA和ALC联用可抑制炎症细胞因子诱导的胰岛细胞凋亡,促进胰岛素分泌。
炎症细胞因子;胰岛细胞功能障碍;α-硫辛酸;乙酰左旋肉碱
研究证实,不论是1型还是2型糖尿病,动物和人群体内均存在炎症细胞因子的过度表达,在糖尿病发病过程中发挥重要的作用,其中影响最大的3 种炎症细胞因子是肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、白细胞介素-1β(interleukin-1β,IL-1β)和γ-干扰素(interferon-γ,IFN-γ)[1-3]。已证实目前临床上的降糖药物 (格列酮类、α-糖苷酶抑制剂)均有一定抗炎作用,能部分改善胰岛细胞功能,但其毒副作用较大;而天然药物具有作用持久、缓和、毒副作用小、从整体水平上防治和治疗糖尿病等特点。α-硫辛酸(α-lipoic acid,LA)属于类维生素物质,是线粒体脱氢酶的天然辅助因子,能清除除超氧阴离子自由基(O2-·)、羧自由基(ROO·)以外的其他自由基和活性氧,从而发挥抗氧化作用,已在糖尿病治疗研究中取得了较大进展[4-5]。乙酰左旋肉碱(acetyl-L-carnitine,ALC)是L-肉碱的一种天然形态,存在于体内的一种自然物质,尤其在肌肉、大脑及精子中含量特别丰富。ALC能够调节细胞线粒体内酰基比率,将短链脂肪酰基从线粒体膜内运到膜外,维持机体生理平衡[6]。研究表明,LA和ALC联用对油酸处理的胰岛MIN6细胞有一定的保护作用[7]。然而ALC单独作用或与LA联合作用对炎症细胞因子介导的胰岛细胞功能障碍是否有效果还未见报道。因此本实验拟通过大鼠胰岛素瘤RIN-m5f细胞的培养,探讨采用LA、ALC干预对炎症细胞因子介导胰岛细胞功能障碍的作用,并进一步探讨其作用机制。
1 材料与方法
1.1 材料与试剂
大鼠胰岛素瘤RIN-m5f细胞株由北京大学基础医学院生物化学与分子生物学系周春燕教授惠赠。
DMEM细胞培养基、胎牛血清(fetal bovine serum,FBS) 美国Gibco公司;anti-β-actin、anti-Bcl-2、anti-Bax、anti-NF-κB、anti-I-κB、anti-Caspase-3、anticytochorome c抗体、辣根过氧化物酶标记的二抗 美国Santa Cruz公司;IFN-γ、IL-1β、TNF-α 美国Peprotech公司;活性氧(reactive oxygen species,ROS)、一氧化氮(nitrogen monoxide,NO)、一氧化氮合酶(nitric oxide synthase,NOS)试剂盒 北京普利莱基因技术有限公司;噻唑蓝(3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide,MTT)、LA、ALC美国Sigma公司。
1.2 仪器与设备
二氧化碳培养箱 日本SANyO公司;CK40-SL倒置显微镜 日本Olympus公司;Bio-Rad 550型酶联免疫检测仪、蛋白质电转移装置 美国Bio-Rad公司;UV762型分光光度计 上海精密科学仪器有限公司;Adventurer™ 通用型分析天平 美国OHAUS公司。
1.3 方法
1.3.1 细胞培养
大鼠RIN-m5f胰岛素瘤细胞用含10% FBS及2 mol/L谷氨酰胺的高糖DMEM培养基培养,置于二氧化碳培养箱(5% CO2,37 ℃)中孵育。待细胞生长至约80%融合,将其用含0.2% FBS的DMEM培养基饥饿过夜,各种不同因素处理细胞。
1.3.2 实验分组
分为5组:1)正常对照组:胰岛细胞不作任何处理;2)炎性细胞因子损伤组:1000 U/mL IFN-γ+ 2 ng/mL IL-1β+10 ng/mL TNF-α组(简称TII组);3)LA干预组(1、10、100 μmol/L);4)ALC干预组(1、10、100 μmol/L);5)LA+ALC联合干预组(10 μmol/L LA+10 μmol/L ALC;100 μmol/L LA+100 μmol/L ALC)。LA、ALC分别预处理2 h,细胞培养48 h。
1.3.3 MTT比色法检测细胞活力
在各种因素处理后,将10 μL的MTT储备液(5 mg/mL)加到每孔,然后将培养板置于37 ℃培养4 h。孵育结束后,弃去各孔培养液,每孔加入100 μL二甲基亚砜,振荡均匀后,在570 nm波长处测定吸光度。每组有6 个复孔,以无细胞孔的吸光度作为空白本底,计算各组细胞的抑制率。
1.3.4 荧光染色法观察细胞形态
细胞以50%~80%密度接种于事先放入爬片的6 孔培养板中,经过相应处理后,去除上清用质量分数为4%多聚甲醛0.5 mL重悬细胞,4℃过夜;PBS洗涤2 次,每次3 min;加入0.5 mL Hoechst 33258染色5 min;用PBS洗涤2次,每次3 min,滴加一滴抗荧光淬灭封片液于载玻片上,盖上贴有细胞的盖玻片,使细胞接触封片液,荧光显微镜检测呈蓝色的细胞核。激发波长在350 nm左右,发射波长在460 nm左右。
1.3.5 基础胰岛素检测和胰岛素释放实验
将培养至对数生长期的RIN-m5f胰岛细胞以104/孔密度接种于96 孔板,经相应处理48 h后,去除培养基,用含2.8、17.6 mmol/L葡萄糖的无酚红培养基200 μL分别刺激1 h后,收集细胞上清液,用I125胰岛素放射免疫分析药盒检测基础状态下和高糖状态下胰岛素释放水平。
1.3.6 细胞NO的检测
各种因素处理后,收集细胞上清液,用NO测定试剂盒检测NO水平,同时各实验组进行蛋白定量,结果以各组NO浓度/蛋白含量比值表示,具体方法:按50 μL/孔,在96孔板中加入标准品及样品。样品为细胞裂解液,各孔加入室温Griess ReagentⅠ 50 μL/孔;按50 μL/孔,各孔加入室温Griess Reagent Ⅱ;540 nm波长处测定吸光度。
1.3.7 细胞NOS的检测
细胞经刺激后,吸尽培养液,加入100 μL NOS检测缓冲液,再加入100 μL检测反应液,轻轻混匀,37 ℃细胞培养箱内孵育30 min。直接取该96孔板用荧光酶标仪检测。以没有细胞的孔为空白对照,激发波长为495 nm,发射波长为515 nm。NOS相对活力的计算:以未刺激样品中NOS的活力为1,刺激后样品中NOS相对活力按下式计算。

式中:RFU为实际测定得到的相对荧光强度。
1.3.8 ROS检测
按照活性氧检测试剂盒,利用荧光探针2,7-二氯荧光黄双乙酸盐(2’,7’-dichlorofluorescein diace-tate,DCFH-DA)进行活性氧检测,最后用流式细胞仪检测荧光强度。
1.3.9 Western blotting实验
收集细胞,加入冰预冷的裂解液并置于冰上裂解30 min,于4 ℃、12 000 r/min离心5 min,收集上清液。用2,2-联喹啉-4,4-二甲酸二钠(bicinchoninic acid,BCA)法定量蛋白浓度。蛋白样品经过聚丙烯酰胺凝胶后,被转到聚偏氟乙烯(polyvinylidene fluoride,PVDF)膜上。用含5%的脱脂奶粉的封闭液4 ℃过夜封闭PVDF膜,并以相应一抗杂交,洗膜后再以辣根过氧化物酶标记的二抗(1∶5 000)孵育。膜经清洗后,用增强化学发光试剂盒进行自显影。采用β-actin作为内对照。用Bio-Rad图像分析系统对条带进行扫描,然后用Quantity One软件进行分析。
1.4 统计分析
2 结果与分析
2.1 LA、ALC对胰岛细胞存活率的影响

图1 MTT法检测LA和ALC对TII处理的胰岛细胞抑制率的影响Fig.1 Effects of LA and ALC on the viability of cytokines-treated RIN-m5f cells by MTT assay
由图1可知,当TII刺激细胞48 h后,RIN-m5f细胞死亡明显增多,与对照组比较,细胞被抑制40.3%,有极显著性差异(P<0.01);而LA和ALC单独或联合干预后,TII的细胞抑制作用明显减轻(P<0.05或P<0.01),并且LA的作用呈剂量反应关系,LA和ALC联合作用比两者单独作用强,说明二者存在协同效应。
2.2 LA、ALC对胰岛细胞形态的影响

图2 LA、ALC对TII处理的胰岛细胞形态的影响(×2 000)Fig.2 Visualization of RIN-m5f cells under fluorescence microscopy after being stained with Hoechst 33258 (× 2 000)
由图2可知,荧光显微镜观察可见正常对照组的细胞核边界规则整齐,呈弥散均匀荧光;TII组细胞核致密浓染,呈碎块状,荧光强度比正常细胞高,核膜皱缩,呈现典型的凋亡形态学改变;而LA和ALC处理组的细胞核与TII组相比,凋亡细胞明显减少,对凋亡细胞的形态变化也有所改善,说明TII能诱导细胞凋亡而LA和ALC能抑制TII诱导的胰岛细胞凋亡。
JDR-30DB绞车是一种交流变频控制的齿轮传动单轴绞车,主要由大功率交流变频电动机、小功率送钻电机(含减速机)、大减速箱、大联轴器、小联轴器、气胎离合器、液压盘刹、滚筒轴、绞车架、气控系统、润滑系统、控制箱等单元部件组成。绞车结构见图1所示。
2.3 LA、ALC对胰岛β细胞胰岛素分泌能力的影响
与正常对照组相比,TII组胰岛细胞基础状态下和高糖刺激下的胰岛素水平降低,刺激指数(stimulating index,SI):高糖刺激状态胰岛素与基础状态胰岛素的比值也下降,胰岛素分泌存在统计学差异(P<0.05)。与TII组比较,当预先给予LA、ALC后,可以发现细胞在基础状态下和葡萄糖刺激下胰岛素分泌能力都有不同程度的改善(P<0.05或P<0.01),并且呈现剂量依赖性,同时刺激指数也有一定的增加,结果见表1。
表1 LA和ALC对RIN-m5f细胞胰岛素分泌的影响Table 1 Effects of LA and ALC on insulin secretion in RIN-m5f cells

表1 LA和ALC对RIN-m5f细胞胰岛素分泌的影响Table 1 Effects of LA and ALC on insulin secretion in RIN-m5f cells
注:#.与对照组相比,差异显著(P<0.05); ##.与对照组相比,差异极显著(P<0.01)。下同。
组别胰岛素分泌水平/(ng/mL)刺激指数2.8 mmol/L葡萄糖16.7 mmol/L葡萄糖对照组47.16±9.43135.70±13.092.88 TII32.34±6.63#60.26±14.58##1.85#LA1036.24±5.2373.20±11.532.02 LA10040.37±8.18*97.69±10.38**2.42*ALC1034.13±5.6368.60±12.642.01 ALC10038.75±8.23*87.96±9.96*2.27*LA10+ALC1039.76±6.48*93.83±16.47**2.36*LA100+ALC10046.23±8.34*125.74±16.52**2.72*
2.4 LA、ALC对胰岛RIN-m5f细胞NO和NOS的影响

图3 LA、ALC对TII处理的胰岛RIN-m5f细胞NO含量和NOS活力的影响Fig.3 Effects of LA and ALC on TII-induced NO production and NOS expression in RIN-m5f cells
由图3可知,与对照组相比,TⅡ刺激细胞48 h后RIN-m5f细胞分泌NO显著增加,NOS表达也增加(P<0.01)。经LA和ALC预处理后,细胞内NO水平较TII组均明显减少(P<0.01),并呈现一定剂量依赖性,提示LA和ALC能够减轻TII诱导的β细胞凋亡同时可减少细胞内NO的水平。同时LA(100 μmol/L)、两个LA+ ALC组预处理明显地抑制TII诱导RIN-m5f细胞NOS表达水平的增加(P<0.01或P<0.05)。
2.5 LA、ALC对胰岛细胞ROS的影响

图4 LA、ALC对胰岛细胞ROS的影响Fig.4 Effects of LA and ALC on ROS levels in RIN-m5f cells
由图4可知,与对照组比较,TII组胰岛RIN-m5f细胞DCFH-DA的荧光强度显著增加(P<0.05)。当预先给予LA和ALC后,细胞内DCFH-DA的荧光强度显著减少(P<0.01)。

图5 LA、ALC对TII诱导的胰岛细胞凋亡相关蛋白的影响Fig.5 Effects of LA and ALC on apoptosis-related protein of RIN-m5f cells treated by TII for 48 h
由图5可知,RIN-m5f细胞经炎症细胞因子处理48 h后,凋亡蛋白的表达明显上调,抗凋亡蛋白表达下调,Bax、Caspase-3蛋白表达分别增加3.25、2.42倍,Bcl-2、I-κB蛋白表达较对照组分别降低58.3%、52.6%,与对照组比较差异具有显著性(P<0.05)。而分别加入LA和ALC后上述凋亡相关蛋白的表达有明显的改善。此外,与对照组对比,炎症细胞因子对总的NF-κB表达没有影响,但TII组细胞核蛋白NF-κB表达增加了1.5倍,而胞浆蛋白NF-κB表达减少了50%(P<0.05),说明炎症细胞因子能促进NF-κB由细胞浆向细胞核转位,而加入LA+ALC后NF-κB转位至细胞核的量均明显减少(P<0.05)。
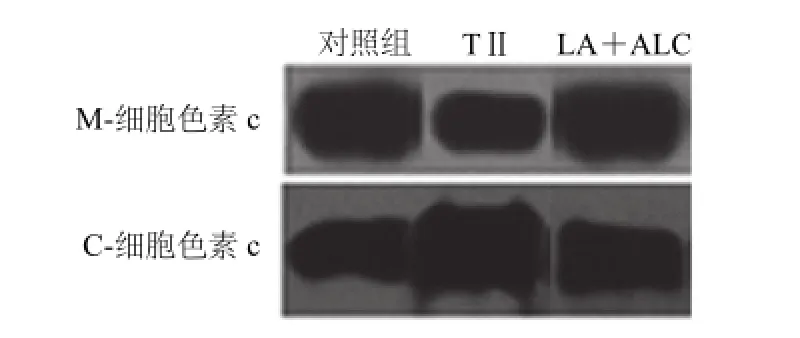
图6 LA、ALC对TII诱导的胰岛细胞线粒体细胞色素c释放的影响Fig.6 Effects of LA and ALC on cytochrome c release of RIN-m5f cells treated by TII for 48 h
由图6可知,RIN-m5f细胞受TII刺激后,线粒体内的细胞色素c释放进入胞浆中,与对照组相比其胞浆细胞色素c的浓度增加了近1倍,线粒体中细胞色素c的浓度降低了50%,显著低于对照组(P<0.05);加入LA+ALC预处理后,线粒体内细胞色素c表达量增加。细胞浆中细胞色素c表达量显著下降(P<0.05)。
3 讨 论
近年来,炎症细胞因子在1型和2型糖尿病的病理生理机制中所起的作用一直是关注的热点。本实验发现TNF-α、IFN-γ和IL-1β联合使用可导致胰岛RIN-m5f细胞凋亡。TNF-α、IFN-γ和IL-1β均为促炎性细胞因子,它们对胰岛β细胞的炎症性损害已见报道[8-9],IFN-γ、IL-1β、TNF-α对β细胞生存能力的影响可能具有不同的机制,这说明3 种炎症细胞因子在促进胰岛细胞凋亡过程中起到了协同作用。凋亡对胰岛细胞产生的直接影响就是胰岛素分泌障碍,本实验发现TNF-α、IFN-γ、IL-1β联合作用48 h后,RIN-m5f细胞的基础胰岛素分泌和高糖状态下胰岛素分泌都减少,刺激指数也明显下降,这与以前文献报道是一致的[10]。本实验还发现LA和ALC预处理能抑制TII诱导的胰岛细胞凋亡,并且LA表现为一定的剂量依赖性,同时LA和ALC联合作用比单独作用效果更加明显,说明二者存在协同效应。加入LA和ALC,胰岛细胞基础状态下和葡萄糖刺激状态下胰岛素分泌量明显增加。再次证明LA和ALC联用对TII引起的胰岛β细胞凋亡具有明显的拮抗作用。
激活NF-κB信号途径后增加β细胞凋亡,被认为是1型和2型糖尿病的共同发病机制[11-12]。静息细胞中NF-κB与其抑制蛋白(I-κB)结合存在于胞浆中处失活状态。炎症细胞因子刺激可诱导I-κB磷酸化而使NF-κB活化,并转位于胞核内,结合至DNA上的相应位点,调控下游相关靶基因的转录表达,包括NOS、IL等,从而促进NO产生而损伤β细胞[13]。本实验发现TNF-α、 IFN-γ和IL-1β联合作用RIN-m5f细胞48 h后,I-κB蛋白表达量细胞下调,但对NF-κB总量影响不大,却能明显增加NF-κB转位至细胞核内的量进而诱导凋亡。为了进一步阐明LA和ALC改善胰岛细胞凋亡的机制,通过分离细胞核蛋白检测了NF-κB的转位情况,发现LA+ALC能减少NF-κB转位至细胞核内的量,从而改善TII诱导的胰岛细胞凋亡。
活化巨噬细胞及T细胞产生的NO、氧自由基等为糖尿病的主要致病因素,以前的文献都报道了炎症细胞因子高表达可能主要通过诱导NOS表达,促进NO产生而损伤β细胞[14]。本实验结果证实:联合使用3 种炎症细胞因子作用48 h可显著增强RIN-m5f细胞内NOS的活性,并使培养上清液中NO的含量显著增加,ROS水平表达增加。运用LA、ALC干预后胰岛细胞NOS活性和NO水平均明显低于炎症细胞因子损伤组。实验结果表明LA、ALC对抗上述炎症细胞因子诱导胰岛β细胞凋亡,抑制NOS活性,减少细胞内NO生成,从而减轻NO对β细胞的毒性作用也是其保护细胞、促进胰岛细胞胰岛素分泌的重要机制。
B细胞淋巴瘤蛋白家族(Bcl家族)是一类原癌凋亡基因家族,在程序性细胞死亡过程中起关键作用,Bcl-2和Bax分别是Bcl家族中最主要的抑制凋亡和促进凋亡蛋白[15]。在本实验中,TNF-α、IFN-γ、IL-1β联合作用RIN-m5f细胞48 h后Bcl-2蛋白表达降低,Bax蛋白表达增强;而LA和ALC能升高胰岛细胞Bcl-2表达,降低Bax表达,提示Bcl-2基因家族既是TNF-α、IFN-γ和IL-1β诱导胰岛细胞凋亡过程中的一种调节因子,也是LA和ALC发挥其保护作用的一个途径。
近年来人们对凋亡的认识已经从细胞核的改变决定凋亡,发展为重视线粒体,因为它构成了细胞存亡的控制中心。细胞色素c是哺乳动物细胞凋亡信号传导过程的关键因素,通常情况下细胞色素c是一个定位于线粒体膜间隙的相对分子质量为14.5 kD的水溶性蛋白质,电稳定性地结合于线粒体内膜,不能通过外膜。凋亡过程中细胞色素c通过线粒体外膜释放,导致Caspase-3激活,并进一步激活下游的Caspases,引起一系列瀑布式反应,最后致使细胞凋亡[16-18]。细胞色素c释放还可通过引起线粒体呼吸链电子传递障碍,使ROS产生增加,介导细胞凋亡。本实验发现:TNF-α、IFN-γ和IL-1β联合作用RIN-m5f细胞48 h后,线粒体细胞色素C释放明显增加,Caspase-3蛋白表达明显升高。这说明炎症细胞因子作用胰岛RIN-m5f细胞48 h能够引起胰岛细胞线粒体凋亡。此外,LA+ALC能够明显降低胰岛细胞ROS水平,抑制线粒体细胞色素c的释放,从而保护胰岛细胞线粒体不被炎症细胞因子破坏,这说明LA、ALC可保护线粒体结构和功能也是LA、ALC改善胰岛细胞凋亡的主要途径。
总之,炎症细胞因子可通过ROS-cytochrome c释放-ROS-NF-κB-NOS-NO通路最终引起胰岛β细胞的凋亡,进而影响胰岛素分泌;而LA和ALC联用可抑制炎症细胞因子诱导的胰岛细胞凋亡,促进胰岛素分泌。
[1] HOTAMISLIGIL G S. Inflammation and metabolic disorders[J]. Nature, 2006, 444: 860-867.
[2] GUEST C B, PARK M J, JOHNSON D R, et al. The implication of proinflammatory cytokines in type 2 diabetes[J]. Frontiers in Bioscience, 2008, 13: 5187-5194.
[3] WILLCOX A, RICHARDSON S J, BONE A J, et al. Analysis of islet inflammation in human type 1 diabetes[J]. Clinical and Experimental Immunology, 2009, 155(2): 173-181.
[4] LEE B W, KWON S J, CHAE H Y, et al. Dose-related cytoprotective effect of alpha-lipoic acid on hydrogen peroxide-induced oxidative stress to pancreatic beta cells[J]. Free Radical Research, 2009, 43(1): 68-77.
[5] BUDIN S B, OTHMAN F, LOUIS S R, et al. Effect of alpha lipoic acid on oxidative stress and vascular wall of diabetic rats[J]. Romanian Journal of Morphology Embryology and Physiology, 2009, 50(1): 23-30.
[6] LOWITT S, MALONE J I, SALEM A F, et al. Acetyl-L-carnitine corrects the altered peripheral nerve function of experimental diabetes[J]. Metabolism, 1995, 44(5): 677-680.
[7] SHEN W, LIU K, TIAN C, et al. Protective effects of R-alphalipoic acid and acetyl-L-carnitine in MIN6 and isolated rat islet cells chronically exposed to oleic acid[J]. Journal of Cell Biochemistry, 2008, 104(4): 1232-1243.
[8] FURSTOVA V, KOPSKA T, JAMES R F, et al. Comparison of the effect of individual saturated and unsaturated fatty acids on cell growth and death induction in the human pancreatic beta-cell line NES2Y[J]. Life Sciences, 2008, 82(13/14): 684-691.
[9] MACFARLANE W M, O’BRIEN R E, BARNES P D, et al. Sulfonylurea receptor 1 and Kir6.2 expression in the novel human insulin-secreting cell line NES2Y[J]. Diabetes, 2000, 49(6): 953-960.
[10] WHITE M F. The IRS-signaling system: a network of docking proteins that mediate insulin and cytokine action[J]. Recent Progress in Hormone Research, 1998, 53: 119-138.
[11] CHEN F. Is NF-kappaB a culprit in type 2 diabetes?[J]. Biochemical and Biophysical Research Communications, 2005, 332(1): 1-3.
[12] FRIER B C, NOBLE E G, LOCKE M. Diabetes-induced atrophy is associated with a muscle-specific alteration in NF-kappaB activation and expression[J]. Cell Stress and Chaperones, 2008, 13(3): 287-296.
[13] SCHIEKOFER S, GALASSO G, ANDRASSY M, et al. Glucose control with insulin results in reduction of NF-kappaB-binding activity in mononuclear blood cells of patients with recently manifested type 1 diabetes[J]. Diabetes Obesity and Metabolism, 2006, 8(5): 473-482.
[14] PAPACCIO G, GRAZIANO A, D’AQUINO R, et al. A biphasic role of nuclear transcription factor (NF)-kappaB in the islet betacell apoptosis induced by interleukin (IL)-1beta[J]. Journal of Cell Physiology, 2005, 204(1): 124-130.
[15] BERNEY T, MOLANO R D, CATTAN P, et al. Endotoxin-mediated delayed islet graft function is associated with increased intra-islet cytokine production and islet cell apoptosis[J]. Transplantation, 2001, 71(1): 125-132.
[16] ZOU Hua, LI Yuchen, LIU Xuesong, et al. An APAF-1.cytochrome c multimeric complex is a functional apoptosome that activates procaspase-9[J]. The Journal of Biological Chemistry, 1999, 274(17): 11549-11456.
[17] HU Yuanming, BENEDICT M A, DING Liyun, et al. Role of cytochrome c and dATP/ATP hydrolysis in Apaf-1-mediated caspase-9 activation and apoptosis[J]. The EMBO Journal, 1999, 18(13): 3586-3595.
[18] PANDEY P, SALEH A, NAKAZAWA A, et al. Negative regulation of cytochrome c-mediated oligomerization of Apaf-1 and activation of procaspase-9 by heat shock protein 90[J]. The EMBO Journal, 2000, 19(16): 4310-4322.
Effects of α-Lipoic Acid and Acetyl-L-Carnitine on Cytokines-Induced Islet Cell Dysfunction
ZHANG ZHAO-feng, GU Jiao-jiao, BAO Lei, CAI Xia-xia, LI Yong*
(Beijing Key Laboratory of Toxicological Research and Risk Assessment for Food Safety, Department of Nutrition and Food Hygiene, School of Public Health, Peking University, Beijing 100191, China)
Objective: To explore the effects of α-lipoic acid (LA) and acetyl-L-carnitine (ALC) on islet cell dysfunction. Methods: LA and ALC were co-incubated with a combination of three cytokines, i.e. tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β) and interferon-γ (IFN-γ) for 48 h, to establish TII-treated rat insulinoma cell lines (Rin-m5F cells). Cell viability was detected with MTT assay and cell morphology was examined with the imunofluorescence technique. Reactive oxygen species (ROS) were measured with fl ow cytometry. The expression levels of apoptotic-related proteins including Bax, Caspase-3, NF-κB and Bcl-2 were examined via Western blotting analysis. Insulin level was measured through radioimmunoassay (RIA). Results: Treatment of rats with TII for 48 h significantly decreased RIN-m5f cells viability, and basal and glucose-stimulated insulin secretion. TII signif i cantly increased ROS and nitrite oxide (NO) levels, promoted NF-κB translocation into nucleus. TII increased NF-κB, Bax and Caspase-3 expression, inhibited I-κB and Bcl-2 expression. In addition, TII increased cytochrome c release from mitochondira. A combination of LA and ALC was seen to facilitate improvement of TII-induced islet cell dysfunction, increase RIN-m5f cells viability and basal and glucose-stimulated insulin secretion, decrease ROS and NO levels, inhibit NF-κB translocation into nucleus, decrease NF-κB, Bax and Caspase-3 expression and increase I-κB and Bcl-2 expression. LA+ALC decreased cytochrome c release from mitochondria. Conclusions: TII can induce RIN-m5f cells dysfunction and inhibit insulin secretion viaROS-mitochondria- apoptosis-NF-κB-NOS-NO pathway. LA+ALC can facilitate RIN-m5f cells function via ROS-mitochondria apoptosis-NF-κB-NOS-NO pathway.
cytokine; islet cell dysfunction; α-lipoic acid; acetyl-L-carnitine
151.2
A
1002-6630(2014)15-0232-06
10.7506/spkx1002-6630-201415047
2013-10-02
教育部博士点基金项目(20100001110060)
张召锋 (1978—),男,讲师,博士,研究方向为营养与疾病。E-mail:zhangzhaofeng@126.com
*通信作者:李勇(1958—),男,教授,博士,研究方向为营养与疾病。E-mail:liyong@bjmu.edu.cn
