不同利用方式下草地优势植物的生态化学计量特征
2014-01-02徐沙龚吉蕊张梓榆刘敏王忆慧罗亲普
徐沙,龚吉蕊,张梓榆,刘敏,王忆慧,罗亲普
(北京师范大学地表过程与资源生态国家重点实验室 北京师范大学资源学院,北京100875)
草地是世界上分布最广的植被类型之一,是陆地生态系统的重要组成部分[1]。我国草地面积达4亿hm2,占世界草地面积的13%,占全国国土面积的41.7%[2]。草地生态系统不仅为人类提供了生态服务功能,是畜牧业的生产基地,同时也是生态安全屏障保护和牧民生活与草原文化传承的基础[3-4]。由于人类活动的干扰和全球气候的变化,草地生态系统的结构、功能以及植被的演替进程和分布格局正在发生着巨大的变化[5]。放牧和割草是2种最重要的草地利用方式,不同的利用方式影响着植物的多样性,物种的组成和初级生产力以及生态系统过程[6-8],同时也影响着植物的生态化学计量特征。生态化学计量学(ecological stoichiometry)以生态学和化学计量学为基础,是研究生态系统能量和多种元素(主要指C、N、P)的平衡,以及元素平衡对生态耦合作用的影响,为研究植物的养分利用状况提供了重要的手段[9-12]。C、N、P元素是植物生长发育所必需的大量元素,在生态化学计量学的理论中,植物体内的营养元素组成处于一个稳定的动态平衡状态,当外界环境发生变化时,植物有保持自身元素组成稳定的能力[10]。植物叶片的元素特征不仅与自身的结构特点有关,还与其生长节律有着密切的关系[13]。植物体内的物质由结构性物质,功能性物质和贮藏性物质3类组成,一般情况下,C是结构性物质,N和P是功能性物质。对于同一物种来说,往往结构性物质受环境的影响较小,含量相对较稳定,而功能性和贮藏性物质的含量受环境影响变化较大[10]。当外界环境发生变化时,植物能够主动地调整养分需求,从而调整体内元素的相对含量[14]。目前,国内外学者对草地生态系统植物的生态化学计量特征进行了广泛的研究,一方面在全球水平和区域大尺度上对陆生植物叶片N、P的分布特征、变化规律及其驱动因子展开了研究[15-18];另一方面探讨了草地退化对植物和土壤养分计量特征的影响[19-24]。优势种植物是长期适应环境不断进化的结果,其体内营养元素的化学计量特征在一定程度上能反映植物对特定环境中的养分利用状况及其自身的代谢能力,同时也反映土壤中养分的供应状况,进而影响着生态系统的稳定性[25-27]。近年来对优势物种生态化学计量特征的影响也开展了一些工作[28-32],放牧和割草是草地利用的2种主要方式,放牧主要通过动物的采食、践踏以及泄物归还来影响草地土壤的养分组成,植物自身通过改变养分的利用策略来适应外界环境的变化[33]。割草能够改变土壤的水热条件和通气状况,影响微生物活动和地表凋落物的累积,进而影响土壤养分的组成[34]。鉴于此,本研究将内蒙古温带草原3种利用方式(放牧、割草和围封)结合起来,对3种不同的利用方式下3种优势种植物的化学计量特征进行了研究,具体目标如下:1)探讨不同草地利用方式下优势种植物养分含量及化学计量比的季节变化;2)分析不同的草地利用方式对土壤化学计量特征产生的影响;3)优势种植物在不同利用方式下的养分限制状况。
1 材料与方法
1.1 研究区概况
本研究在距内蒙古锡林浩特市东部36km的内蒙古大学草地生态系统研究站(116°02′~116°30′E,44°48′~44°49′N)长期试验样地进行的。该地属温带干旱大陆性气候,年平均气温0~1℃,年积温1800℃左右,年降水量300~360mm,降雨多集中在6-8月,无霜期90~115d,具有光、热、水同期特点。土壤以栗钙土为主和少量褐色土,软栗钙土层15~80cm,土壤有机质含量2%~3%,土质比较肥沃。地带性植被的基本类型是大针茅草原。分别设置围封、割草、放牧3个不同草地利用方式的样地,每个样地面积为14hm2,围封样地自2008年起被围住;割草是每年8月进行1次;放牧样地则是常年放养牲畜,且放牧地使用年限已超过50年,强度为3.67hm2/头。3个实验样地的海拔均在1055m。优势种植物重要值(表1)计算公式为:
重要值=(相对密度+相对频度+相对盖度)/3[35]
1.2 样品的采集与测定
于2011年6,7,8和9月在每个样地随机布设1个50m的样带,每隔10m设置1个1m×1m的样方,共5个样方。齐地面刈割地上部分,选取每个样方内的3种优势种植物大针茅、羊草和冷蒿,同种植物混合到一起。各植物样品在105℃下杀青15min,65℃烘干至恒重,冷却后用研磨仪研磨用于各指标的测定。于8月,每个样方用土钻在0~40cm土壤内每隔10 cm取土,3次重复,带回实验室在105℃下烘干至恒重后用研钵磨碎装袋。分别测定每种植物叶片和每层土壤的全碳、全氮和全磷,每个指标重复3次。C、N的含量在北京师范大学化学分析实验中心用德国Elementar公司生产的Vario EL元素分析仪测量,P的含量用ICP-AES光谱仪测定。另外,土壤有机质含量采用重铬酸钾容量法测定[36]。

表1 3种不同类型样地优势种重要值Table 1 The importance value of dominant species of three different style plots
1.3 数据处理及分析方法
利用SPSS 19.0进行ANOVA方差分析处理,对不同草地利用方式下不同优势种植物间的差异性,再用单因素方差分析进行LSD多重比较,使用Origin 8.5作图。
2 结果与分析
2.1 不同利用方式下优势种植物叶片C、N、P含量及其计量比的季节变化
在放牧条件下,3种优势植物叶片C、N、P含量及其计量比存在着明显的季节变化(图1)。对于植物叶片C含量,3种植物均在整个生长季内略有变化,总体上C含量比较稳定(P>0.05)(图1A)。3种植物叶片N、P含量变化基本一致,均呈先增加后降低的趋势,且不同月份之间差异显著(P<0.05),只有大针茅的N含量是随时间推进逐渐降低的(图1A,B)。对于植物叶片C∶N,在整个生长季内叶片总体上呈增加的趋势,并且三者均在8至9月明显增加,其中,羊草C∶N 9月明显高于冷蒿和大针茅(图1D)。对于植物叶片C∶P和N∶P,大针茅在6月最高并极显著高于冷蒿(P<0.01),冷蒿的N∶P在6月也相对较高(P<0.05)(图1E,F)。
在割草条件下,3种植物的C、N、P含量季节变化差异比较大(图2A~C),冷蒿和大针茅叶片C含量的季节波动不大(P>0.05),而羊草则是逐渐降低的(P<0.05),叶片N、P含量的变化总体上和放牧条件下的基本一致,季节变异较显著(P<0.05)。对于植物叶片C∶N,3种植物均呈增加的趋势,而冷蒿叶片C∶P和N∶P,6月极显著高于其他月份(P<0.01)(图2D,F)。
围封条件下,3种植物叶片的N、P含量以及C∶N、C∶P和N∶P季节波动均较明显(P<0.05)(图3),只有C含量在整个生长季的变化相对较稳定(P>0.05)。叶片N含量和C∶N的变化是一致的,这也说明了叶片C含量是比较稳定的。对于叶片C∶P和N∶P来说,6月冷蒿和大针茅均显著高于其他月份(P<0.01),这与二者P含量在6月较低是对应的。

图1 放牧条件下3种优势种植物叶片化学计量特征的季节变化Fig.1 The seasonal change of stoichiometric characteristic of three kinds of dominant species in grazing plot
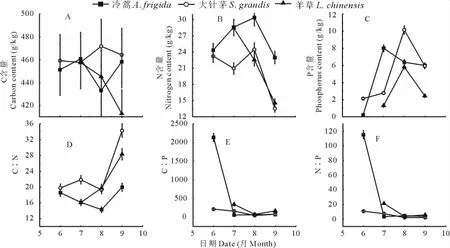
图2 割草条件下3种优势种植物叶片化学计量特征的季节变化Fig.2 The seasonal change of stoichiometric characteristic of three kinds of dominant species in mowing plot

图3 围封条件下3种优势种植物叶片化学计量特征的季节变化Fig.3 The seasonal changes of stoichiometric characteristic of three dominant species in the grazing exclusion plot
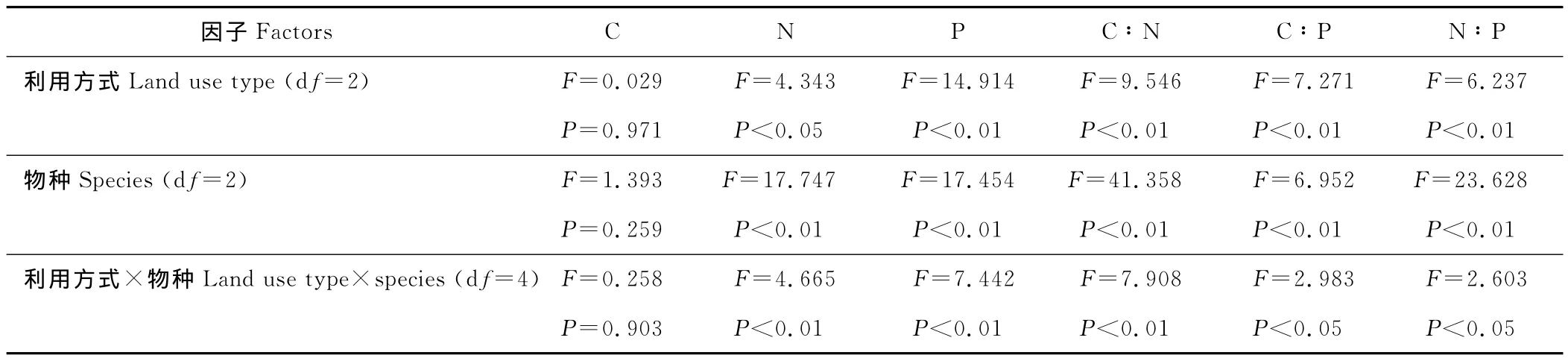
表2 不同的利用方式和不同物种对叶片养分含量及其计量比的分析Table 2 Two-way ANOVA results for effects of Species,land use type on leaf nutrient content and its mass ratio
2.2 不同利用方式对土壤养分含量及其计量比的影响
全N含量随着土层的加深总体上呈降低趋势(图4B),土壤全碳只有在放牧条件下随土层加深略有变化,其他均保持基本不变(图4B),其中,土壤全C和全N含量在每个土层均是放牧样地高于割草和围封样地,并且在每个土层3种样地之间基本差异显著。对于土壤全P含量,各土层在3种利用方式下均差异不显著(图4D),除了30~40cm土层,其他土层均是放牧样地P含量高于其他2个样地。在0~10cm,土壤C∶N在3个样地之间无显著差异 (图4E),随土层加深,3个样地均略有增加,在30~40cm土层,放牧样地C∶N显著高于割草和围封样地。土壤C∶P和N∶P总体上均是放牧样地高于其他2个样地(图4F,G),其中,土壤N∶P在放牧与割草样地之间基本无显著差异,围封样地则与二者有一定的差异。
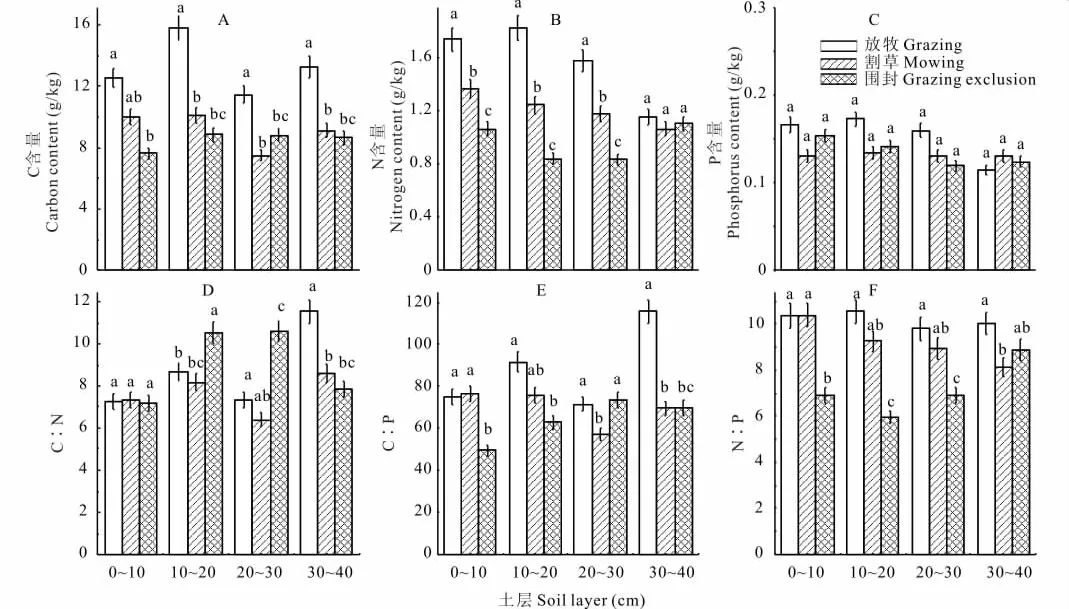
图4 不同土层土壤在不同利用方式下的养分含量及其计量比Fig.4 The soil nutrient content and its mass ratio in different soil layers under the different land use types
3 讨论
3.1 不同利用方式下优势种植物的化学计量特征响应机制
植物体内的物质由结构性物质、功能性物质和贮藏性物质3类组成,一般情况下,C是结构性物质,N和P是功能性物质。对于同一物种来说,往往结构性物质受环境的影响较小,含量相对较稳定,而功能性和贮藏性物质的含量受环境影响变化较大[10]。本研究中C作为结构性物质在3种植物中的季节变异比较小,略有变化,并且不同的利用方式对C含量的影响也较小,这与上述规律较一致。叶片N、P含量在放牧、割草和围封条件下,3种植物均有较大的季节变化,N含量在整个生长季内总体上呈下降的趋势。植物在生长季推移的过程中,叶片中的N、P浓度有一个稀释效应[37-38],因为叶片中的N含量决定着植物的光合作用,植物在光合作用过程中需要大量蛋白酶的参与,N是组成蛋白酶的重要元素,而蛋白酶的形成又需要大量的遗传物质核酸的复制与转录,其中P是遗传物质组成的一部分[10]。本研究中只有N含量的季节变化符合上述规律,P则不符合,可能是由于物种本身的特性造成的。另外,无论在放牧、割草还是围封条件下,冷蒿和大针茅叶片P含量在6月表现相当低,这可能是由于6月植物的生长受到P的限制。根据Koerselman和Meuleman[39]的研究,当N∶P小于14时,表示生态系统是受N限制;N∶P大于16时,表示生态系统是受P限制;当N∶P在14~16之间时,生态系统同时受N和P的限制或者同时不受二者限制。在6月,3种利用方式中除了放牧条件下的冷蒿,冷蒿和大针茅的N∶P均大于16,说明2种植物在6月基本是受到了P的限制,才导致了P含量偏低。植物吸收N、P营养元素的途径和C同化不同,一般情况下,C的含量比较稳定,变异较小,C∶N和C∶P的变化主要由N、P的变化决定[40]。本研究中,3种植物C∶N和C∶P的季节变化在3种利用方式下均表现为与N、P的季节变化规律相反,这说明N、P的变化决定了C∶N和C∶P的变化,与上述规律一致[41-42]。N∶P的季节变化与P含量的变化也是相反的规律,说明N∶P的变化主要由P的变化决定[25]。根据N∶P的季节变化,在放牧条件下,除了大针茅在6月受到P的限制作用外,3种植物在整个生长季均受到N的限制。割草条件下,冷蒿和大针茅在6月,羊草在7月受P的限制,其他均受N的限制。围封条件下养分限制的规律和割草是一致的。
3.2 不同利用方式对土壤养分特征的影响
土壤作为植物体营养元素的主要来源,其养分的变化和分布情况会对植物体的生长发育产生很大的影响[43]。目前,草地生态系统越来越多地受到人类活动的干扰,在人类管理草原的过程中,放牧、围封和割草是3种主要的管理方式,影响着草地生态系统过程。不同的管理方式不仅影响着植物的化学计量特征,同时对土壤的养分特征也有一定的影响,各土层每种养分指标在3种利用方式间均有一定的差异。本研究中0~30cm土层内放牧和割草条件下各土层有机质含量均高于围封条件,说明放牧和割草在一定程度上增加了土壤有机质的含量,总体上,放牧样地是高于其他2个样地的,主要由于牲畜排出的粪便增加了土壤中容易分解的有机物,从而增加了土壤的有机质[44]。土壤全C含量放牧样地显著高于割草和围封,这可能是由于放牧增加了土壤中的有机物所致,在表层土壤(0~20cm),割草样地土壤全C含量高于围封样地,可能是由于割草样地地上生物量的减少,导致植物生长加速,并使得地下生物量增加,从而使土壤全C增加,土壤中全C来源于地上生物量和根系生物量的归还[45],而围封样地植物生长较缓慢,地上和地下生物量的归还相对较少。土壤N含量主要来源于凋落物归还、大气沉降及一些固氮植物,土壤P含量则主要来源于岩石的风化及凋落物归还,凋落物归还N过程类似于P归还过程,而岩石的风化则是一个漫长的过程,相对较稳定[46]。不同的利用方式也影响着土壤的N、P含量,在0~30cm土层内,放牧样地显著高于其他两样地,主要由于食草动物的排泄物所致,这与安慧和徐坤[47]的研究中放牧样地土壤N含量大于围封样地是一致的。对于土壤30~40cm土层各养分含量在不同利用方式间基本无显著差异,可能是由于深层土壤受干扰的影响较小造成的。放牧样地N∶P总体上同样高于其他2个样地,这与土壤N、P含量的特征密切相关。本地区土壤C∶N的变化范围为7.19~11.53,低于我国C∶N的平均值10~12[48]。有研究表明,土壤C∶N大于30或小于30分别是硝酸盐淋溶风险低或高的阈值[49],因此可以判断,本地区土壤有较高的硝酸盐淋溶风险,有可能造成土壤N的损失,这与本研究植物普遍受到N的限制是一致的。对于土壤C∶P,一方面,土壤C∶P的高低对植物生长发育具有重要影响。如果C∶P较低,则有利于微生物在有机质分解过程中的养分释放,促进土壤中有效磷的增加;反之,C∶P较高,则会出现微生物在分解有机质的过程中存在磷受限,从而与植物存在对土壤无机磷的竞争,不利于植物的生长及NPP的增加[50]。有研究表明,当C∶P<200时将会出现土壤微生物碳的短暂增加和有机磷的净矿化,当C∶P>200时微生物碳大幅增加,微生物竞争土壤中的速效磷,出现有机磷的净固持现象[51]。本地区土壤C∶P整体上均小于200,因此会出现有机磷的净矿化。
[1]赵同谦,欧阳志云,贾良清,等.中国草地生态系统服务功能间接价值评价[J].生态学报,2004,24(6):1101-1110.
[2]Fan J,Zhong H,Harris W,etal.Carbon storage in the grasslands of China based on field measurements of above-and belowground biomass[J].Climatic Change,2008,86(3-4):375-396.
[3]于格,鲁春霞,谢高地.草地生态系统服务功能的研究进展[J].资源科学,2006,27(6):172-179.
[4]赵有益,龙瑞军,林慧龙,等.草地生态系统安全及其评价研究[J].草业学报,2008,17(2):143-150.
[5]任书杰,于贵瑞,陶波,等.中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J].环境科学,2008,28(12):2665-2673.
[6]Bai Y,Wu J,Pan Q,etal.Positive linear relationship between productivity and diversity:evidence from the Eurasian Steppe[J].Journal of Applied Ecology,2007,44(5):1023-1034.
[7]Sasaki T,Okayasu T,Jamsran U,etal.Threshold changes in vegetation along agrazing gradient in Mongolian rangelands[J].Journal of Ecology,2008,96(1):145-154.
[8]Smith M D,Knapp A K.Dominant species maintain ecosystem function with non-random species loss[J].Ecology Letters,2003,6(6):509-517.
[9]Elser J J,Dobberfuhl D R,MacKay N A,etal.Organism size,life history,and N∶P stoichiometry[J].BioScience,1996,46(9):674-684.
[10]Sterner R W,Elser J J.Ecological Stoichiometry:the Biology of Elements from Molecules to the Biosphere[M].Princeton:Princeton University Press,2002.
[11]王振兴.国内生态化学计量学研究进展[J].绿色科技,2011,(7):195-196.
[12]林新坚,王飞,王长方,等.长期施肥对南方黄泥田冬春季杂草群落及其C,N,P化学计量的影响[J].中国生态农业学报,2012,20(5):573.
[13]Aerts R,Chapin III F S.The mineral nutrition of wild plants revisited:a re-evaluation of processes and patterns[J].Advances in Ecological Research,1999,30:1-67.
[14]曾德慧,陈广生.生态化学计量学:复杂生命系统奥秘的探索[J].植物生态学报,2005,29(6):1007-1019.
[15]Reich P B,Oleksyn J.Global patterns of plant leaf N and P in relation to temperature and latitude[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101(30):11001-11006.
[16]Han W,Fang J,Guo D,etal.Leaf nitrogen and phosphorus stoichiometry across 753terrestrial plant species in China[J].New Phytologist,2005,168(2):377-385.
[17]He J S,Wang L,Flynn D F B,etal.Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes[J].Oecologia,2008,155(2):301-310.
[18]任书杰,于贵瑞,姜春明,等.中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征[J].应用生态学报,2012,23(3):581-586.
[19]Frank D A,Evans R D.Effects of native grazers on grassland N cycling in Yellowstone National Park[J].Ecology,1997,78(7):2238-2248.
[20]银晓瑞,梁存柱,王立新,等.内蒙古典型草原不同恢复演替阶段植物养分化学计量学[J].植物生态学报,2010,34(1):39-47.
[21]罗亚勇,张宇,张静辉,等.不同退化阶段高寒草甸土壤化学计量特征[J].生态学杂志,2012,31(2):254-260.
[22]林丽,李以康,张法伟,等.高寒矮嵩草群落退化演替系列氮,磷生态化学计量学特征[J].生态学报,2013,33(17):5245-5251.
[23]陈军强,张蕊,侯尧宸,等.亚高山草甸植物群落物种多样性与群落C,N,P生态化学计量的关系[J].植物生态学报,2013,37(11):979-987.
[24]丁小慧,罗淑政,刘金巍,等.呼伦贝尔草地植物群落与土壤化学计量学特征沿经度梯度变化[J].生态学报,2012,32(11):3467-3476.
[25]杨阔,黄建辉,董丹,等.青藏高原草地植物群落冠层叶片氮磷化学计量学分析[J].植物生态学报,2010,34(1):17-22.
[26]牛得草,董晓玉,傅华.长芒草不同季节碳氮磷生态化学计量特征[J].草业科学,2011,28(6):915-920.
[27]马剑英,陈发虎,夏敦胜,等.荒漠植物红砂叶片元素和水分含量与土壤因子的关系[J].生态学报,2008,28(3):983-992.
[28]LüX T,Kong D L,Pan Q M,etal.Nitrogen and water availability interact to affect leaf stoichiometry in a semi-arid grassland[J].Oecologia,2012,168(2):301-310.
[29]LüX T,LüF M,Zhou L S,etal.Stoichiometric response of dominant grasses to fire and mowing in a semi-arid grassland[J].Journal of Arid Environments,2012,78:154-160.
[30]宾振钧,王静静,张文鹏,等.氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响[J].植物生态学报,2014,38(3):231-237.
[31]安卓,牛得草,文海燕,等.氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C∶N∶P化学计量特征的影响[J].植物生态学报,2011,35(8):801-807.
[32]董晓玉.放牧与围封对黄土高原典型草原植物碳,氮,磷生态计量特征及其贮量的影响[D].兰州:兰州大学,2009.
[33]张婷,翁月,姚凤娇,等.放牧强度对草甸植物小叶章及土壤化学计量比的影响[J].草业学报,2014,23(2):20-28.
[34]郭明英,朝克图,尤金成,等.不同利用方式下草地土壤微生物及土壤呼吸特性[J].草地学报,2012,20(1):42-48.
[35]汤诗杰,彭志,汤庚国.宝华山南京椴群落的特征分析[J].扬州大学学报(农业与生命科学版),2008,29(1):90-94.
[36]鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000.
[37]Santa Regina I,Rico M,Rapp M,etal.Seasonal variation in nutrient concentration in leaves and branches ofQuercus pyrenaica[J].Journal of Vegetation Science,1997,8(5):651-654.
[38]Sardans J,Pe uelas J.Drought changes nutrient sources,content and stoichiometry in the bryophyteHypnumcupressiformeHedw.growing in a Mediterranean forest[J].Journal of Bryology,2008,30(1):59-65.
[39]Koerselman W,Meuleman A F M.The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation[J].Journal of Applied Ecology,1996,33(6):1441-1450.
[40]Hedin L O.Global organization of terrestrial plant-nutrient interactions[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101(30):10849-10850.
[41] gren G I.Stoichiometry and nutrition of plant growth in natural communities[J].Annual Review of Ecology,Evolution,and Systematics,2008,39:153-170.
[42]牛得草,李茜,江世高,等.阿拉善荒漠区6种主要灌木植物叶片C∶N∶P化学计量比的季节变化[J].植物生态学报,2013,37(4):317-325.
[43]刘兴诏,周国逸,张德强,等.南亚热带森林不同演替阶段植物与土壤中N,P的化学计量特征[J].植物生态学报,2010,34(1):64-71.
[44]高英志,韩兴国,汪诗平.放牧对草原土壤的影响[J].生态学报,2004,24(4):790-797.
[45]Ren H Y,Zheng S X,Bai Y F.Effects of grazing on foliage biomass allocation of grassland communities in Xilin River Basin,Inner Mongolia[J].Journal of Plant Ecology(Chinese Version),2009,33(6):1065-1074.
[46]刘万德,苏建荣,李帅锋,等.云南普洱季风常绿阔叶林演替系列植物和土壤C,N,P化学计量特征[J].生态学报,2010,30(23):6581-6590.
[47]安慧,徐坤.放牧干扰对荒漠草原土壤性状的影响[J].草业学报,2013,22(4):35-42.
[48]黄昌勇,李保国,潘兴根,等.土壤学[M].北京:中国农业出版社,2000.
[49]Gundersen P,Callesen I,DeVries W.Nitrate leaching in forest ecosystems is controlled by forest floor C/N ratio[J].Environmental Pollution,1998,102(1):403-407.
[50]王建林,钟志明,王忠红.青藏高原高寒草原生态系统土壤碳磷比的分布特征[J].草业学报,2014,23(2):9-19.
[51]王绍强,于贵瑞.生态系统碳氮磷元素的生态化学计量学特征[J].生态学报,2008,28(8):3937-3947.
