霍山七里峪次生落叶阔叶林边缘效应研究
2013-12-29翟静娟杨志芳毕润成
翟静娟,杨志芳,毕润成
(1.朔州职业技术学院,山西 朔州 036002;2.山西师范大学生命科学学院,山西 临汾041004)
霍山七里峪次生落叶阔叶林边缘效应研究
翟静娟1,2,杨志芳2,毕润成2
(1.朔州职业技术学院,山西 朔州 036002;2.山西师范大学生命科学学院,山西 临汾041004)
以霍山七里峪随机选择的落叶阔叶林、油松针叶林边缘为研究对象进行对比分析。主要用样方法对4个落叶阔叶林和2个油松针叶林的边缘进行了调查,总共设置了192个5 m×5 m落叶阔叶林样方和72个5×5 h绿素含量进行测定发现:各物种叶绿素含量沿边缘表现出边缘-10~10 m区域>边缘10 m内>边缘-10 m外区域。
边缘效应;落叶阔叶林;针叶林;生物多样性;叶面积指数
由于人口的增长、经济持续快速的发展等原因,人类对自然资源的需求日益增大,连续的原始森林多被破坏,大量的次生林由此形成,并且山现了由农田、草场和荒地等退化生态系统包围的森林片断,形成了与林区具鲜明对比的大量林缘景观。林缘是能量和物质(或物种)水平交换最强的地带[1],由其所产生的生物和环境边缘效应不仅影响林缘物种组成、分布,而且导致森林生境改变,进而影响森林生态系统结构与功能[1-3]。因此,边缘效应的研究受到了各领域(生物、生态、大气科学等)学者的广泛关注,并成为当前森林生态学研究的一个热点。1933年,野生动物学家Leopold[4]提出了边缘效应(edge effect)的概念,并对森林片断的边缘效应进行了开创性的研究[5]。本研究将从不同的森林群落类型入手,探索边缘效应对暖温带地区不同群落类型的物种多样性、物种种类、生物量等的影响,对小尺度边缘效应产生的机理进行研究探索。
1 样地环境与研究方法
1.1 样地自然环境
霍山位于山西省南部,临汾盆地的东北边缘, 介 于 北 纬 36º21'~ 36º45', 东 经 115º40'~112º20'[2]。本区属暖温带大陆性季风气候,年平均气温在9.3~12.3ºC之间[2]。无霜期可达120~200 d,年降水量在500~700 mm之间,春末易发生干热风[2]。土壤以褐土为主,随海拔升高依次为石灰性褐土、褐土性土、淋溶褐土、棕色森林土、山地草甸土等[3]。
1.2 研究方法
1.2.1 样地基本概况
山西霍山七里峪用样方法进行调查,调查样地共有6个。其中落叶阔叶林样地4个,针叶林样地2个。1~4号样地是典型的落叶阔叶林。1990年左右被人工采伐,由于国家天然林自然保护政策的实施,使得现在的部分森林得以保存。5~6号样地为油松针叶林。其为人工种植的油松林,1980年左右被人工采伐,大部分的油松林得到保存。如上所述,致使大量森林边缘的出现。各样地基本情况如表1示:

表1 霍山样地基本情况Table 1 Condition of study samples of different forest of Huoshan
1.2.2 群落调查方法
在霍山七里峪选取边缘明显的落叶阔叶林与针叶林,对每一个样地进行如下的测量研究:
每个样地样方设置如图1、图2所示,每个样方大小为5 m×5 m,具体进行以下测量:乔木的胸径、高度、冠幅;灌木的高度、冠幅、株丛数、盖度;选择1 m×1 m的草本样方测量草本的种类、株丛数、平均高度、盖度,以及光照指数、叶绿素含量等。

图1 落叶阔叶林样方调查及样方命名示意图Fig. 1 The arrangement of plot in latifoliate

图2 针叶林样方调查及样方命名示意图Fig.2 The arrangement of plot in taiga
1.2.3 数据分析方法
运用SPSS13.0,Excel,BΙO-DAP等软件进行数据分析。
2 结果与分析
2.1 边缘效应与物种多样性
由图 3可以看出,落叶阔叶林灌木层多样性指数(H’),在边缘±5 m区域达到最大值2.483(H’)。随着距离边缘距离的变大,多样性指数(H’)逐渐变小,直至边缘±10 m以外或以内区域趋于稳定。边缘外-10 m左右区域物种多样性指数一般维持在2.200(H’)左右。边缘内+10 m左右区域物种多样性指数一般维持在2.100(H’)左右。边缘外-10 m左右区域物种多样性较边缘内+10 m左右区域大。落叶阔叶林草本层多样性指数(H’)与灌木层多样性指数(H’)表现出相同的变化趋势。在边缘±5 m区域达到最大值2.815(H’)。边缘外-10 m左右区域物种多样性指数一般维持在2.300(H’)左右。边缘内+10 m左右区域物种多样性指数一般维持在2.400(H’)左右。边缘内+10 m左右区域物种多样性较边缘外-10 m左右区域大。但是总体上,草本层物种多样性指数要大于灌木层多样性指数。

图3 落叶阔叶林边缘不同区域 H’Figs. 3 The species diversity(H’) about braod-leaved forest
2.2 边缘效应与植物生长状况
2.2.1 边缘效应与植物光合作用日变化
边缘外区域,榛子光合速率日变化均呈现明显的双峰曲线,而边缘区域、边缘内区域没有出现明显的双峰曲线(如图4)。边缘外区域, 11:00左右达到第 1 个峰值 Pn = 10.01 μmol·m-2s-1,随后出现低谷,即“午休现象”,在13:00~15:00间出现第 2 个峰值 Pn = 11.12 μmol·m-2s-1,15:00以后光合速率迅速下降。边缘区域与边缘内区域,植株从早上开始。光合速率随光合有效辐射的加强逐渐升高,在13:00达到最大值Pn(边缘-10~ 10 m 区域)= 10.51 μmol·m-2s-1和 Pn(边缘 10 m内区域)= 5.91 μmol·m-2s-1,随后光合速率逐渐下降。光合有效辐射是瞬时变化最大的生态因子,相关分析表明,三个区域光合有效辐射与光合速率呈极显著正相关(r=0.933,P=0.05)。这说明,光合有效辐射是影响榛子光合速率的的主要生态因子之一(图5)。3个区域温度的日变化与PAR的日变化趋势基本相同,呈单峰曲线。且在同一时间也表现出,边缘外>边缘>边缘内。边缘外区域叶温变化范围在25~35℃;边缘区域叶温变化范围在25~30℃;边缘内区域叶温变化范围在20~25℃(图6)。由图7,空气相对湿度的日变化曲线与光合日进程曲线不同。在边缘的不同区域,早晨空气相对湿度(RH)最大,然后逐渐降低,在12:00~16:00这段时间RH非常低,之后逐步回升。相关性表明,各区域相对湿度与净光合速率存在负相关关系。如图8,不同区域内的榛子气孔导度与光合有效辐射呈显著正相关,说明光照强度是影响气孔导度变化的主要环境因子。如图9,胞间CO2浓度日变化表明,随着时间的进程,各个区域榛子胞间CO2浓度逐渐下降,到15:00左右逐渐上升。中午12:00~13:00左右,各区域胞间CO2浓度,表现为:边缘内>边缘外区域>边缘区域。相关分析表明,榛子植株光合速率与胞间CO2浓度呈极显著负相关。

图4 不同区域榛子光合速率日动态变化Fig. 4 The daily dynamic of photosynthesis of Corylus eterophylla Fisch.ex Bess.in different zone
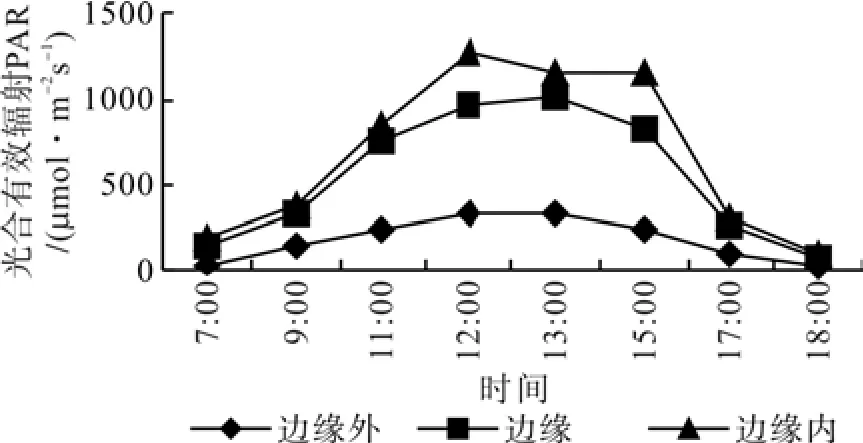
图5 不同区域光合有效辐射日动态变化Fig. 5 The daily dynamic of PAR in different zone

图6 不同区域叶片温度日动态变化Fig. 6 The daily dynamic of Tl in different zone

图7 不同区域相对空气湿度日动态变化Fig. 7 The daily dynamic of RH in different zone
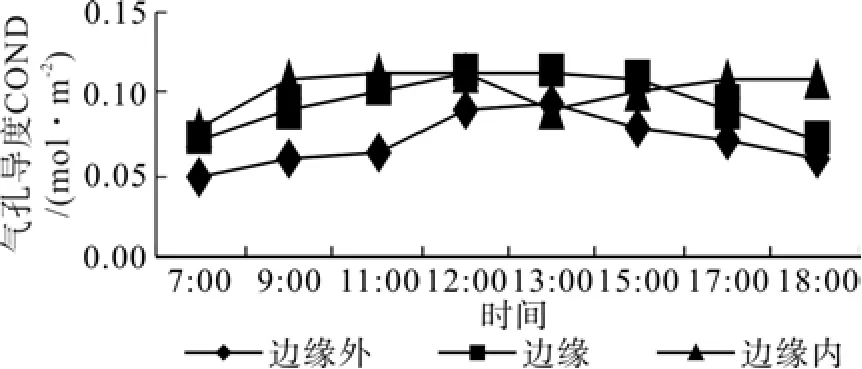
图8 不同区域气孔导度日动态变化Fig. 8 The daily dynamic of COND in different zone

图9 不同区域胞间 CO2 浓度日变化Fig. 9 The daily dynamic of Ci in different area
2.2.2 边缘效应与植物叶绿素含量
本研究对1号样地中,分布比较广泛的物种的叶绿素含量进行测定。每个样方中的每种所选定的植物,选择10片叶片,测定其叶绿素含量。由图10可知,各物种叶绿素含量沿边缘表现出边缘-10~10 m区域>边缘10 m内>边缘-10 m外区域。有学者研究得出:同一种植物的阴生叶叶绿素含量大于阳生叶[11]。边缘10 m内叶绿素含量大于边缘-10 m外区域是由于光照强度的不同引起的。
以下将对这几种植物的叶绿素含量的沿边缘的变化趋势进行分析:
2.2.3 边缘效应与植物叶面积指数 (LAΙ)
叶面积指数(LAΙ),即每单位陆地面积上的叶面积[12]。LAΙ控制着植被许多生理和物理过程,如光合、呼吸和蒸腾作用以及碳循环和降水截留等[13-16],是森林生态系统的一个重要结构参数。本研究利用灌层分析仪,在各落叶阔叶林和针叶林分林外、林缘、林内三个区域测量其叶面积指数,每个区域分乔木层下、灌木层下、草本层下三层测量。从表2可知,各个样地中各层叶面积指数从林外到林内呈上升趋势。且从乔木层下、灌木层下、草本层下依次也呈上升趋势。

图10 几种植物叶绿素含量随边缘变化Fig. 10 Eff i cience of Chloropy Ⅱ about several plant in each plot

表2 各样地各层LAΙTable 2 The LAI of each layer in each sample
3 结论与讨论
由于人为干扰而导致的森林片断化发生后,局部气候和土壤环境发生变化,从而导致生态系统功能的改变。本研究对边缘形成10年左右的落叶阔叶林、油松针叶林进行植被、环境方面的调查研究。
(1) 落叶阔叶林距离边缘不同区域的物种数量,在边缘±5 m左右达到最高;针叶林不同区域的物种数量,从边缘外到边缘内呈下降趋势。其乔木、灌木物种数量在距离边缘±5左右达到最大。
(2) 落叶阔叶林灌木层多样性指数(H’),在边缘±5 m区域达到最大值2.483(H’)。落叶阔叶林草本层多样性指数(H’)与灌木层多样性指数(H’)表现出相同的变化趋势。针叶林草本层多样性指数(H’)与灌木层多样性指数(H’)表现出不同的变化趋势。草本层多样性指数,在边缘-5 m左右以外区域较其他区域大。其物种多样性指数维持在相对稳定的数值(2.60~2.70)。
(3) 边缘外-10 m区域,榛子光合速率日变化均呈现明显的双峰曲线,而边缘-10~10 m区域、边缘10 m内区域没有出现明显的双峰曲线。各物种叶绿素含量沿边缘表现出边缘-10~10 m区域>边缘10 m内>边缘-10 m外区域。各个样地中各层叶面积指数从林外到林内呈上升趋势。且从乔木层下、灌木层下、草本层下依次也呈上升趋势。
本研究对人为干扰而导致的森林片段化的边缘区域进行群落学、环境因子、植物光合生长状况等进行调查分析后对边缘效应的作用机理做出如下推断:
在由于人为干扰而导致的森林片段化发生后,造成局部环境因子的改变,最初变化最大的是地上环境,这些环境因子对地上植物生长起着决定性作用。边缘不同区域光合有效辐射的不同,不同的植物对光合有效辐射的强度要求不同,从而使不同区域植物种类发生改变。另外,不同区域光合有效辐射等环境因子的不同,也使同种植物叶面积指数、叶绿素含量在不同区域不同。因此,气候-植被-土壤是一个有机统一的整体,“气候-植被-土壤”互相作用导致边缘区域特殊景观的出现。
[1]Lee R. Forest Microclimatology[M]. New York∶ Columbia University Press, 1978.
[2]Forman, R T T. Ιnteraction among landscape elements∶ a core of landscape ecology. Ιn∶ S. P. Tjallingii and A. A. de Veer Ιnternational Congress.Netherlands Society of landscapes Ecology[M]. 1981.
[3] 刘宏茂,许再富,陶国达.西双版纳傣族“龙山”的生态学意义 [J]. 生态学杂志,1992, 11(2)∶ 41-43.
[4]Leopold A.Game management[M]. New York∶ Charles Scribner's Sons, 1933.
[5]马友鑫, 刘玉洪, 张克映. 西双版纳热带雨林片断小气候边缘效应的初步研究 [J]. 植物生态学报 , 1988, 22(3)∶ 250-255.
[6]Anderson J M. Ecology of environmental science-resources and environmental science series[J]. Edward Arnold (Ltd.),198l∶ 87-95.
[7]Holland M M. SCOPE/MAB technical consultations on landscape boundaries∶ report on a ACOPE/MAB workshop on ecotones[J]. Biology Ιnternational(Speciallssue),1988,(17)∶47-106.
[8]Castri F D, Hansen A J. The environment and development crises as determinants of landscape dynamics[A].Ιn∶ Hansen A J,Castris F D(eds.). Landscape Boundaries[C]. New York∶ Springer-Verlag, 1992∶ 3-18.
[9]毕润成, 杨焕根, 朱新军. 山西霍山落叶阔叶林边缘效应的研究 [J]. 西北植物学报 , 2004, 24(8)∶ 1441-1447.
[10] 许大全 , 徐宝基 , 沈允钢 . 3 植物光合效率的日变化 [J]. 植物生理学报 , 1990, 16(1)∶ 1-5.
[11] 薛云申. 光照强度对叶片叶绿素含量的影响[J].生物学教学, 2001, 26(10)∶ 24.
[12] 周宇宇 , 唐世浩 , 朱启疆 , 等 . 长白山自然保护区叶面积指数测量及结果 [J]. 资源科学 , 2003, 25(6)∶ 38-42.
[13]颜文洪, 胡玉佳. 海南石梅湾青皮林LAΙ的冠层数字成像间接法测算 [J]. 中山大学学报 , 2004, 43(3)∶ 71-74.
[14] 徐全芝 , 张万昌 , 刘三超 , 等 . 黑河流域叶面积指数的遥感反演 [J]. 干旱区研究 , 2003, 59(1)∶ 27-31.
[15] 张金屯 . 数量生态学 [M]. 北京 ∶ 科学出版社 , 2004.
[16] 闫 明,钟章成,乔秀红. 缙云山片断长绿阔叶林小气候边缘效应的初步研究 [J]. 应用生态学报 , 2006,17(1)∶ 17-21.
[17]马文俊 , 李际平 , 李建军 , 等 . 杉—阔林间边缘效应与杉木林龄的相关性研究[J]. 中南林业科技大学学报,2011,31(7)∶75-79.
[18] 王国华, 李际平, 赵春燕. 基于层次分析法的森林景观边缘效应强度分析 [J]. 中南林业科技大学学报 , 2012,32(4)∶110-116.
Edge-effect about secondary Latifoliate in Huoshan Mountain
ZHAΙ Jing-juan1,2, YANG Zhi-fang2, BΙ Run-cheng2
(1. Shuozhou Vocational Technology College, Shuozhou 036002, Shanxi, China;2. School of life science, Shanxi Normal University, Linfen 041004, Shanxi, China)
The latifoliate and taiga was searched. The plot was setted upright the forest edge. The plant community survey, environment factors and the daily dynamic of photosynthesis of Corylus eterophylla Fisch. ex Bess. was mensurated. The result showed that∶The grass layer is as the same as the shrub layer. Ιn latifoliate, the number of species, the species diversity of shrub, the species diversity of grass have the same change trend. The Chlorophy Ⅱ mensuration about several plant that distribute abroad show that∶ The Chlorophy Ⅱ content of the plant is different depend on the distance from the edge. The Chlorophy Ⅱ content of the plant in the district that the distance from the edge shorter than 10 m is much than the distance from the edge longer than 10m in forest . The Chlorophy Ⅱ content of the plant in the district that the distance from the edge longer than 10m in forest is much than the distance from the edge longer than 10 m out forest .
edge effect; latifoliate; taiga; species diversity; LAΙ
S718.5
A
1673-923X(2013)12-0057-05
2013-08-22
山西省化学优势重点学科建设项目(912019)资助;山西省留学基金项目(20081073)资助
翟静娟(1983-),女,山西临汾人,硕士研究生,主要从事种子生理生态方面的研究;E-mail: zhaijingjuan@163.com
毕润成(1955-),男,山西寿阳人,教授,硕士生导师,山西师范大学生命科学学院,主要从事植物生态学研究;
E-mail: rcbi@dns.sxtu.edu.cn
[本文编校:吴 彬 ]
