半干旱沙地生境变化对植物地上生物量及其碳、氮储量的影响
2014-01-02周欣左小安赵学勇王少昆罗永清岳祥飞张腊梅
周欣,左小安,赵学勇,王少昆,罗永清,2,岳祥飞,2,张腊梅,2
(1.中国科学院寒区旱区环境与工程研究所奈曼沙漠化研究站,甘肃 兰州730000;2.中国科学院大学,北京100049)
在生态系统中,群落生物量作为生态学研究中一种重要的数量特征能够衡量植被的动态和生态系统功能的变化[1-2]。生态系统中植被的碳主要来源于植物通过光合作用固定大气中的CO2;碳含量反映了植物的光合碳同化能力,是植物碳储量的一种度量[3]。氮元素作为构建植物光合器官的重要营养元素,决定着植物生长和植被生态系统的发育。光合器官的氮含量对植物适应干旱环境具有重要的作用[4]。植物碳、氮含量及其储量与生态系统的物质周转和养分循环具有密切的联系,主要受植物生长型、植物构件、生境变化及演替时间等多种因素的影响[5-6]。而且,植被碳、氮储量的估算也是揭示植被恢复对陆地生态系统影响的核心内容之一[7]。中国沙漠化和沙地面积分别约占国土面积的17%和6%[8],因此,研究沙漠化逆转过程和沙地退化植被过程中的植被生物量和碳氮储量变化及其植被碳氮固存潜力对区域生态系统恢复和碳氮平衡具有重要理论和实践意义[9]。目前,有关这方面的研究主要集中在人工固沙植被生物量的变化[1,10-11]、沙地植被退化或恢复过程中的碳氮变化[3,12-14]等,而缺少半干旱沙地沙丘固定过程中生境变化对植被碳氮储量及其固存潜力影响的报道[15]。
科尔沁沙地沙丘固定过程中的不同生境对植物有较大的影响,表现在生境对群落物种组成,植被生物量变化以及植物功能型组成等方面的影响[16]。然而,沙地沙丘固定过程中生境变化对群落组成的生活型、C3和C4植物功能型的生态功能研究较少。因此,研究沙丘固定过程中的流动沙丘、半固定沙丘和固定沙丘以及草地4种生境类型的植物地上部分生物量及碳、氮储量特征,旨在为沙地退化生态系统的恢复重建与可持续管理提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于科尔沁沙地中南部,隶属内蒙古自治区通辽市奈曼旗境,地理位置120°55′E,42°41′N,平均海拔约为360m。该区域属温带大陆性半干旱季风气候,年平均气温6.4℃,≥10℃年积温在3000℃以上,无霜期约150d。年平均降水量364.6mm,主要集中在6,7,8三个月,年平均蒸发量1972.8mm,年平均风速3.5m/s;研究区地带性土壤为沙质栗钙土,但因长期的风蚀作用已退化为风沙土。该区的地貌类型以平缓的流动沙丘、半固定沙丘、固定沙丘和草地等镶嵌分布为特征。优势植物种有沙米(Agriophyllumsquarrosum)、砂蓝刺头(Echinopsgmeliniturcz)、冷蒿(Artemisiafrigida)、糙隐子草(Cleistogenessquarrosa)、差巴嘎蒿(Artemisia halodendron)和小叶锦鸡儿(Caraganamicrophylla)等。
1.2 研究方法
1.2.1 样地选择和样品采集 于2011年8月中旬植物达到最大生物量期间,选择沙丘长期封育固定过程中的流动沙丘、半固定沙丘和固定沙丘以及封育的草地4种生境类型作为研究对象,每个生境类型设置3个重复,生境类型间的距离为0.5~10.0km。在每个生境类型上建立20m×20m坡度较为平缓(0°~5°)的典型样地。应用该区域常用植被调查方法[2,12,16],在每个样地中随机设置5个1m×1m的样方,用于调查记录每个样点的植物种类、高度、盖度等。在每个样方中用收割法,分物种获取每种植物,装入纸袋带回实验室。样方中每种植物按照叶和枝(包括极少量的茎)分类,并在65℃下将样品烘干至恒重,分别测得叶干重与枝干重,二者之和即为该物种生物量;最终将每块样地15个样方的生物量平均值代表该样地的地上生物量。根据物种i的生物量占样方总生物量的比值计算出该物种的权重pi;叶干重占物种i生物量的比重为ai,枝干重的比重为bi(ai+bi=1)。
1.2.2 植物碳、氮含量的测定 为保证每种植物有足够的叶和枝用以测定样品的碳氮含量,在每个样方附近获取部分植物的叶和枝,带回实验室烘干称重后,与原来每个样方烘干称重后的样品混合,制成分析植物碳氮含量的样品。再将每个植物叶与枝的样品混合粉碎,过0.25mm筛,储存备用。分析前在65℃的干燥箱中烘干1h,放入干燥器中保存待测。采用元素分析仪(vario Macro cube,Elementar,Germany)测定样方中每种植物叶和枝样品的碳氮含量。
1.2.3 植物功能性分类 将4种生境的植物种划分为不同的功能型:按照植物种生活型划分为一年生草本(包含一年生或二年生草本)、多年生草本和灌木3种类型;根据不同光合作用途径分为C3和C4植物[17]。
1.2.4 碳氮含量及储量的计算 以碳含量及储量为例,每个样地中样方叶碳含量为物种i在样方中的权重,qi为物种i的叶碳含量)[18-19],样方叶碳储量(leaf carbon stock,LCS)为该样方地上生物量与叶碳含量的乘积;枝碳含量(branch carbon content,BC)及储量(branch carbon stock,BCS)的算法同理;样方总碳含量(total carbon content,TC)为物种i的枝碳含量,括号内的计算值即为物种i的碳含量;样方总碳储量(total carbon stock,TCS)=LCS+BCS。按生活型、功能型分析的植物碳氮含量均以各物种碳氮含量乘以其权重为数据源。
1.3 数据处理
每个样方中每种植物的生物量由叶干重和枝干重求和获得;每种植物的碳氮含量由叶和枝的碳氮含量乘以其占该样方植物生物量的比重后加权平均获得;每个样方的植物群落碳氮含量由每种植物的碳氮含量乘以其占该样方生物量的比重后加权平均获得。用单因素方差分析(ANOVA)对不同生境之间的生物量及其碳氮含量进行差异显著性分析,用最小显著性差异(LSD)多重比较方法,在95%的可靠性下对不同生境、不同植物组织、不同生活型、不同光合途径的植物功能型之间进行分析比较其差异性,所有数值均采用平均值±标准误(SE)表示;数据差异显著性分析采用SPSS 17.0软件。
2 结果与分析
2.1 沙地不同生境群落组成的变化
沙地4种不同生境的植物组成和重要值变化如表1所示。在固定沙丘和草地上,黄蒿均为优势种,分别占其相应群落总优势度的39.59%和47.20%;半固定沙丘的优势种为差巴嘎蒿,占群落总优势度的44.07%;在流动沙丘上,一年生草本植物沙米为优势种,重要值为61.86%。

表1 沙地不同生境植物种类组成、光合途径及重要值Table 1 Vegetation species composition,photosynthetic pathways and important values among four different sandy habitats%
2.2 沙地不同生境植物生物量及碳氮分配特征
沙地不同生境的叶干重、枝干重、总生物量及其碳氮含量与储量(表2)具有较大的差异(P<0.01)。固定沙丘的叶干重、枝干重、总生物量及其碳氮储量均显著大于(P<0.05)流动沙丘与半固定沙丘;固定沙丘的总生物量、总碳氮储量分别是流动沙丘的23.94,24.41,22.17倍,是半固定沙丘的3.84,3.54,5.15倍。草地的叶干重、枝干重、总生物量、枝碳储量、总碳储量、枝氮储量均显著小于(P<0.05)固定沙丘,而大于流动沙丘;草地的总生物量是固定沙丘的53.09%,是流动沙丘的12.71倍。半固定沙丘叶碳含量、枝碳含量、总碳含量、枝碳储量均显著大于流动沙丘(P<0.05),分别高出9.40%,10.42%,10.24%,10.31%;而叶氮含量、枝氮含量、总氮含量均小于流动沙丘(P<0.05),分别低了27.66%,36.11%,37.77%。

表2 沙地不同生境植物生物量及碳氮分配特征(平均值±标准误)Table 2 Plant biomass,carbon and nitrogen distribution characteristics among 4different habitats(Mean±SE)
2.3 不同生活型植物生物量及碳氮分配
从生物量的生活型组成来看(图1),除流动沙丘外,其他各生境不同生活型植物之间存在极显著差异(P<0.01)。在流动沙丘、固定沙丘和草地3种生境上,一年生植物是主要组成部分(一年生植物生物量占总生物量的比值分别为63.99%,79.28%和70.86%);半固定沙丘上的主要为灌木,占73.15%。其中,一年生植物生物量随着沙丘的固定(流动沙丘-半固定沙丘-固定沙丘)依次递增,由9.03g/m2增加到127.15g/m2。
除流动沙丘外,其他各生境不同生活型植物碳含量之间也存在极显著差异(P<0.01)。随着沙丘的固定,一年生和多年生草本的碳含量依次增加,分别由流动沙丘的36.69%和37.40%增至草地的42.81%和43.38%。半固定沙丘的灌木碳含量(45.68%),显著高于(P<0.05)流动沙丘(43.6%)和固定沙丘(42.43%)。4种生境不同生活型植物氮含量之间均存在差异,其中半固定沙丘和草地差异极显著(P<0.01)。灌木氮含量在草地最大(2.19%),分别高出流动沙丘、半固定沙丘、固定沙丘90.27%,97.94%,37.47% (P<0.05)。除固定沙丘外,4种生境不同生活型植物碳氮比均存在极显著差异(P<0.01)。一年生植物碳氮比(19.15~31.56)在流动沙丘最小;多年生草本碳氮比(18.73~26.72)在半固定沙丘和固定沙丘最小;灌木碳氮比(20.70~41.73)在草地最小。
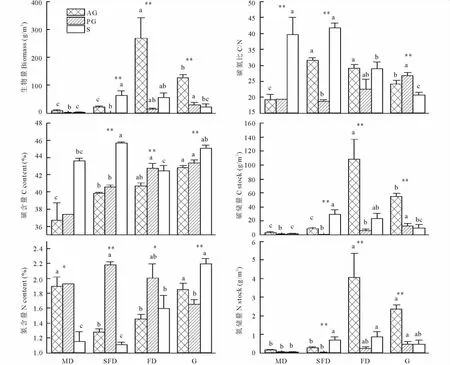
图1 沙地4种生境上不同生活型植物生物量、碳氮分配及储量Fig.1 Biomass,allocation and stock of C and N among different plant life forms
一年生、多年生草本碳储量分别在固定沙丘(108.25g/m2)、草地(12.73g/m2)最高;固定沙丘一年生植物、灌木的碳储量分别是流动沙丘的31.76和16.45倍(P<0.05)。固定沙丘一年生植物和灌木的氮储量(分别为4.04和0.85g/m2)显著大于(P<0.05)流动沙丘(分别为0.14和0.04g/m2);草地上多年生草本氮储量(0.47 g/m2)是半固定沙丘的16.79倍。半固定沙丘灌木氮储量是多年生草本的24.75倍,固定沙丘的一年生植物是多年生草本的16.31倍(P<0.05)。除流动沙丘外,不同生活型植物碳、氮储量之间存在极显著差异(P<0.01)。
2.4 不同光合途径植物生物量及碳氮分配
从图2可以看出,除流动沙丘外,其余3个生境C3植物生物量极显著大于C4植物(P<0.01);固定沙丘中C3植物生物量为C4植物的24.04倍。流动沙丘、半固定沙丘、固定沙丘和草地C4植物功能群占其群落的比重分别是60.01%,9.13%,4.00%和17.35%。

图2 沙地4种生境上不同光合途径植物生物量、碳氮分配及储量Fig.2 Biomass,allocation and stock of C and N between plants with different photosynthetic pathways
流动沙丘、半固定沙丘、草地的C3植物碳含量极显著高于C4(P<0.01);除草地外,其他3个生境C3植物氮含量均小于C4(P<0.05),而C3植物碳氮比大于C4(P<0.01)。随着沙丘的固定,C4植物的碳含量逐渐增大,由流动沙丘的38.54%增大到草地的40.42%。草地C3植物氮含量最大(1.85%),半固定沙丘C3植物氮含量(1.14%)最小。半固定沙丘C3及C4植物碳氮比显著高于其他各生境(P<0.05)。2种碳型植物碳含量只在固定沙丘上无显著差异(P>0.05),而氮含量和碳氮比2项指标只在草地上差异不显著。
除流动沙丘外,其他3种生境上C3、C4植物碳、氮储量差异极显著(P<0.01)。C3植物在固定沙丘的碳、氮储量分别是流动沙丘的57.06和54.18倍,C4植物在草地上的碳、氮储量分别是半固定沙丘的3.98和4.91倍。
3 讨论
据统计科尔沁沙地植被净初级生产力近十年间总体呈上升趋势[20],在沙地退化植被恢复过程中,生物量及其碳氮储量的变化存在着显著的差异。固定沙丘的地上部分生物量显著高于其他生境,表明植被组成较好且相对稳定的固定沙丘植被生产力较大,因此,固定沙丘的植物碳储量较高。草地作为沙地生态系统中较为稳定的生境类型,其植物地上生物量显著小于固定沙丘,这与科尔沁沙地生境及其资源的高度空间异质性有关,土壤水肥资源的空间异质性分布决定了植物的分布及其种间竞争[16],进而影响了植物群落的分布和演替及其碳氮储量的变化。
沙丘的固定促进了植被的演替,进而导致植物C、N含量等的差异。生物量的分配受植物遗传特性等影响,其动态变化是植物与环境相互作用的重要标志,体现了环境影响下植物生长发育的特点[21]。本实验枝、叶干重的比例在流动沙丘、半固定沙丘、固定沙丘、草地上依次是0.83,2.31,1.38,1.01,说明植物为适应环境的改变调节着自身的生长,叶的比重高表明植物光能利用效率高,枝则是植物体的巨大的能量贮藏库及调节器。因各植物根系吸收土壤元素的能力各异,同时受凋落物、根系等分解释放到土壤的影响,植物元素含量亦不同。本研究与郑帏婕等[7]研究结果相一致,表明植物不同组织的碳含量表现为枝高于叶。本试验初步表明各生境中氮素在植物叶片的含量较高,已有研究表明植物叶片氮含量[22]对植物的生产力和适应干旱环境的能力具有重要影响。
群落生活型组成的变化反映出沙丘固定和退化植被恢复过程中群落结构与功能的变化特征。从生活型组的构成变化来看,一年生植物是流动沙丘、固定沙丘和草地3种生境上的优势植物,灌木是半固定沙丘上的主要生活型。尽管流动沙丘有少量的一年生植物,但其死亡形成的凋落物及根系在流动沙丘上具有一定的防风固沙和碳氮增汇作用。一年生植物生物量及碳、氮储量均随着沙丘的固定(流动沙丘-半固定沙丘-固定沙丘)依次递增。一年生植物是沙地生态系统重要的初级生产者,由于其在时空分布上的独特性,使其对恢复沙地生态系统生产力及防止荒漠化具有重要作用[23]。流动沙丘、半固定沙丘和草地生境碳含量均是灌木最大,这主要是因为灌木多数是双子叶植物,其根、枝等有次生结构而草本植物多为单子叶植物,在营养器官的构建上消耗较少,所以灌木碳含量高于以单子叶植物为主的草本类植物。流动及半固定沙丘上,一年生、多年生草本氮含量较灌木高。氮含量高有利于提高植物光合作用能力和水分利用效率,草本植物具有更强的资源利用能力[24]。一年生草本植物的生长与水分条件密切相关,而多年生草本比一年生草本有更强的抗逆境、保持种群稳定的能力[25]。在半固定沙丘上,灌丛可有效拦截风蚀、水蚀产物,有利于凋落物富集[26];差巴嘎蒿作为优势种经适度沙埋可促其萌发出大量不定根和新枝,对维持半固定沙丘群落功能有重要作用[27]。
韩梅等[28]研究中国东北样带温带草原的草地群落发现,C4植物生物量随环境干旱化等程度的加重呈增加趋势。本研究中C4植物功能群生物量也具有明显的变化规律,对环境变化响应显著。沙丘固定过程中C3和C4植物生物量与群落总生物量关系得到,C3植物生物量占群落的比例随着沙丘的固定依次增加,而C4植物的比例依次递减,C3植物起到了补偿C4植物生物量的降低的作用。C4植物与C3植物相比,具有高净光合速率、高水分利用效率、高氮素利用效率等特点,具有抗逆性强、耐放牧干扰等优势[29]。一些C4植物往往是植被恢复演替的先锋种,如试验中流动沙丘的沙米就是固沙先锋C4植物。此外,在4种不同的生境中,C4植物氮含量均大于C3植物,这主要是由于C4植物枝的氮含量均大于C3植物所导致的,而植物叶氮含量仅在半固定沙丘和草地上表现为C3植物大于C4植物。
4 结论
本研究表明,植物的地上生物量(包括叶干重、枝干重、总生物量)及其碳氮含量与储量对不同的沙地生境有响应并具有显著差异。1)不同生境的叶干重、枝干重、总生物量及其碳氮含量与储量均有极显著差异(P<0.01)。2)随着沙丘的固定,流动沙丘、半固定沙丘的优势种分别为沙米和差巴嘎蒿,黄蒿为固定沙丘和草地的优势种。一年生植物是流动沙丘、固定沙丘和草地3种生境的优势生活型,半固定沙丘上主要为灌木;固定沙丘一年生植物的生物量及碳氮储量在4个生境中最高,一年生植物有较强的固存碳氮的潜力。3)除流动沙丘外,其余3个生境C3、C4植物生物量及碳、氮储量差异极显著(P<0.01)。固定沙丘C3植物的生物量及碳氮储量在4个生境中最高;半固定沙丘、固定沙丘和草地的C3植物碳含量、碳氮储量均高于C4植物,C3植物能较有效地固存碳氮。沙丘固定过程中,C4植物生物量占群落的比例依次递减,C3植物依次递增,C3植物起到了补偿C4植物生物量的作用。
[1]韩彬,樊江文,钟华平.内蒙古草地样地植物群落生物量的梯度研究[J].植物生态学报,2006,30(4):553-562.
[2]常学礼,鲁春霞,高玉葆.科尔沁沙地不同沙漠化阶段植物种多样性与沙地草场地上生物量关系研究[J].自然资源学报,2003,18(4):475-482.
[3]高伟.退化草地生态系统碳-氮变化规律及氮添加的生态效应研究[D].呼和浩特:内蒙古大学,2010.
[4]苗艳明,吕金枝,毕润成.不同功能性植物叶氮含量与光合特性的关系研究[J].植物研究,2012,32(4):425-529.
[5]银晓瑞,梁存柱,王立新,等.内蒙古典型草原不同恢复演替阶段植物养分化学计量学[J].植物生态学报,2010,34(1):39-47.
[6]李英年,赵新全,张法伟,等.祁连山冷龙岭南坡移地植物叶片的碳氮特征[J].西北植物学报,2011,31(4):788-794.
[7]郑帷婕,包维楷,辜彬,等.陆生高等植物碳含量及其特点[J].生态学杂志,2007,26(3):307-313.
[8]张国平,刘纪远,张增祥,等.近10年来中国耕地资源的时空变化分析[J].生态学报,2002,22(9):1500-1506.
[9]Ni J.Carbon storage in grasslands of China[J].Journal of Arid Environments,2002,50:205-218.
[10]任海彦,郑淑霞,白永飞.放牧对内蒙古锡林河流域草地群落植物茎叶生物量资源分配的影响[J].植物生态学报,2009,33(6):1065-1974.
[11]冯丽,张景光,张志山,等.腾格里沙漠人工固沙植被中油蒿的生长及生物量分配动态[J].植物生态学报,2009,33(6):1132-1139.
[12]左小安,赵哈林,赵学勇,等.科尔沁沙地退化植被恢复过程中土壤有机碳和全氮的空间异质性[J].环境科学,2009,30(8):2387-2393.
[13]李国栋,张俊华,陈聪,等.气候变化背景下中国陆地生态系统碳储量及碳通量研究进展[J].生态环境学报,2013,22(5):873-878.
[14]林丽,张法伟,李以康,等.高寒矮嵩草草甸退化过程土壤碳氮储量及C/N化学计量学特征[J].中国草地学报,2012,34(3):42-47.
[15]Zhao H L,Zhao X Y,Zhang T H,etal.Desertification processes of sandy rangeland due to over-grazing in semi-arid area,Inner Mongolia,China[J].Journal of Arid Environments,2005,62:309-319.
[16]左小安,赵学勇,赵哈林,等.科尔沁沙质草地群落多样性、生产力与土壤特性的关系[J].环境科学,2007,28(5):945-951.
[17]Wang R Z.Photosynthetic pathways,life forms and reproductive types for forage species along desertification gradient on Hunshandake desert,North China[J].Photosynthetica,2002,40(3):321-329.
[18]王平,盛连喜,燕红,等.植物功能性状与湿地生态系统土壤碳汇功能[J].生态学报,2010,30(24):6990-7000.
[19]Cortez J,Garnier E,Perez-Harguindeguy N,etal.Plant traits,litter quality and decomposition in a Mediterranean old-field succession[J].Plant and soil,2007,296:19-34.
[20]穆少杰,李建龙,杨红飞,等.内蒙古草地生态系统近10年NPP时空变化及其与气候的关系[J].草业学报,2013,22(3):6-15.
[21]陶冶,张元明.准噶尔荒漠6种类短命植物生物量[J].草业学报,2014,23(2):38-48.
[22]李玉霖,毛伟,赵学勇,等.北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究[J]环境科学,2010,31(8):1716-1725.
[23]Li X R,Kong D S,Tan H J,etal.Changes in soil and vegetation following stabilisation of dunes in the southeastern fringe of the Tengger Desert,China[J].Plant and Soil,2007,300(1):221-231.
[24]闫建成,梁存柱,付晓玥,等.草原与荒漠一年生植物性状对降水变化的响应[J].草业学报,2013,22(1):68-76.
[25]Alhamad M N,Alrababah M A.The impacts of biologically induced micro-environments on biodiversity in a dry Mediterranean grassland[J].Plant Ecology & Diversity,2013,6(2):279-288.
[26]彭海英,李小雁,童绍玉.干旱半干旱区草原灌丛化研究进展[J].草业学报,2014,23(2):313-322.
[27]张继义,赵哈林,张铜会,等.科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态[J].植物生态学报,2004,28(1):86-92.
[28]韩梅,杨利民,张永刚,等.中国东北样带羊草群落C3和C4植物功能群生物量及其对环境变化的响应[J].生态学报,2006,26(6):1825-1832.
[29]唐海萍,刘书润,张新时.内蒙古地区的C4植物及其生态地理特性的研究[J].植物学报,1999,41(4):80-84.
