青藏高原20种灌木生长时期根系特征及抗旱性初探
2014-01-04李吉跃王军辉马建伟
邱 权 ,潘 昕 ,李吉跃 ,王军辉 ,董 蕾 ,马建伟 ,杜 坤
(1.华南农业大学 林学院,广东 广州 510642;2.中国林业科学研究院 林业研究所 国家林业局林木培育重点实验室, 北京 100091;3甘肃省小陇山林业科学研究所, 甘肃 天水 741022)
根系是植物吸收水分和养分的重要器官,与土壤水分有密切关系,在植物适应干旱胁迫生境中发挥着重要作用[1-5]。国内外很多专家学者认为通过研究植物根系特征可以一定程度上指示植物抗旱性能差异[6-10],其中以研究根系生物量、根系活力、根系形态特征、生理指标为主。在最近报道的植物抗旱性能研究中,根总长度、表面积、体积和根尖数等根系特征运用较多[5,10],并且已有研究表明抗旱性与根系生物量和微细根特征关系密切[5,11]。微细根是主要的吸收和代谢根,表皮薄,幼嫩,膜透性强,具有吸收表面积大、生理活性强等特点,是树木水分和养分吸收的主要器官[12]。微细根生物量(直径≤2 mm)仅占根系总生物量的3%~30%[12],但其数量占全根数量比例最大[13-14],研究认为微细根长度、表面积、根尖数量等指标对土壤环境变化更敏感[15-19],更能直接体现灌木对干旱胁迫的响应,而关于此方面内容的研究报道不多。青藏高原具有典型的高原干旱、寒冷气候特征,生态环境处于持续退化状态之中,而通过栽植抗旱的固沙固土植物,是防治环境进一步恶化的有效手段,筛选出抗旱性强树种便是这项工作中的重点之一。目前的研究中,已有很多通过研究根系特征评价植物抗旱性能差异的相关报道[5-11,20-24],其中陈明涛等[5]便通过根系特征研究黄土高原4种造林树种抗旱性能差异,也有通过田间试验测定植物根系特征来进行植物抗旱性筛选的相关研究[9],但针对青藏高原地区此方面内容研究较少。
本研究以青藏高原地区各地自然分布的柠条Caragana korshinskii等20种灌木树种为试材在苗圃培育,通过测定3年生苗生长时期全根系特征(总长度、表面积、体积、根尖数、分叉数)以及对应微细根特征,从根系特征角度综合分析种间差异,并通过聚类分析初步评价其抗旱性能的差异,为青藏高原干旱地区灌木树种抗逆性(耐旱)评价指标的建立及各种灌木树种的合理立地配置提供必要的科学依据。
1 材料与方法
1.1 研究地区概况
试验设在甘肃省小陇山林科所苗圃,该地位于 105°48′E,34°29′N,海拔 1 160 m。属温带半湿润季风气候,年降水量600~800 mm,年蒸发量1 290 mm, 年平均气温 11.0 ℃,极端最高气温39.0℃,极端最低气温-19.2℃。4~9月晴天日平均光照时数11~14 h,夏季最高光照强度 2 500 μmol·m-2s-1,无霜期约 180 d。
1.2 研究材料
2011年5月将分布在青藏高原地区各地的柠条等20种灌木树种2年生幼苗移栽在甘肃小陇山林业科学研究所苗圃内培养。采用完全随机区组排列,同种灌木种植在同一区组内,每种50株。
1.3 研究方法
根系分析方法参考文献[25-26],根系划分为微细根(直径≤2 mm)、粗细根(2~5 mm)、粗根(>5 mm)。于2012年5月(生长初期)和7月(生长旺季)每种灌木分别随机选取5株苗木的完整根系(外业获取的根系要放入冰盒中保鲜,防止根系脱水、体积变小)置于密封袋,带回实验室后用根系扫描系统分析根系形态特征,包括总根长(TRL)、表面积(TRSA)、体积(TRV)、根尖数(TRTN)和分叉数(TRBN),并对根系进行分级,重点研究微细根(d≤2 mm)根长(FRTL)、表面积(SFRSA)、体积(SFRV)及根尖数(SFRTN)。根系扫描仪为中晶ScanMaker i800 Plus,分析软件为万深LA-S植物根系分析系统(杭州万深检测科技有限公司)。处理前,将根系样品从冰盒中取出,用清水小心冲洗掉根系表面的泥土。如遇到很细的根,则在根系下面放置100目筛以防止脱落的根系被水冲走。

表1 灌木基本信息Table 1 Basic status of shrubs
1.4 数据处理与分析
采 用 Excel 2007 作 图,SPSS(19.0) 进 行ANOVA方差分析、相关性分析和主成分分析。
2 结果与分析
2.1 灌木生长时期全根系特征
2.1.1 灌木生长初期(5月)全根系特征
本研究20种灌木树种根系形态均为直根系,根系分为主根和侧根。大部分灌木为浅根型植物,垂直根系不发达,其根系主要分布在土层深度0~60 cm处,但水平方向侧根发达,不断向四周延展,仅柠条、胡颓子、锦鸡儿、树锦鸡儿等深根型灌木树种主根扎根较深。由表2可以看出,5月份灌木间根系各全根系特征值均存在显著性差异(P<0.05),不同灌木不同根系特征值有所不同。根系总长度方面,金露梅、三颗针、胡颓子、柠条、绣线菊较大,分别为(4 571.69±127.89)、(2 250.71±323.18)、(2 197.67±74.72)、(1 387.77±139.02)、(1 311.15 ±76.01)cm,而驼绒藜、西北沼委陵菜、西北小蘗、尼泊尔锦鸡儿、江孜沙棘则相对较小,介于250.15~417.79 cm之间。表面积和体积方面,基本呈现出一致的规律,均是柠条、胡颓子、金露梅、三颗针、蔷薇相对较大,而江孜沙棘、柳叶沙棘、尼泊尔锦鸡儿、西北小蘗、西北沼委陵菜在20种灌木中数值普遍偏低。根尖数和分叉数方面,数值较高的均是柠条、胡颓子、金露梅、三颗针、绣线菊,根尖数量较少的为江孜沙棘、柳叶沙棘、尼泊尔锦鸡儿、西北小蘗 、西藏沙棘,介于639~982之间,而分叉数量较少的是江孜沙棘、尼泊尔锦鸡儿、驼绒藜、西藏沙棘、西北沼委陵菜,分别仅占分叉数最多的金露梅(1 6043±635)的3.07%、3.65%、3.40、4.04%、5.49%。
2.1.2 灌木生长旺期(7月)全根系特征
由表3可以看出,7月份灌木根系各全根系特征值均存在显著性差异(P<0.05),7月份灌木各全根系特征值所呈现出的大小顺序规律与5月份有所不同。就根系总长度而言,金露梅、蔷薇、树锦鸡儿、三颗针、绣线菊相对较高,江孜沙棘、柳叶沙棘、尼泊尔锦鸡儿、驼绒藜、西北沼委陵菜则相对较低,仅介于282.74~518.83 cm之间。表面积和体积方面,数值较高的分别为胡颓子、金露梅、蔷薇、三颗针、绣线菊,而数值相对较小的均是江孜沙棘、柳叶沙棘、尼泊尔锦鸡儿、西北小蘗、西北沼委陵菜,其数值分别介于80.11~142.85 cm2和3.11~8.93 cm3之间。根尖数和分叉数方面,金露梅、蔷薇、树锦鸡儿、三颗针、绣线菊均高于其它灌木树种,而江孜沙棘、柳叶沙棘、尼泊尔锦鸡儿、驼绒藜、西北沼委陵菜则相对较低,其根尖数和分叉数分别介于718~1 532和560~1 231之间。以图1可以看出,
与5月份相比,7月份20种灌木全根长、表面积、体积、根尖数、分叉数的平均值分别增加了84.91%、119.87%、175.14%、85.08%、109.28%,说明当灌木进入生长旺期时,全根体积比其它4个全根系特征值增加更多。

表2 5月份灌木全根系特征值†Table 2 Characteristics values of total roots of 20 shrubs in May
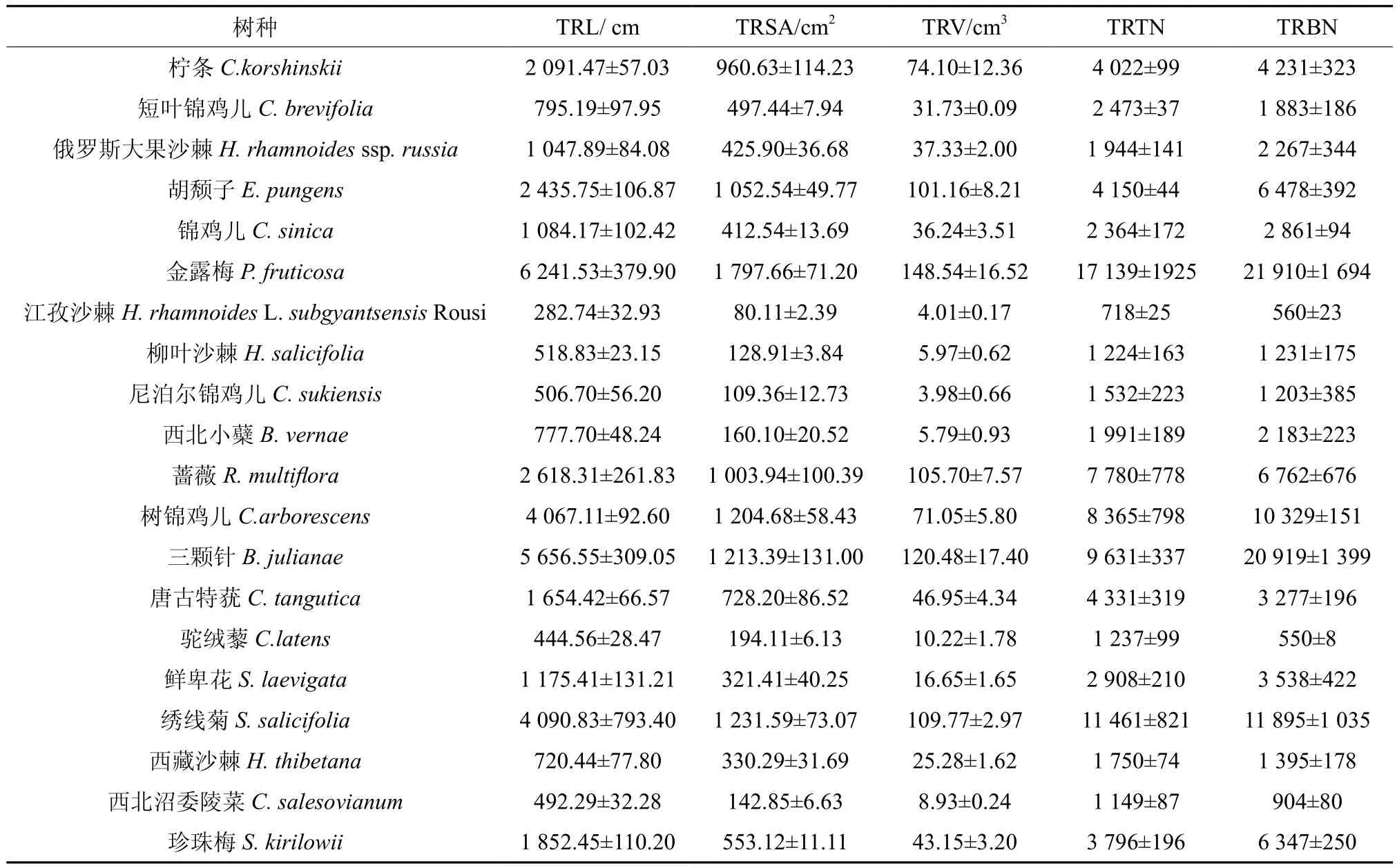
表3 7月份灌木全根系特征值Table 3 Characteristics values of total roots of 20 shrubs in July
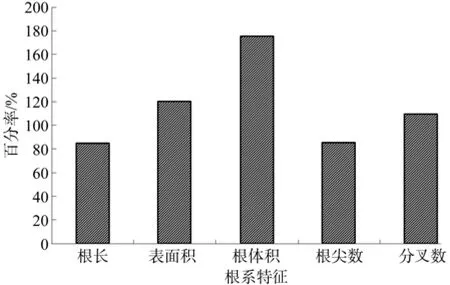
图1 7月全根系特征值平均值比5月增加百分率Fig.1 Percentage of increase in average characteristic values of total roots in July over that in May
2.2 灌木不同生长时期微细根特征
2.2.1 灌木生长初期(5月)微细根系特征
从表4可以看出,5月份灌木根系微细根特征值均存在显著性差异(P<0.05),灌木各微细根特征值大小顺序不一致。微细根长度方面,柠条、胡颓子、金露梅、三颗针、绣线菊高于其它灌木,分别达到了(1 208.31±64.60)、(1 698.17±28.97)、(4 395.36±93.34)、(2 230.44±221.26)、(1 234.68±115.62) cm,而江孜沙棘、尼泊尔锦鸡儿、西北小蘗、驼绒藜、西北沼委陵菜相对偏低,仅介于225.15~349.44 cm之间。微细根表面积、体积和根尖数方面,数值较高均是柠条、胡颓子、金露梅、三颗针、珍珠梅,数值较低灌木树种总体上相同,其中微细根表面积和体积较小的为江孜沙棘、尼泊尔锦鸡儿、西北小蘗、鲜卑花、西北沼委陵菜,而根尖数方面数值较小的为江孜沙棘、尼泊尔锦鸡儿、西北小蘗、西藏沙棘、西北沼委陵菜。
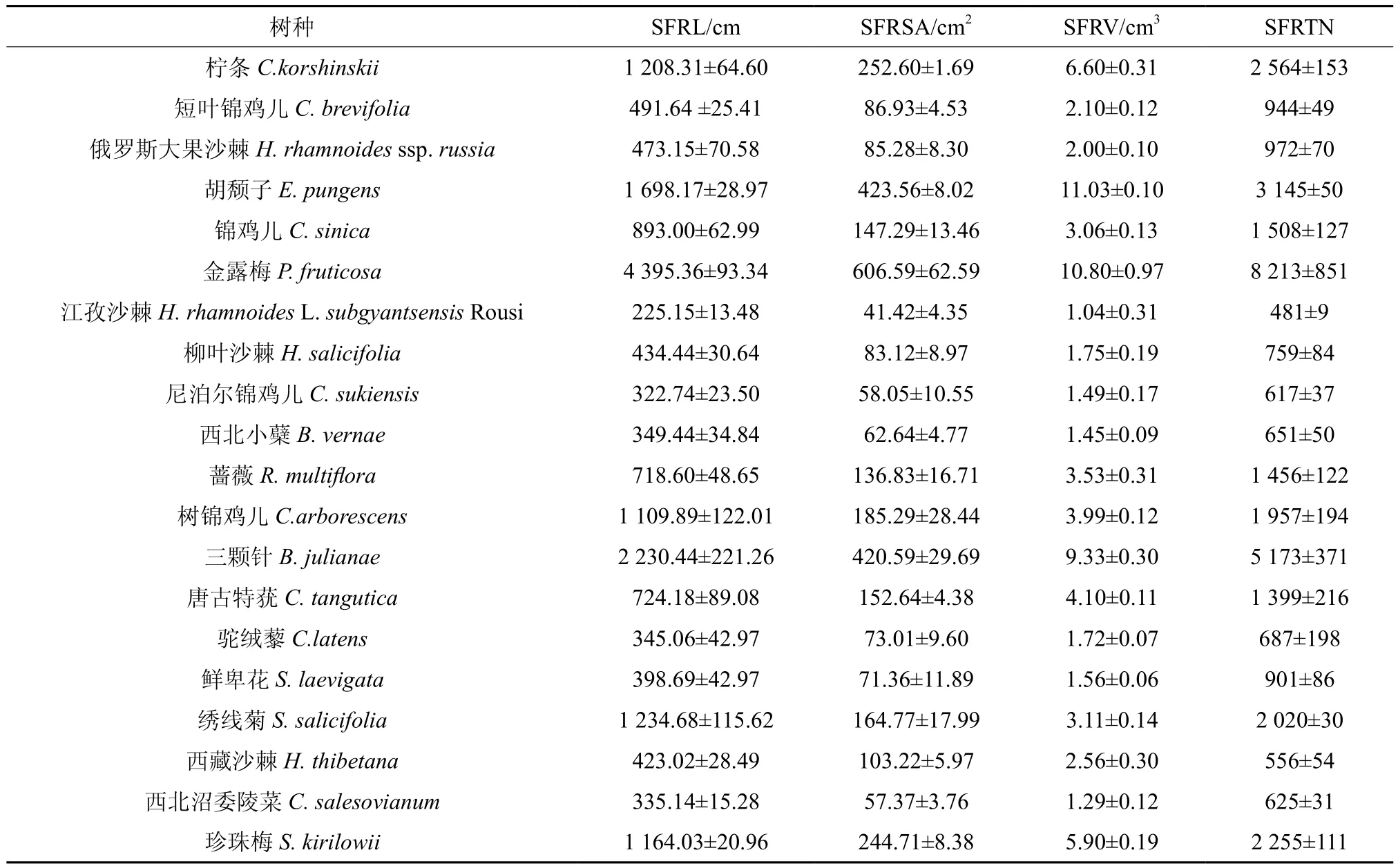
表4 5月份灌木微细根系特征值†Table 4 Characteristics values of micro-fine roots of 20 shrubs in May
2.2.2 灌木生长旺期(7月)微细根系特征
7月份灌木根系微细根特征值均存在显著性差异(P<0.05),灌木微细根特征值大小顺序规律总体上与5月份一致,但也存在一定差异。由表5可知,微细根根长数值较大的为金露梅、蔷薇、树锦鸡儿、三颗针、绣线菊,分别为(4 972.53±152.21)、(2 232.63±223.26)、(3 651.55±154.64)、(5 371.61±402.26)、(3 114.82±335.05) cm。微细根表面积、体积方面,数值较大均是胡颓子、金露梅、树锦鸡儿、三颗针、绣线菊,其微细根表面积和体积分别介于451.15~972.90 cm2和11.44~23.86 cm3之间。根尖数方面则是胡颓子、金露梅、蔷薇、树锦鸡儿、三颗针高于其它灌木树种,尤其是金露梅,甚至高达(12 516±795)。另一方面,20种灌木4个微细根特征值较小均是江孜沙棘、柳叶沙棘、尼泊尔锦鸡儿、驼绒藜、西北沼委陵菜,表明此5种灌木在7月份微细根生长状况总体表现不佳,明显滞后于其它灌木树种。与5月份相比,20种灌木7月份微细根根长、表面积、体积和根尖数总体呈现不同幅度增加(见图2),此4项指标平均增幅分别为54.28%、61.55%、72.67%和74.56%,增幅最大的为微细根根尖数。

表5 灌木7月微细根系特征值Table 5 Characteristics values of micro-fine roots of 20 shrubs in July
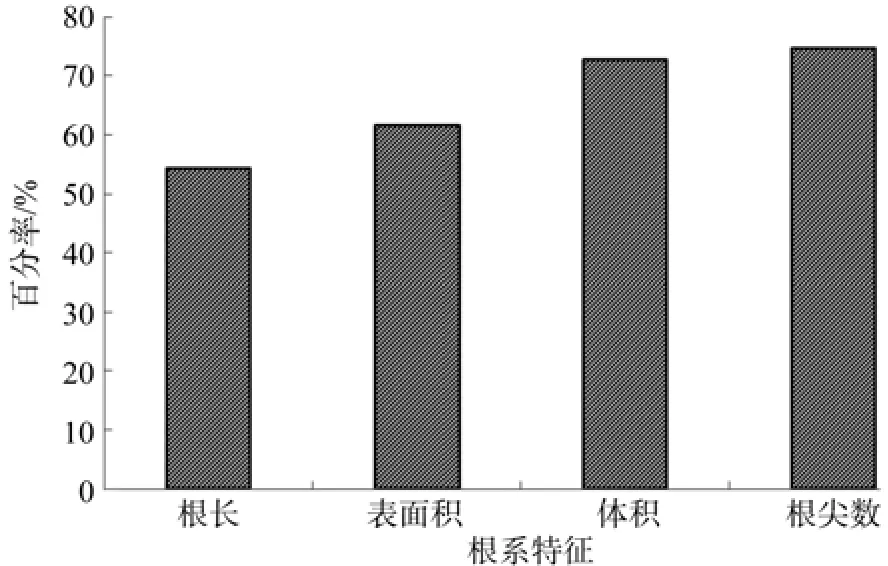
图2 7月微细根系特征值平均值比5月增加百分率Fig.2 Percentage of increase in average characteristic values of total roots in July over that in May
2.3 根系特征值相关性分析
根系特征值相关系分析结果显示,各根系特征值之间存在不同程度的相关性(见表6)。表6可以看出,各指标间总体上呈显著正相关关系,并且大部分指标间相关性程度达到了极显著水平,说明各指标间存在着紧密联系。其中,4个微细根系特征值之间均呈极显著正相关,5个全根系特征值之间相关性总体呈极显著正相关,仅全根体积与全根分叉数呈显著相关,而值得注意的是微细根系特征值与全根系特征值之间相关性出现了一些指标间相关性不显著的现象,即5月份全根体积与各微细根特征值相关性不显著,相关系数仅介于0.366~0.424之间。比较5月份和7月份各指标间相互关系对应的相关系数,可以明显看出,总体上呈现7月份大于5月份的规律,而且7月份相关系数均超过了0.8,由此可见7月份各指标间联系更为密切。
2.4 主成分分析
为了更好地对灌木根系特征指标进行综合评价,选择5月份和7月份总根长、表面积、体积、根尖数、分叉数和微细根根长、表面积、体积、分叉数共18个相关指标进行主成分分析。表7中可以看出,第一主成分贡献率最大,达到了79.490%,涵盖了主要的根系特征信息,第一、二主成分累计贡献率为90.247%,可以很好地反映出各根系特征值的相对重要性以及相互关系。从各主成分上的因子载荷分析结果(见表7)来看,各指标第一主成分影响均较大(>0.6),对于揭示根系特征均有重要意义,而第二主成分中,各指标载荷均较低,仅有5月份微细根长和微细根表面积载荷略大于0.5。就第一主成分各指标因子载荷情况来看,7月份各项指标载荷均比5月份相对应指标高,并且7月份根系特征测定指标中,微细根指标载荷相对高于对应全根系指标,说明在进行根系特征值测定时,选择灌木生长旺期(7月)的微细根相关根系特征值,即能很好地指示灌木根系特征。

表6 根系特征值之间的相关系数†Table 6 Correlation coefficients of characteristic values of roots
2.5 根系特征聚类分析
根据主成分分析结果,分别计算第一、二主成分因子得分,以第一主成分因子得分为横轴,第二主成分因子为纵轴,绘制散点图(见图3)。为系统研究各灌木树种根系特征差异,对20种灌木树种按照第一、二主成分得分值进行聚类分析。研究结果表明,从根系特征角度综合考虑,20种灌木大致可以分为5种类型:金露梅为第1类;三颗针、树锦鸡儿为第2类;柠条、胡颓子、蔷薇和绣线菊为第3类;唐古特莸和珍珠梅为第4类;其余11种灌木为第5类。

表7 主成分因子载荷矩阵及主成分特征根Table 7 Factor loading matrix of main composition and principal component eigenvalues
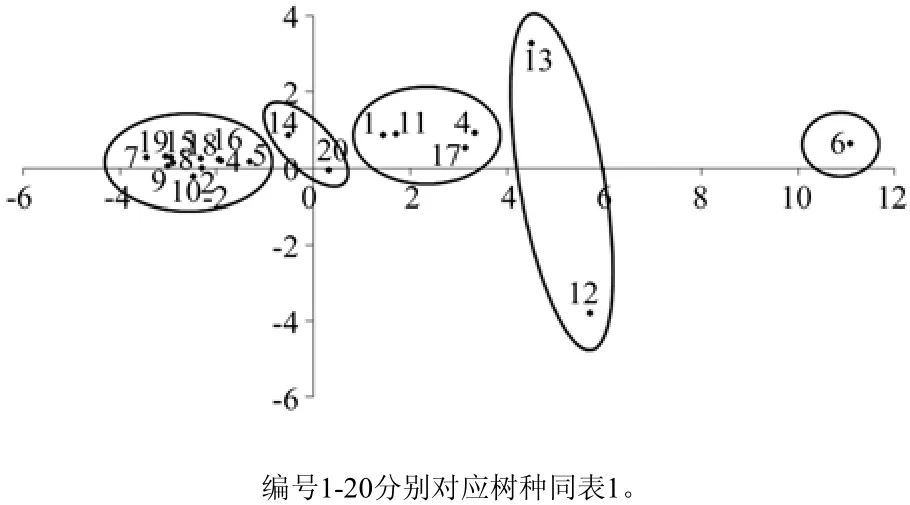
图3 聚类结果Fig.3 Cluster result
3 结论与讨论
3.1 灌木根系抗旱机制
根表面积反映根系的吸收能力,可作为衡量根系质量的一个重要标准[25]。根长、根体积、根尖数、分叉数也是衡量根系质量的重要指标[26-27],根系越长说明垂直扎根深或者侧根水平伸展范围广,植物根系可以吸收土层深度较深或水平方向上距离较远的土壤水分,根体积越大表明可吸收利用水分范围越大,根尖数越多说明须根越多,吸水能力越强,而分叉数越多说明侧根及分枝越多。从各根系特征值之间相关性分析结果来看,各指标间总体上呈显著正相关关系,说明各指标间联系密切,可见灌木在长期干旱逆境生长过程中,根系形态特征已经朝根系特征值最大化方向趋于统一,通过全面提高根系抗旱能力以适应干旱胁迫逆境,这可能与灌木树种在青藏高原地区长期逆境生长过程中产生的环境适应性有关,因为已有大量研究表明植物根系生长动态与土壤水分状况具有密切联系[28-29],而Tjeerd 等[30]的研究也证实了土壤水分通过影响土壤的通透性和根系呼吸进而影响到根系生长的相关结论。本研究中所用灌木树种均主要分布在水分较亏缺的青藏高原各地区,根系抗旱特征存在一定共性,而这些灌木均依赖其共同根系抗旱机制,才能在干旱逆境中生存。
3.2 全根系特征值与微细根特征值的比较
研究中通常把根系划分为微细根(直径≤2 mm)、粗细根(直径2~5 mm)和粗根(直径>5 mm),而微细根主要功能是进行水分、矿物养分的吸收和运输。已有研究表明,细根和粗根对土壤的反应敏感程度不一致,其中细根较为活跃[31]。本研究中灌木生长时期各微细根特征值与全根系特征值之间总体呈显著正相关,可见微细根和全根系之间存在一定的联系。研究结果显示灌木生长时期微细根系和全根系呈现出了相同的生长变化规律,也表现出一定差异性。全根系特征值指标之间和微细根特征值指标之间均呈显著正相关,此外,在相同生长时期,20种灌木全根系特征值和微细根特征值均存在显著性差异,可见研究全根系特征值和微细根特征值均是研究灌木树种间根系特征差异的有效指标。当由5月份进入7月份后,微细根和全根系特征值均有所增加,但各特征值增幅有所不同,其中全根系和微细根特征值中增幅最大的分别是全根体积和微细根根尖数。另一方面,主成分分析结果表明微细根特征值指标载荷会普遍高于全根系特征值,可见研究灌木树种微细根特征值更能说明灌木根系特征差异。
3.3 灌木根系特征值生长时期动态变化
已有研究表明,根系特征具有明显的季节变化[29],主要受土壤水分、温度、养分和树木自身生物学特性等因素综合影响[32-35]。本研究中灌木从5月份进入7月份后,其根系特征值指标呈不同幅度增加趋势,可见当灌木处在生长旺盛时期时,更好的水热条件以及植物本身更活跃的生理活性一定程度促进了根系的生长发育。从研究结果来看,灌木7月份各根系特征值之间相关性程度总体高于5月份,而且7月份根系特征值指标标载荷相对高于5月份,由此说明研究生长旺盛时期(7月)灌木各项根系特征指标效果更好。
3.4 灌木抗旱能力评价以及与原产地的联系
植物抗旱性是形态解剖、生理生化等许多因素协调作用的结果,但根系居主导地位。从根系特征角度来讲,根长、表面积、体积根尖数和分叉数这些根系特征值均与抗旱性相关[9]。一般认为,植物抗旱能力与各根系特征值呈正相关关系,即根系特征值越大抗旱性更强[25-27],因此,可以根据根系特征聚类结果对灌木树种抗旱性能进行初步评定。通过田间试验观察灌木生长时期根系特征,综合分析20种灌木全根系和微细根系特征值,从根系特征角度聚类分析结果初步评定其抗旱能力,大致可以分为5种类型(抗旱能力依次减弱):金露梅为第1类,抗旱性最强;三颗针、树锦鸡儿为第2类;柠条、胡颓子、蔷薇和绣线菊为第3类;唐古特莸和珍珠梅为第4类;其余11种灌木为第5类。从初步研究结果来看,金露梅、三颗针、树锦鸡儿、柠条、胡颓子、蔷薇、绣线菊、唐古特莸和珍珠梅根系发育较好,其发达的根系组织能更好地适应干旱逆境,可以在青藏高原地区耐旱造林树种选择时给予重点考虑,而锦鸡儿等其余11种灌木树种根系抗旱能力则相对较弱,应配置于相应立地。本研究中20种所选灌木树种主要以叶面积小、旱生结构明显、能在长期干旱环境下生存以及已经报道的抗旱性较强的灌木为主,分别来自青海、西藏和甘肃地区,三个地区均属于干旱地区,但气候因子有明显差异。甘肃省属温带季风气候,具有明显的向大陆性气候过渡的特征,全省干旱缺雨。青海和西藏属青藏高原,其中青海气候以高寒干旱为总特征,典型的大陆性高原气候。西藏气候特征复杂,主要以高原气候和温带大陆性气候为主,大部分地区全年降雨较少。金露梅、三颗针、树锦鸡儿为根系抗旱性较强的树种,其原产地均在青海地区,而抗旱性较差的灌木树种主要来自西藏地区,可见选择青海地区灌木在干旱地区进行造林效果较好,可能与青海地区特殊的气候环境有关,其较为干旱的气候条件及水分相对亏缺的土壤条件使灌木产生了较强的生态适应性。
本研究采用田间试验的方法,通过搜集青藏高原地区灌木资源在大田野外种植,进行根系特征观察,所以灌木树种抗旱能力评价只是一个初步判定结果,并且所得结论仅仅基于灌木在生长季节的根系特征,存在一定局限性。另一方面,本研究选择在相对干旱的甘肃地区进行,可能土壤水分环境比较接近青藏高原地区,但是其它土壤理化特性存在一定差异,因此本文中相关研究结论可能无法真实反映各灌木树种野外原产区的根系生长表现,必须及时开展各灌木树种在各自原产地的不同生长季的根系特征研究,以期科学认识其在原产区根系生长时空动态变化规律。
[1] Christmann A, Weiler E W, Steudle E, et al. A hydraulic signal in root-to-shoot signaling of water shortage[J].The Plant Journal,2007,52(1):167-174.
[2] Dodd I C. Root-to-shoot signaling: assessing the roles of “up”in the up and down world of long-distance signaling in planta[J].Plant and Soil, 2005, 274(1-2):251-270.
[3] Wright R A, Wein R A, Dancik B P. Population differentiation in seedling root size between adjacent stands of jack pine[J]. Forest Science, 1992,38(4):777-785.
[4] Toorchi M, Shashidhar H E, Hittalmani S, et al. Rice root morphology under contrasting moisture regimes and contribution of molecular marker heterozygosity[J].Euphytica,2002,126(2):251-257.
[5] 陈明涛,赵 忠.干旱对4种苗木根系特征及各部分物质分配的影响[J].北京林业大学学报,2011,33(1):16-22.
[6] 张军红,韩海燕,雷雅凯,等. 不同固定程度沙地油蒿根系与土壤水分特征研究[J]. 西南林业大学学报,2012,32(6): 1-5.
[7] 蒲伟凤,李桂兰,张 敏,等. 干旱胁迫对野生和栽培大豆根系特征及生理指标的影响[J]. 大豆科学,2010,29(4): 615-622.
[8] 李鲁华,李世清,翟军海,等.小麦根系与土壤水分胁迫关系的研究进展[J]. 西北植物学报,2001,21(1): 1-7.
[9] 贾强生,卫 铃,杨海峰. 大豆幼苗期根系特性和抗旱性关系初探[J]. 陕西农业科学, 2006(2): 12-13.
[10] 李德顺,刘 芳,马永光. 玉米根系与抗旱性关系研究[J]. 杂粮作物,2010,30(3): 195-197.
[11] 单长卷,梁宗锁. 土壤干旱对冬小麦幼苗根系生长及生理特性的影响[J]. 中国生态农业学报,2007,15(5):38-41.
[12] Usman S, Singh S P, Rawat Y S, et al. Fine root decomposition and nitrogen minerralisation patterns in Quercus leucotrichophora and Pinus roxburghii forest in central Himalaya[J]. Forest Ecology and Management, 2000,131(1-3):191-199.
[13] 李林锋,刘新田. 干旱胁迫对桉树幼苗的生长和某些生理生态特性的影响[J]. 西北林学院学报,2004,19(1):14-17.
[14] Cheng S, Widden P, Missier C. Light and tree size influence belowground development in yellow birch and sugar maple[J].Plant and Soil, 2005,270(1):321-330.
[15] Cheng S. Lorentzian model of roots for understory yellow birch and sugar maple saplings[J]. Journal of Theoretical Biology,2007, 246(2):309-322.
[16] Bakker M R,Kerisit R,Verbist K, et al. Effects of liming on rhizoshere chemistry and growth of fi ne roots and of shoots of sessile oak (Quercus petraea) [J]. Plant and Soil, 1999,217(1/2):243-255.
[17] Block R M A, van Rsse K C J, Knight J D. A review of fi ne root dynamics in Populus plantations[J]. Agroforestry Systems, 2006,67(1): 73-84.
[18] Trubat R, Cortina J, Vilagrosa A. Plant morphology and root hydraulics are altered by nutrient def i ciency in Pistacia lentiscus L[J]. Trees-Structure and Function, 2006, 20(3): 334-339.
[19] Moore J D, Camire C, Ouimet R. Effects of liming on the nutrition, vigor, and growth of sugar maple at the Lake Clair Watershed, Quebec, Canada[J]. Canadian Journal of Forest Research, 2000,30(5): 725-732.
[21] 王家顺,李志友.干旱胁迫对茶树根系形态特征的影响[J].河南农业科学,2011,40(9):55-57.
[22] 张承林,付子轼. 水分胁迫对荔枝幼树根系与梢生长的影响[J].果树学报,2005,22(4):339-342.
[23] 孔艳菊,孙明高,胡学俭,等. 干旱胁迫对黄栌幼苗几个生理指标的影响[J].中南林学院学报,2006,26(4):42-46.
[24] 刘锦春,钟章成.水分胁迫和复水对石灰岩地区柏木幼苗根系生长的影响[J].生态学报,2009,29(12):6439-6445.
[25] 王 冉,李吉跃,张方秋,等. 不同施肥方法对马来沉香和土沉香苗期根系生长的影响[J]. 生态学报,2011,31(1):0908-0106.
[26] 王力朋,晏紫伊,李吉跃,等.指数施肥对楸树无性系生物量分配和根系形态的影响[J].生态学报,2012,32(23):7452-7462.
[27] 何 茜,丁晓刚,王 冉,等. 指数施肥下黑木相思根系特征值的动态变化[J]. 广东林业科技,2011,27(5):1-6.
[28] 英 慧,殷 有,于立忠,等.土壤水分、养分对树木细根生长动态及周转影响研究进展[J].西北林学院学报,2010,25(3):36-42.
[29] 王迪海,赵 忠,张 彦. 黄土高原刺槐细根与土壤水分特征[J].西北林学院学报,2012,27(1):1-5.
[30] Bouma T J, Bryla D R. On the assessment of root and soil respiration for soils of different textures: Interactions with soil moisture contents and soil CO2concentrations[J]. Plant and Soil,2000(1-2), 227: 215-221.
[31] 卫 星,张国珍. 树木细根主要研究领域及展望[J]. 中国农学通报,2008,24(5):143-147.
[32] 梅 莉,韩有志,于水强,等.水曲柳人工林细跟季节动态及其影响因素[J].林业科学,2006,42(9):7-12.
[33] Pregitzer K S,King J S,Burton A J. Response of tree fi ne roots to temperature[J].New Phytologist,2000,147(1):105-115.
[34] 孔红岭,孙明高,孔艳菊,等. 盐分, 干旱及其交叉胁迫对皂角幼苗生长性状的影响[J]. 中南林业科技大学学报,2007,27(1): 55-59.
[35] 李树战,田大伦,王光军,等. 樟树人工林细根生物量的时空动态[J]. 中南林业科技大学学报,2008,28(6):51-55.
