应用RACE 技术扩增柞蚕Lectin基因及原核表达载体构建
2014-01-28林坤章邱建烽尹志亮刘朝良朱保建
林坤章 邱建烽 尹志亮 刘朝良 朱保建
(1.安徽农业大学生命科学学院 223096;2.江西省蚕桑茶叶研究所)
昆虫在适应外界环境的过程中,已经形成一套自身的免疫系统,包括体液免疫和细胞免疫,而这些免疫反应是由模式识别受体调节的先天免疫识别所引发的[1~2]。体液免疫系统能产生包括凝集素、抗菌肽等一系列的抗菌多肽[3]。凝集素是一种重要的模式识别受体,在宿主抵御和识别外界病原物方面扮演着重要的角色,已成为近几年来的研究热题之一。C-型凝集素(C -type lectins)是一种钙依赖的糖结合蛋白,属于凝集素的重要成员之一,在机体的免疫防御、控制糖蛋白的生物合成等方面具有重要的功能[4~5]。它们一般含有1 个或多个由约130氨基酸残基组成的糖基识别域[6]。近几年来,一些新的凝集素基因陆续被报道,它们具有参与细菌的识别与清除、调节血液封装和黑化等多方面的功能也一度成为研究的热点[7~10]。柞蚕(Antheraea pernyi)是我国重要的经济昆虫之一,关于柞蚕抗病能力和防御系统相关功能基因的关系是值得研究的课题。克隆获得柞蚕凝集素基因对今后研究柞蚕的免疫反应机制,提高柞蚕的抗病能力具有一定的意义。
1 材料与方法
1.1 实验材料
柞蚕五龄幼虫由河南农业厅赠送;大肠杆菌和Top10 感受态细胞由本实验室所保存。
RNA 提取试剂盒Total RNA kit、质粒抽提试剂盒、DNA 纯化回收试剂盒购于OMEGA 公司,反转录试剂盒Transcriptor First strand cDNA synthesis kit 购于全式金公司,SMARTTM RACE cDNA Amplification Kit 购于BD Clontech 公司,Bam HI 和XhoI 核酸内切酶、T4 DNA Ligase 、pMD19 -T 载体试剂盒购于TaKaRa 公司,pET-28a 载体购于Promega 公司,其他试剂均为国产分析纯。
1.2 实验方法
1.2.1 引物设计与合成
根据核酸和氨基酸序列比对,用Primer premier 5.0 软件设计用于扩增柞蚕模式识别受体Lectin 部分片段的引物为:176F:5’- CTTCGCCCAGCAACTCCAATA - 3’ 和 176R:5’ - TTGACCCGTCGCCCAACTAAC - 3’,扩增片段长度为176bp。用于RACE - PCR 的引物RC3:5’- CTTCGCCCAGCAACTCCAATA-3’和RC5 :5’- TTGACCCGTCGCCCAACTAAC -3’,根据扩增获得的上述基因片段设计用于扩增ORF 框的引物为:fw:5’- GGCGGATCC- ATGTATAAGACATTCTTTTGTG - 3’和 rev:5’- CGCCTCGAGTCAATAGATATTTGGCGAA-3’(下划线部分分别为BamHI和XhoI 酶切位点)。以上引物均由上海英骏生物技术有限公司合成。
1.2.2 柞蚕总RNA 的提取和反转录
柞蚕五龄幼虫经灭活的大肠杆菌注射12 小时后按RNA 提取试剂盒Total RNA kit 提取总RNA,然后根据反转录试剂盒Transcriptor First strand cDNA synthesis kit 方法进行反转录,定量检测后保存于-20°C 冰箱备用。
1.2.3 PCR 扩增
以上步合成的cDNA 第一链为模板进行Lectin基因cDNA 的PCR 扩增。PCR 反应体系为:5 μL 10×Buffer、2 μL 引物176F、2 μL 引物176R、4 μL dNTPmix、1. 5 μL cDNA、0. 5 μL Taq 酶、35 μL ddH2O;PCR 反应条件为:预变性:94 ℃、4min;变性:94 ℃、0.5 min;退火:55 ℃、0. 5 min;延伸:72℃、0.5 min;从第2 步开始30 个循环;延伸:72 ℃、5 min。
1.2.4 RACE 技术获得全长cDNA 序列
参照SMARTTMRACE cDNA Amplification Kit(Clontech 公司)的使用说明书,将所提取高质量的RNA 反转录成RACE cDNA。利用BD AdvantageTM 2 PCR Enzyme System 和试剂盒中提供的10 ×Universal Primer A Mix (UPM)引物,以及所设计的特异引物分别以5’RACE cDNA 和3’RACE cDNA 为模板进行PCR 扩增。扩增程序为:预变性:94 ℃、5 min;5 个循环(变性:94 ℃、1 min;退火:65 ℃、2min);30 个循环(变性:94 ℃、1 min;退火:60 ℃、30 s;延伸:72 ℃、1 min)。扩增产物经1%琼脂糖凝胶电泳检测。
1.2.5 开放阅读框的序列扩增
通过序列分析比对和NCBI 上的ORF Finder 确定lectin 的开放阅读框序列,对ORF 框进行引物设计并引入酶切位点后进行PCR 扩增,扩增程序为:预变性:94 ℃、5 min;30 个循环(变性:94 ℃、30 s;退火:58 ℃、30 s;延伸:72 ℃、1 min);总延伸:72℃、5 min。扩增产物经1%琼脂糖凝胶电泳检测,将大小一致条带胶回收后连入pMD19 -T 载体中,挑选阳性克隆菌落进行扩大培养和质粒抽提,并送至上海英骏生物技术有限公司测序。
1.2.6 原核表达载体的构建
从前面构建的测序鉴定正确的Lectin 的T 载体质粒上用BamHI 和XhoI 双酶切,切下目的片段,原核表达载体pET28a 经限制性内切酶BamHI 和XhoI双酶切,酶切反应体系共50 μL:质粒10 μL,10 ×K Buffer 5 μL,BamHI 和XhoI 酶各1 μL,ddH2O 33 μL。37 ℃酶切4h 后产物分别胶回收,然后经1%琼脂糖凝胶电泳检测并测浓度,再用T4DNA 连接酶将Lectin 片段连入原核表达载体pET28a 的BamHI和XhoI 酶切位点间。连接反应体系为10 μL:酶切Lectin 5 μL,酶切pET28a 1 μL,10 ×T4 Buffer 1 μL,T4 DNA Ligase 1 μL,ddH2O 2 μL,4 ℃连接过夜。将连接产物转化入E. coli DH5α 中.挑取阳性单菌落培养,并用质粒提取试剂盒提取质粒,用BamHI和XhoI 双酶切后,经1%琼脂糖凝胶电泳来鉴定是否有外源片段插入,并送至上海英骏生物技术有限公司测序。
2 结果
2.1 凝集素基因部分片段的获得
以柞蚕幼虫cDNA 为模板进行RT-PCR。PCR扩增产物经1%琼脂糖凝胶电泳检测后的结果如图1,在分子量约176 bp 左右大小的位置存在一条清晰的条带,该条带的大小与实验预期的大小相近。
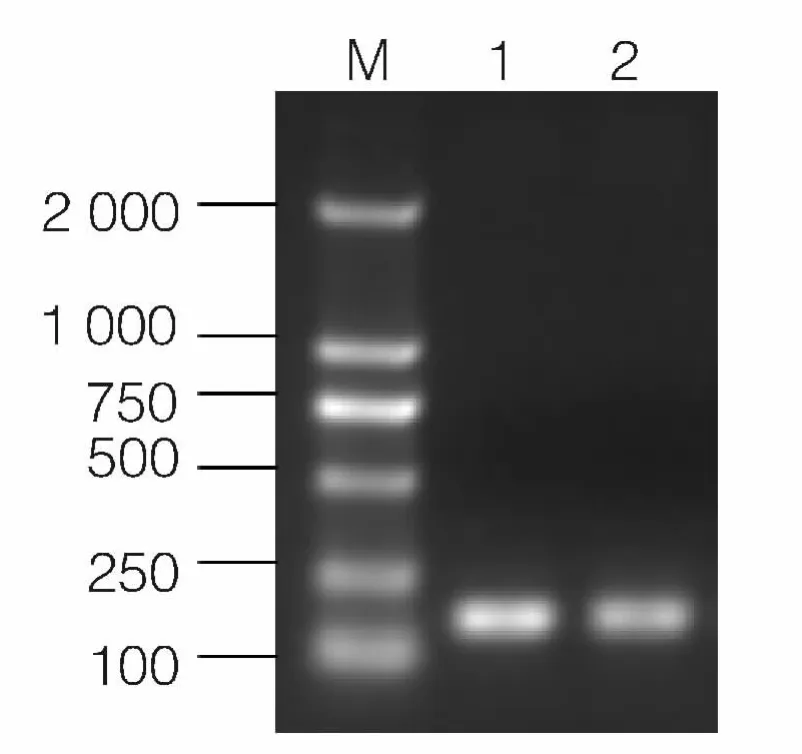
图1 PCR 扩增获得的Lectin 部分片段泳道1、2 为目的片段;M 为分子量标准
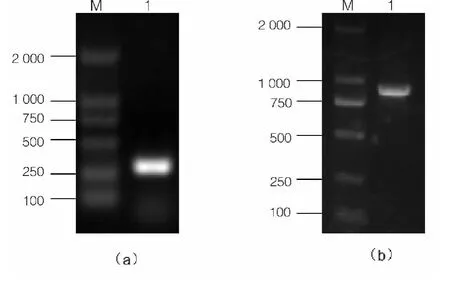
图2 3’-和5’-RACE PCR 产物的电泳分析
2.2 RACE 技术获得基因全长cDNA
根据已获得的Lectin 基因的部分cDNA 序列,使用Primer premier 5.0 设计用于扩增全长cDNA 的RACE 引物。以合成的相对应的RACE cDNA 为模板,通用引物进行RACE PCR 反应。结果表明,3'-RACE 和5'-RACE 分别获得了分子量大小为825 bp 和368 bp 的片段,分别见图2(a)和图2(b)。通过DNAMAN 软件进行序列拼接,获得lectin 基因全序列(图3)。

图3 柞蚕凝集素全长cDNA 序列
起始密码子用方框标出,终止密码子用双下划线标出。
2.3 开放阅读框的确定和扩增
通过序列分析比对和NCBI 上的ORF Finder 确定lectin 的开放阅读框,对ORF 框引入酶切位点进行PCR 扩增,方法如前所述,片段与预期大小一致,如图4。
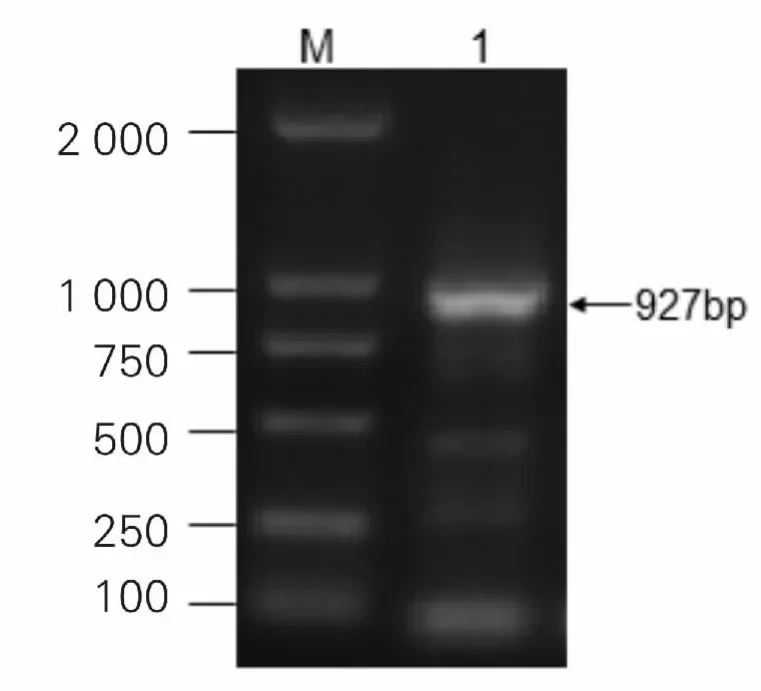
图4 PCR 扩增获得的Lectin 目的片段1 为目的片段;M 为分子量标准
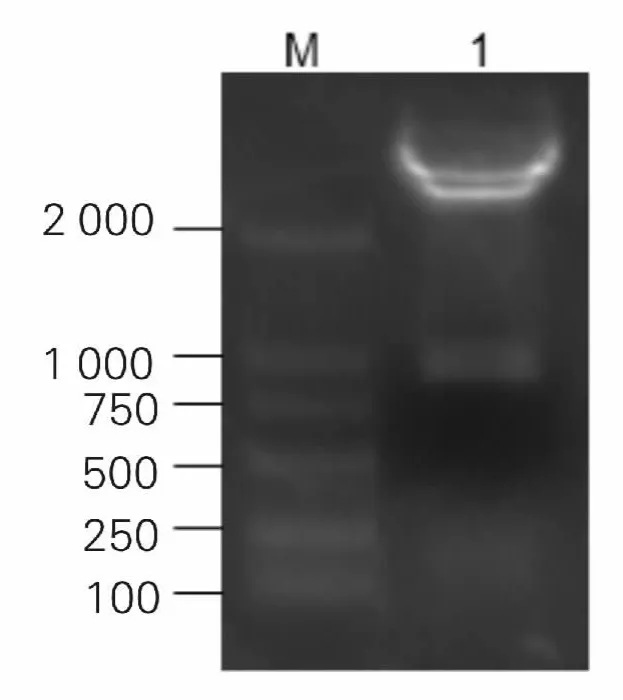
图5 重组质粒的酶切鉴定
2.4 重组质粒PET-28a-Lectin 的构建和鉴定
Lectin 基因经BamHI 和XhoI 双酶切克隆入经同样酶切的原核表达载体pET28a,挑选阳性重组质粒进行酶切及电泳,方法如前,可见大小约为927 bp的目的条带,经测序表明重组质粒构建成功,见图5。将其命名为PET-28a-Lectin。
3 讨论
先天免疫系统是脊椎和无脊椎的第一道免疫防御阵线,其发生过程借助于模式识别受体蛋白来识别与结合病原微生物表面的相关分子模式[11~14]。因而,通过模式识别受体蛋白,昆虫的先天免疫反应能够识别和结合不同的病原微生物,从而达到抗菌和抑菌的作用。动物C-型凝集素通过结合包括细菌、真菌和病毒等病原微生物表面的碳水化合物来促进吞噬、黑化和酶原反应等免疫反应[15~20]。关于柞蚕的C 型-凝集素及其先天免疫反应系统的研究鲜见报道,研究柞蚕的C-型凝集素是必要的,本研究运用RT -PCR 和RACE 技术从柞蚕中获得了C 型-凝集素基因的全长序列并构建了一个能够表达Lectin 蛋白的重组表达载体,以便为今后研究Lectin 蛋白的功能及参与柞蚕的先天免疫反应的机理奠定基础。
[1]Janeway Jr.,C.A. The immune system evolved to discriminate infectious nonself from noninfectious self [J]. Immunol.Today 1992,13:11 ~16.
[2]Janeway Jr.,C.A.,Medzhitov,R. Innate immune recognition[J]. Annu. Rev.Immunol. 2002,20:197 -216.
[3]李志强,陈国生,王茂先,等. 昆虫体液免疫的分子生物学[J].生命的化学,2003,23 (5):348 ~351.
[4]Chen CL,Rowley AF,Newton RP,et al. Identification,purification and properties of a beta -1,3 - glucan - specific lectin from the serum of the cockroach,Blaberus discoidalis which is implicated in immune defense reactions [J]. Comp Biochem Physiol B,Biochem Mol Biol,1999,12(2):309 ~319.
[5]Stratton L,Wu S,Richards RC,et al. Oligomerisation and carbohydrate binding in an Atlantic salmon serum C-type lectin consistent with non-self recognition[J]. Fish Shellfish Immunol,2004,17(4):315 ~323.
[6]Drickamer K. Two distinct classes of carbohydraterecognition domains in animal lectins [J]. J Biol Chem,1988,263(20):9557 ~9560.
[7]Jingqun Ao,Erjun Ling,Xiao-Qiang Yu. Drosophila Ctype lectins enhance cellular encapsulation[J]. Molecular Immunology,2007,44:2541 ~2548.
[8]Yu,X. Q.,Tracy,M. E.,Ling,E.,Scholz,F. R.,Trenczek,T. A novel C - type immulectin -3 from Manduca sexta is translocated from hemolymph into the cytoplasm of hemocytes[J]. Insect Biochem. Mol. Biol. 2005,35:285 ~295.
[9]Ling,E.,Yu,X.Q. Cellular encapsulation and melanization are enhanced by immulectins,pattern recognition receptors from the tobacco hornworm Manduca sexta [J]. Dev. Comp.Immunol. 2006,30:289 ~299.
[10]Jia -Lin Wang ,Xu -Sheng Liu,Qi Zhang,et al,. Expression profiles of six novel C-type lectins in response to bacterial and 20E injection in the cotton bollworm (Helicoverpa armigera)[J]. Developmental and Comparative Immunology.2012,37:221 ~232.
[11]Choe K M,Lee H,Anderson K V. Drosophila peptidoglycan recognition protein LC (PGRP - LC )acts as a signal -transducing innate immune receptor[J]. Proc Nail Acad Sci U S A,2005,102(4):1122 ~126
[12]Hoffmann J A. The immune response of Drosophila[J].Nature,2003,426 (6962):33
[13]许平震,张美蓉,程廷才,等. 家蚕模式识别受体βGRP4 的序列分析及诱导表达[J].蚕业科学,2011,37(4):637 ~641
[14]Dziarski R. Peptidoglycan recognition proteins (PGRPs)[J].Mol Immunol,2004,40 (12):877.
[15]Weis,W.I.,Taylor,M.E.,Drickamer,K.,1998. The C - type lectin superfamily in the immune system. Immunol.Rev. 163,19 ~34.
[16]Holmskov,U.,Thiel,S.,Jensenius,J.C.,2003. Collections and ficolins:humoral lectins of the innate immune defense[J]. Annu. Rev. Immunol. 21,547 ~578.
[17]Jack,D.L.,Turner,M.W.,2003. Anti -microbial activities of mannose-binding lectin[J]. Biochem. Soc. Trans.31,753 ~757
[18]Yu,X. -Q.,Kanost,M. R.,2004. Immulectin -2,a pattern recognition receptor that stimulates hemocyte encapsulation and melanization in the tobacco hornworm,Manduca sexta[J]. Dev. Comp. Immunol. 28,891 ~900.
[19]Ling,E.,Yu,X. - Q.,2006. Hemocytes from the tobacco hornworm Manduca sexta have distinct functions in phagocytosis of foreign particles and self dead cells[J]. Dev. Comp.Immunol. 30,301 ~309.
[20]Yu,X. -Q.,Gan,H.,Kanost,M.R.,1999. Immulectin,an inducible C-type lectin from an insect,Manduca sexta,stimulates activation of plasma prophenolOxidase [J]. Insect Biochem. Mol. Biol. 29,585 ~597.
