侧柏叶温及叶绿素荧光特性对土壤水分胁迫的响应
2014-01-02董果戴勐赵勇李建中侯娜宋晓明樊巍
董果,戴勐,赵勇†,李建中,侯娜,宋晓明,樊巍
(1.河南农业大学,450002,郑州;2.河南省济源市国有大沟河林场,454650,河南济源;
3.河南省林科院,450003,郑州)
侧柏(Platycladus orientalis)适应性和抗逆性较强,是河南省太行山地区的主要造林树种之一,也是分布面积较大的森林类型。由于特殊的生境条件,水分是影响该区群落分布、森林发育和造林成活率的最关键生态因子。光合作用是植物最基本的生命活动,是植物有机质合成和获取能量的根本来源,是干旱影响作物生长和代谢的第一环节[1]。光合作用是植物体内重要的代谢过程。由于其在植物进化过程中的稳定性而一直成为植物生理生态研究的重要内容,因此可以作为判断植物生长和抗逆性强弱的指标[2]。干旱胁迫下植物的光合能力有不同程度降低[3-4],引起植物水分亏缺,延缓、停止或破坏植物的正常生长[5],植物会产生伤害、修复以及补偿等不同阶段性反应[6]。植物体内发出叶绿素荧光信号的特性与所受到干旱胁迫程度密切相关。叶绿素荧光动力学参数被视为揭示植物光合作用与环境关系的内在探针[7],成为研究作物光合生理与干旱胁迫关系的有力证据[8]。以往对侧柏的研究多侧重于其光合蒸腾特性方面[9-11],多从气孔导度Gs、水分利用效率Wu,E等方面研究其抗旱性,而对其在水分胁迫下叶绿素荧光特性变化方面的详细研究还不多。笔者以太行山侧柏为对象,通过观测不同土壤水分条件下叶温和叶绿素荧光特性的对应变化,研究土壤水分与植物生长间的关系,探讨其抗旱特性及适应水分胁迫的机制,以期为太行山地区造林树种选择和抗旱品种培育提供理论依据。
1 研究地概况
试验地选择在位于黄河小浪底库区北岸的河南省济源市大沟河林场,也是国家林业局小浪底森林生态站。地貌类型为太行山低山丘陵区。中心地理位置为E 112°30'、N 35°03',平均海拔410 m。属暖温带大陆性季风气候。年均气温14.3 ℃,年均日照率54%,年均降水量646.4 mm,年际变化较大。土壤为褐土,pH 值在7.0 ~7.5 之间。
2 材料与方法
2.1 试验材料
供试材料为林场现有盆栽侧柏。选择生长一致且长势良好的20 盆进行试验。将盆栽分为4 个处理,每个处理5 个重复。侧柏的树龄为10 年生,平均树高5 m,平均胸径6.5 cm。盆高50 cm,口径45 cm,土层深度40 cm。
2.2 试验设计
采用环刀法测量田间持水量。按照田间最大持水量的75%~80%(正常供水,对照组CK)、60%~65%(轻度胁迫,LS)、45%~50%(中度胁迫,MS)、20%~25%(重度胁迫,SS)进行控水处理。3 个处理梯度,各5 个重复。处理期间除土壤水分有显著性差异外其他管理一致,处理20 d 后进行试验测定。
2.3 测定指标及方法
2.3.1 土壤含水量 利用时域反射仪(Time Domain Reflectometry,简称为TDR)分别在土层深度10 cm、20 cm 和30 cm 处测量土壤含水量,以做到土壤含水量维持在试验设计的范围之内。同时用土壤烘干法进行校正。
2.3.2 叶温 在08:30—17:30 期间,每1.5 h 循环测量一次。选取2 片完全展开且受光方向一致的植株中部叶片(叶片同光合)进行测量,同时做好标记,以便选择同一叶片测定其叶绿素荧光特征参数。测定仪器为CB-0231 热电偶测温仪(CID 北京生态科学仪器有限公司生产)。
2.3.3 叶绿素荧光 测定仪器为便携式调制叶绿素荧光仪Mini-PAM(德国Walz 公司)。对测量过叶温的同一个叶片,在06:30 时对其进行20 ~30 min暗适应处理后测定初始荧光F0和最大荧光Fm。同样在08:30—17:30 期间,每隔1.5 h 测量各叶绿素荧光参数,分别为即时最大荧光F'm、量子产量、相对光合电子传递速率和光合有效辐射强度。
2.4 数据统计分析
采用Excel2007 和SPSS17.0 软件对试验数据进行分析处理并绘图。
3 结果与分析
3.1 植物叶温对水分胁迫的响应
图1 示出不同水分处理下侧柏叶温日变化趋势都为单峰型曲线。08:30 后随气温升高而上升,峰值出现在13:00,之后随着气温的下降而下降。随着胁迫的增加,叶温逐渐升高,即:正常供水(对照)条件下,在各个时段叶温值均最低;重度胁迫下,叶温值均最高。

图1 侧柏叶温和叶温日均值对水分胁迫的响应Fig.1 Responses of leaf temperature and diurnal average of leaf temperature of Platycladus orientalis to soil water stress
由图1 可知,侧柏叶温日均值随土壤水分胁迫的加剧而逐渐增加。对照组叶温日均值为31.79℃,重度水分胁迫下叶温日均值为32.57 ℃,轻度、中度及重度水分胁迫下分别比对照升高了0.8%、1.7%和2.4%。
叶面温度变化受到如水分、温度、光照、空气湿度等众多因素影响,但在其他条件基本一致的情况下,植物蒸腾是叶温的最主要影响因素[12]。当植物受到水分胁迫时,叶片能通过保卫细胞来调节气孔开闭,从而降低蒸腾强度。相对于供水充足的植株,植物蒸腾的潜热减少,显热增加,叶片温度相应上升;因此,在土壤水分胁迫下的植物叶温要高于土壤水分充足的叶温。
3.2 侧柏叶绿素荧光动力学参数对土壤水分胁迫的响应
3.2.1 F0、Fm和Fv对土壤水分胁迫的响应 F0为初始荧光(固定荧光、0 水平荧光),是光系统Ⅱ(PSⅡ)反应中心处于完全开放时的荧光产量。由图2知,随着水分胁迫程度的增加,F0表现为上升趋势。与对照相比,轻度水分胁迫下F0没有上升,而是小幅下降,降幅为4.9%;中度及重度水分胁迫下,F0均上升。其中重度胁迫下,上升幅度达到18.6%(与对照相比)。F0数值的增大,说明PSⅡ光化学反应中心受到了一定程度的破坏或可逆失活。而轻度胁迫下F0变化不大,说明轻度水分胁迫对侧柏光合作用的影响较小。

图2 侧柏F0、Fm和Fv对土壤水分胁迫的响应Fig.2 Responses of F0,Fm and Fv of Platycladus orientalis leaves to water stress
最大荧光Fm是PSⅡ反映中心完全关闭时的荧光产量,可反映PSⅡ的电子传递情况,它的降低是光抑制的一个特征。与对照相比,Fm在各个水分处理下均下降。轻度水分胁迫下降5.3%,中度胁迫下降12.7%,重度胁迫下降20.0%。这表明水分胁迫使PSⅡ原初光化学活性受到抑制,PSⅡ活性中心受到损伤。
可变荧光Fv是Fm与F0的差值,反映了PSⅡ原初电子受体QA的还原情况,与PSⅡ的原初反应过程有关[13]。从图2 可以看出,随着水分胁迫的加剧,Fv逐渐下降。重度水分胁迫下下降幅度最大,为27.7%(与对照相比),说明侧柏的光化学活性随着水分胁迫的加剧而减弱,且重度胁迫下最弱。
由表1 看出,不同的水分处理间,F0、Fm和Fv的方差分析均表现为显著性差异,表明F0、Fm、Fv对水分胁迫的响应较敏感。
3.2.2 Fv/Fm和Fv/F0对土壤水分胁迫的响应 PSⅡ光化学效率(Fv/Fm)是指PSⅡ最大原初光化学量子效率。不同程度水分胁迫下,Fv/Fm都表现为下降(图3)。与对照相比,轻度水分胁迫下变化较小,中度和重度胁迫下分别下降4.1%和9.7%,下降较为明显。一般植物在正常环境下叶片荧光参数Fv/Fm的值变化极小,不受物种和生长条件的影响[14];但是在逆境条件下或受到伤害时Fv/Fm值可能会明显降低。Fv/Fm下降,表明PSⅡ原初光化学活性受到抑制,PSⅡ活性中心受到损伤。

表1 土壤水分胁迫下F0、Fm和Fv的方差分析Tab.1 Analysis of variance of F0,Fm and Fv at different degrees of water stress
Fv/F0对水分胁迫的响应的变化趋势与Fv/Fm相似(图3)。与对照相比,轻度水分胁迫下同样变化较小,中度和重度胁迫下分别下降20.1%和39.1%,下降更为明显。Fv/F0则反映了PSⅡ的潜在活性,其数值的下降可能是由于F0值上升,Fv值下降。
水分胁迫下,Fv/Fm和Fv/F0均出现下降趋势。这说明在水分胁迫下,PSⅡ光系统的光合电子由PSⅡ反应中心向QA、QB及PQ 库传递过程受到了影响,PSⅡ光系统的原初光能转化效率降低,其潜在活性中心受损。
对侧柏Fv/Fm和Fv/F0进行方差分析(表2),各个水分胁迫下差异显著,而正常供水与轻度胁迫下未达到显著性差异。
3.2.3 实际光化学量子产量日变化 实际光化学量子产量是PSⅡ实际光化学量子效率,指PSⅡ所吸收的光能用于光化学反应的比例。各水分胁迫下侧柏的实际光化学量子产量的日变化规律(图4)均为凹型曲线,且在4 个处理中,各个时段里正常供水(对照)组的值最高,重度水分胁迫下其值最低,其日均值随着水分胁迫程度的加重而降低。在轻度、中度及重度水分胁迫时分别比对照降低了3.6%、8.8%和13.3%。实际光化学量子产量反映了PSⅡ反应中心部分关闭情况下的原初光能捕获效率,表示植物体光合机构将吸收的光能进行转化的能力。其数值的降低说明光系统受到了水分胁迫的伤害,且重度胁迫下最为严重。
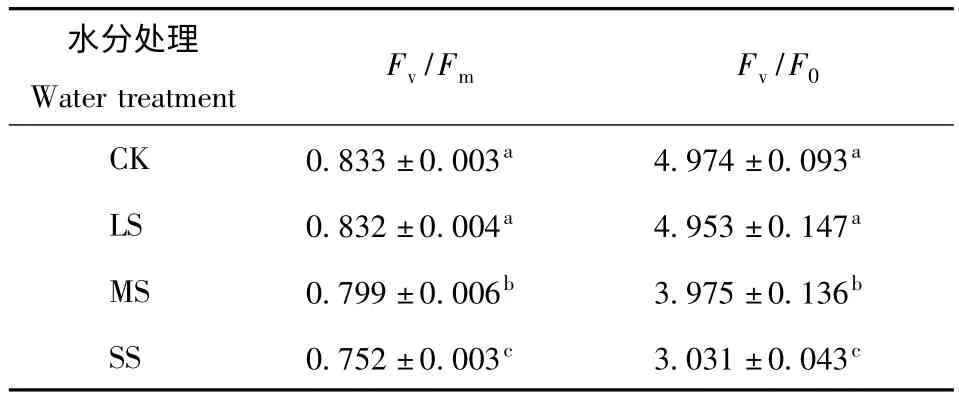
表2 土壤水分胁迫下Fv/Fm和Fv/F0的方差分析Tab.2 Analysis of variance of Fv/Fm and Fv/F0 at different degrees of water stress
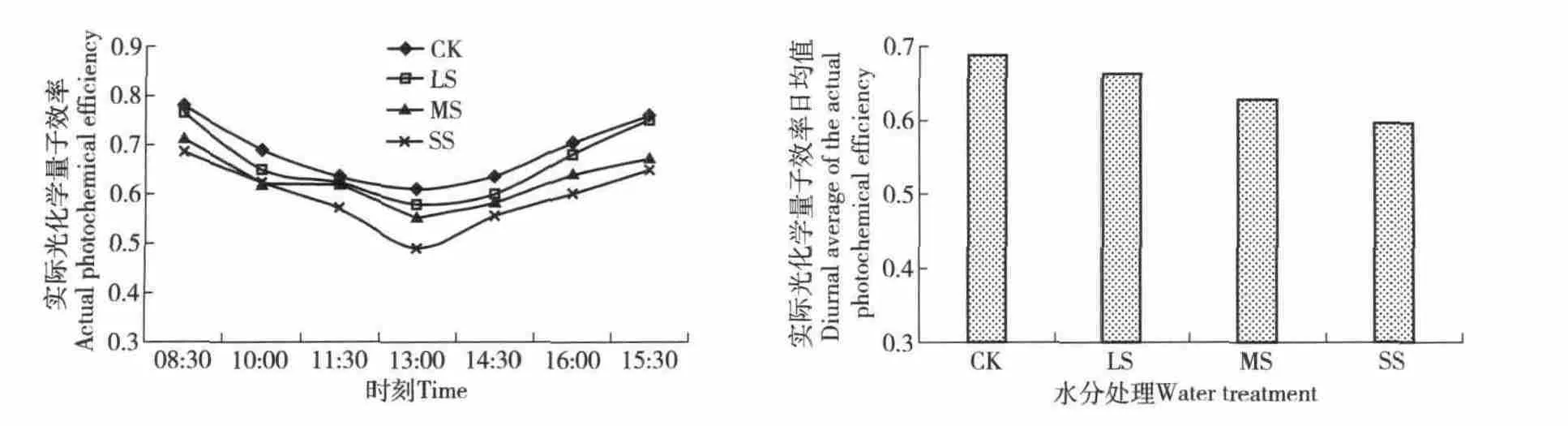
图4 侧柏PSⅡ实际光化学量子效率的日变化与其日平均值Fig.4 Daily variation of the actual efficiency and diurnal average of the actual efficiency of Platycladus orientalis
3.2.4 表观光合电子传递速率日变化 表观光合电子传递速率,是光合机构吸收光能发生电荷分离产生电子并沿电子传递链向下传递的速率。水分胁迫下,表观光合电子传递速率的日变化为单峰凸型曲线(图5),在13:00 左右达到最大值。在4 个处理中,各个时段里正常供水(对照)组的值最高,重度水分胁迫下其值最低。这一现象也可以从表观光合电子传递速率的日均值中得以验证。表观光合电子传递速率的日均值(图5)变化规律也很明显,从正常供水到重度胁迫依次降低。与对照相比,重度水分胁迫下其日均值下降13.8%。以上趋势及数据表明,水分胁迫抑制了PSⅡ光系统电子传递。
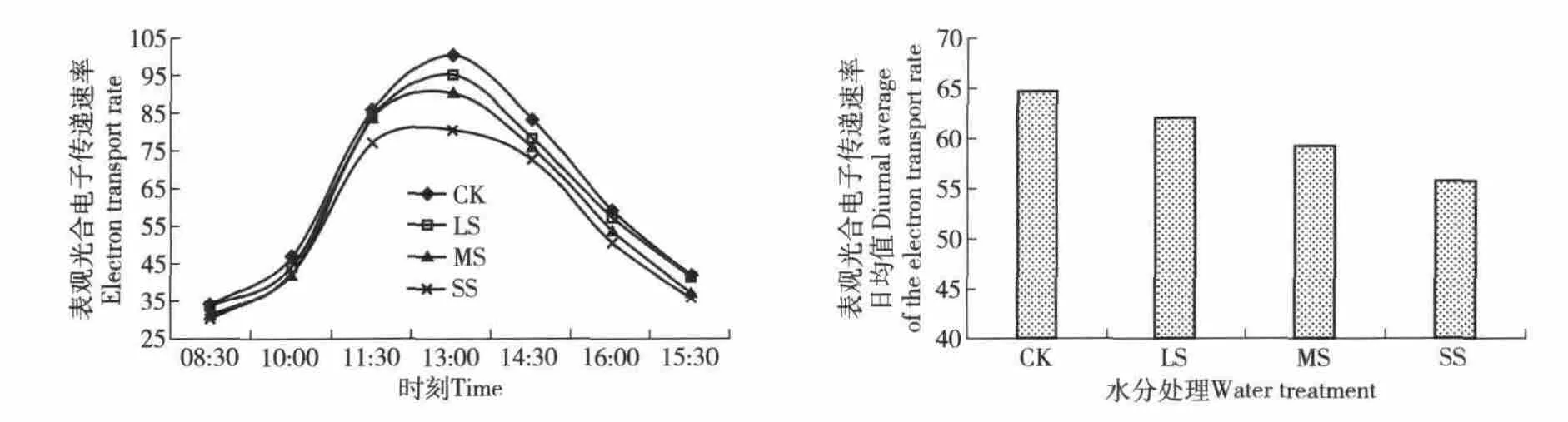
图5 侧柏表观光合电子传递速率的日变化与其日平均值Fig.5 Daily variation of the electron transport rate and diurnal average of the electron transport rate of Platycladus orientalis
3.2.5 非光化学淬灭日变化 非光化学淬灭反映的是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分。不同水分胁迫下非光化学淬灭的日变化为单峰凸型曲线,在13:00 左右达到最大。与表观光合电子传递速率的日变化不同的是,在4 个水分处理中,各个时段中正常供水(对照)下值最低,重度水分胁迫下最高。其日均值是随着水分胁迫程度的增加而升高,重度水分胁迫下达到0.807。非光化学淬灭随水分胁迫的加剧而增加,说明PSⅡ天线色素吸收的光能以热的形式耗散掉的光能增加。这种耗散缓解了逆境条件对植物的不利影响,是侧柏对水分胁迫或者午间高温的调节机制,保护了其光合机构。

图6 侧柏非光化学淬灭的日变化与其日平均值Fig.6 Daily variation of the non-photochemical quenching coefficient and diurnal average of the non-photochemical quenching coefficient of Platycladus orientalis
4 结论与讨论
1)侧柏的叶温、表观光合电子传递速率和非光化学淬灭参数的日变化变现为单峰凸型曲线。实际光合电子传递速率的日变化为单峰型凹函数。都在13:00 左右达到峰值。
2)随着水分胁迫的加剧,侧柏的叶温和非光化学淬灭参数日均值逐渐升高,而Fm、Fv、Fv/Fm和Fv/F0以及实际光合电子传递速率、表观光合电子传递速率参数的日均值均逐渐下降,而F0表现为逐渐上升的趋势,即在轻度水分胁迫下略微降低,而在中度及重度水分胁迫下逐渐升高。
土壤水分直接影响植物水分。其他条件一致时,土壤水分充足,叶温则低;土壤水分亏缺,叶温则高[15]。本研究结果与此一致,土壤含水量越小,叶温越高(图1)。在其他因素一致的情况下,叶温可以作为反映侧柏土壤水分状况的指标之一。
水分胁迫对植物光合作用的影响有很多方面,不仅直接引发光合机构异常,而且影响光合电子传递[16]。本实验结果表明,干旱胁迫导致F0出现上升的趋势,Fm、Fv、Fv/Fm和Fv/F0值下降。许多研究[17-18]都表明,水分胁迫会导致F0上升和Fv下降。抗旱性越强的品种,干旱胁迫对其反应中心尤其是光系统的破坏也越小,PSⅡ光化学活性也越大;因此F0和Fm可作为植物能否抵抗干旱的指标。水分胁迫会导致最大原初光化学量子效率降低,PSⅡ的潜在活性中心受损,表明轻度和中度胁迫下光系统未受到或受到较轻的胁迫,重度干旱胁迫下,叶片光合器官受到不可逆伤害,光合细胞活性降低,光合作用受到严重抑制。这与文献[19-20]的研究结果一致。
随着干旱胁迫的加重,侧柏的表观光合电子传递速率显著下降,水分胁迫使侧柏PSⅡ反应中心的电子传递效率下降,并且重度水分胁迫下的下降最为显著。在干旱胁迫下导致植物PSⅡ反应中心的电子捕获效率降低,从而降低电子传递速率[21]。
在水分胁迫的研究中,对叶绿素荧光淬灭的非光化学淬灭的变化探讨较多。本研究中,侧柏的非光化学淬灭值随水分胁迫的加剧而上升,与已有研究报道[22-24]一致。干旱胁迫造成光抑制时,植物以增加非光化学猝灭的来适应逆境[25]。多余的光能以热的形式耗散掉,防止光合器官的破坏,对光合器官起一定的保护作用,是植物的一种自我保护机制[22]。从另一个方面来说,非光化学淬灭明显升高显著降低了光能利用效率,多余的光能无法用于光合电子传递及热耗散,造成光能在光系统内部的积累,引起光系统结构的破坏和光反应中心活性的降低,进一步影响植物的光合作用[23];因此,这种适应只是一种内耗损的被动适应方式,可以维持生存,但对生长不利。
本研究中,侧柏在经受了20 d 的水分胁迫下,通过增加热耗散(非光化学淬灭升高)来保护光合机构,可见侧柏叶片启动了抗旱机制,其光合机构并未受到严重的损伤,依然具有一定的活性,说明侧柏在抗旱能力和适应性方面表现较好,具有一定的适应调节能力,对水分胁迫的适应较为积极主动;然而,参阅文献后发现,对干旱胁迫后侧柏恢复能力强弱[26]以及在干旱胁迫中生理指标如抗氧化剂(过氧化氢酶、过氧化物酶和超氧化物歧化酶等)、脯氨酸和可溶性糖[27]的变化还可以进一步的探讨,以获得更为详尽的理论依据。
