江西大岗山杉木人工林生态系统碳汇功能研究
2013-12-28潘勇军陈步峰彭清珍
潘勇军,王 兵,陈步峰,彭清珍
(1.中国林业科学研究院 热带林业研究所,广东 广州 510520; 2.中国林业科学研究院 森林与环境保护研究所,北京 100091;3.广州市第四十七中学汇景实验学校,广东 广州 510060)
江西大岗山杉木人工林生态系统碳汇功能研究
潘勇军1,王 兵2,陈步峰1,彭清珍3
(1.中国林业科学研究院 热带林业研究所,广东 广州 510520; 2.中国林业科学研究院 森林与环境保护研究所,北京 100091;3.广州市第四十七中学汇景实验学校,广东 广州 510060)
研究森林生态系统碳储量以及碳汇功能是一项基础工作,对评价森林生态系统在碳平衡中的作用有着重要意义。对江西大岗山16年生杉木人工林生态系统碳储量和碳平衡进行研究,估算其碳汇/源功能。结果表明:杉木人工林生态系统总生物量为156.256 t·hm-2,乔木层占94.9%,林下层生物量为3.197 t·hm-2,年凋落物归还量为4.756 t·hm-2;杉木人工林碳储量为77.246 t·hm-2,其平均年净生产力为6.837 t·hm-2,其碳汇能力为1.595(±1.179) t·hm-2a-1,可见杉木人工林林生态系统显著地固定了大气中的CO2,有着明显的碳汇作用。
杉木人工林;生物量:碳储量;森林碳汇
森林碳汇是指森林通过光合作用吸收大气中的二氧化碳并将其固定在植被或土壤中,从而减少二氧化碳在大气中的浓度,在全球大气碳平衡中的作用极为显著[1-2]。森林生态系统维持着约占全球植被碳库80%以上的碳库[3-4],森林碳汇已成为国际陆地系统碳循环研究的热点。目前国内外有关国家、区域及生态系统水平上的森林碳平衡的研究较多[5-8]。我国在森林碳平衡研究方面也进行了深入的研究,方精云等[1]对我国近50 a来森林碳库,CO2源汇功能变化进行了研究,方晰等[9]对会同森林定位站第二代杉木人工林速生阶段的碳密度、碳储量及其空间分布特征和碳平衡进行了研究;方精云等[10]对北京东灵山3种温带森林生态系统碳储量、碳通量和碳平衡进行了研究;部分研究人员基于野外的实测数据,分别研究了中国东部热带雨林、亚热带常绿阔叶林和温带落叶阔叶林、温带针叶林等森林类型的碳储量和生产力,为中国森林生态系统碳循环的研究提供了重要的基础数据[11-14]。
我国人工林面积占世界人工林面积的26%,年均增长面积大,人工造林对森林碳汇具有重要贡献[15],建立一些代表性的人工林生态系统碳循环模式研究基准点,进行生态系统碳循环的评价和预测,准确估算我国人工林的年碳汇量,对我国应对《京都议定书》第二承诺期谈判和CDM碳交易,具有重要的现实意义。杉木人工林Cunninghamia lanceolata是中国南方集体林区的造林的主要森林类型,占人工林面积的30%[16],在区域林业碳汇项目中起到重要作用,其碳汇功能的研究将为进一步深入研究我国乃至全球范围内的森林生态系统碳循环提供正确的基础数据,为正确估算我国乃至全球森林生态效益提供参考。目前对杉木生态系统碳循环过程的研究较多[17-20]。但是,对整个杉木林生态系统各个碳库之间的碳汇/源研究相对较少。本研究研究了16年生杉木人工林植被生物量,利用树木年轮分析其生物年生产力,结合土壤呼吸量研究,系统估算杉木人工林生态系统碳汇功能,为全国森林生态系统碳汇计量提供基础数据。
1 研究地概况
试验区设置在江西大岗山国家级森林生态站,江西省分宜县中国林科院亚热带林业实验中心的年珠林场(114°30′~ 114°45′ E,27°30′~ 27°50′N)。气候温暖湿润,属中亚热带季风湿润类型,年平均温度为15.8℃~17.7 ℃,7月平均最高温度28.8 ℃,最高温度39.9 ℃,1月平均最低温度为-5.3 ℃,日最低温度为-8.3 ℃。多年平均降水量为1 591 mm,年均无霜期为268 d,生长季较长,土壤属低山丘陵红壤、黄壤类型及其亚类[21],主要植被类型有天然次生常绿阔叶林、落叶阔叶林、各类针阔混交林、毛竹林及大面积杉木林[22]。
研究区域样地为1994年营造的杉木纯林,平均密度为1 269 株·hm-2,平均胸径为17.96 cm,平均树高为15.36 m,林分胸径各径级分布呈正态分布,结构合理,符合人工同龄林胸径结构的一般规律。该林分处于生长旺盛期。林下植被较少,主要有狗脊Woodw ardia japonica (L. f) Sm.、毛药红淡Adinandra millettii (Hook. et Arn. ) Benth.、機木Loropetalum chinense (R. Br) Oliv.、悬钩子Rubus trianthus Focke.、 飞 蓬 Erigeron acris L.、大青Clerodendrum cyrtophyllum Turcz.、杜茎山Μaesa japonica (Thunb) Moritz.i 、连蕊茶Camelllia fraternal Hance.、 油 茶 Camellia oleifera Abe1.、土茯苓Smilax glabra Roxb.、木荷Schima superb Garde. et Champ. 等。设置了10 m×10 m的固定样地62个,进行每木调查(胸径和树高),采伐的标准木在样地外附近采伐,立地条件相差不大。
2 研究方法
2.1 标准木生物量测定方法
依据样地调查资料,分8个不同径级(8~11 cm、11~13 cm,…,23~25 cm),每个径级选取2株标准木,共16株。伐倒解析木,把带枝叶的树干,从地表0 m,地上1.3 m、3.3 m、5.3 m及以上每隔2 m锯开,逐层分干、枝、叶单独称质量,取混合样称鲜质量带回烘干。截取2~5 cm的圆盘,称质量,标出记号。
根系生物量及林下植物生物量测定:从树干位置开始向四周清除表土,逐渐向下挖掘,使根系全部露出,分粗、中、细分别称质量,即为全株根量。同时设1 m×1 m小样方进行林下木草本层生物量测定,将所有地上灌木和草本全部连根挖出,称量茎叶及根系的鲜质量,并取样带回80℃烘干至恒质量。
年凋落物归还量测定:在样地内设置10个1 m×1 m凋落物收集框,每月按时收集框中凋落物,称质量后取样烘干称干质量。
2.2 生物量相对生长模型
用实测不同径级标准木的生物量和胸径、树高数据建立各器官生物量与胸径、树高的相对生长模型。
W=a(D2H)b;
W=aDb。
其中W为生物量,D为胸径(单位:cm),H为树高(单位:m),a、b为待定参数。
2.3 年轮宽度测量与交叉定年
用Lintab5年轮分析仪进行年轮宽度测量,用Coffcha(version 6.06P)程序进行交叉确定年数[22]。
2.4 森林碳汇量估算方法
乔木层生产力测定:林木胸径与生物量之间存在明显的相关性,建立了相对生长模型,估算林分生物量。对标准木树木圆盘年轮宽度的测量,根据年轮宽度和相对生长模型推算标准木的年生物量增量,估算其年生产力。
凋落物归还量即为一年每月收集的总凋落物生物量;林下植被年净生物产量以其平均年龄除之(灌木6 a,草本3 a),所有生物量均按样品测定的含碳率折算为含碳量。
生态系统碳平衡计算:生态系统净初级生产力(NPP)为生物量年净增量和凋落物归还量加和所得;净生态系统生产力(NEP)为净初级生产力(NPP)再减去异养生物(土壤)的呼吸作用所消耗的光合作用产物之后的净差值,表示CO2进入生态系统的净光合产量[23-24]。在大尺度上可以应用NEP来评价生态系统的对CO2的源/汇状况,当NEP>0时,表明生态系统是大气CO2的汇;当NEP<0时,则表明生态系统是CO2的源[10]。
3 结果与分析
3.1 杉木标准样木相对生长模型
根据采伐的16株标准木干、枝、叶,根等各组分生物量的实测值,建立胸径(D)、树高(H)与各器官生物量及总生物量的相对生长模型(见表1),杉木单木总生物量、地上部分生物量及各器官生物量与树高和胸径之间均存在密切的相关关系。图1表示实测标准木生物量与利用相对生长模型估算生物量拟合度很好,两组数据差异不显著(F=2.041)。
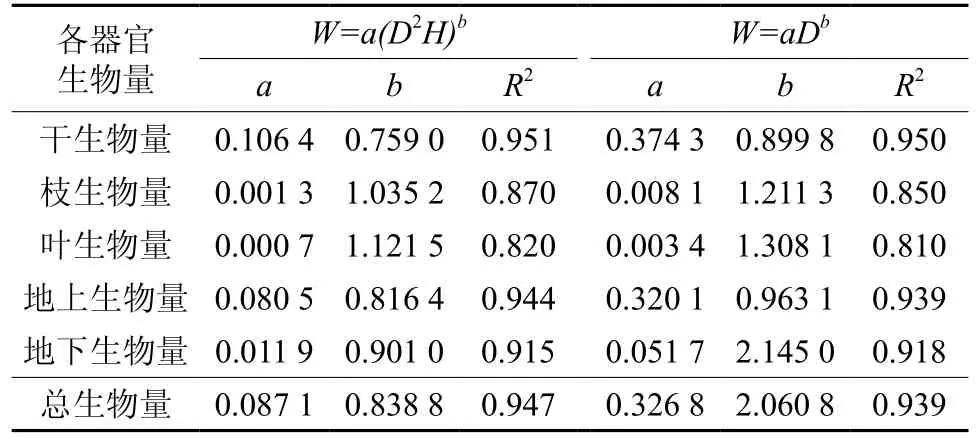
表 1 杉木林各器官生物量相对生长模型Table 1 DBH-weight allometry model parameters in different organs of Chinese fir
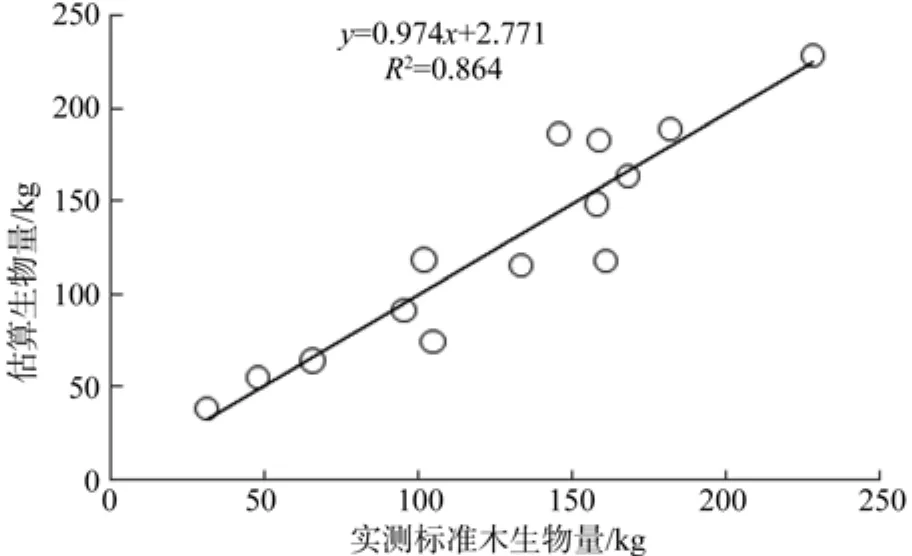
图 1 杉木标准木实测生物量与估算生物量拟合Fig. 1 Simulated and measured biomass fi tting of sample tree Chinese fi r
3.2 杉木林分生物量
利用相对生长关系建立回归估测方程,来推算杉木林生态系统单位面积的生物量,估算整个生态系统的初级生产量和生产力。估算江西大岗山16年生杉木人工林生态系统总生物量为156.256 t·hm-2,乔木层的总生物量为143.303 t·hm-2,占总生物量的94.9%。其中地上部分生物量为113.66 t·hm-2,占整个乔木层生物量的76.64%。杉木林内下层发育较差且分布不均,灌木较少,多为草本层,盖度仅为11%左右,林下层生物量为3.197 t·hm-2。2009年年凋落物归还量为4.756 t·hm-2a-1,是地被层碳库和土壤碳库的主要来源。
杉木乔木层各器官生物量分配存在一定差异(见图2),树干生物量为92 t·hm-2,占乔木层总生物量62.04%,树干是地上生物量的主要部分。其次是根系生物量,占总生物量的23.36%,为34.65 t·hm-2,枝和叶的生物量各占8.33%和6.28%。
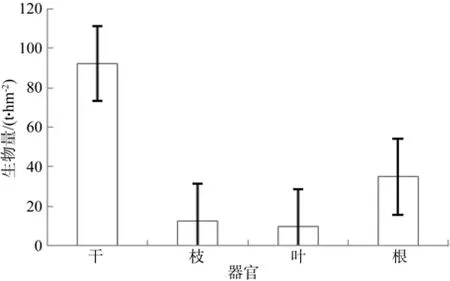
图 2 杉木乔木层各器官生物量及分配Fig.2 Biomass and its distribution in tree story of Chinese fi r
杉木林乔木层地下根系生物量所占比例较大,地上、地下生物量分配比为3.28,根系部分生物量不能忽略不计。根系生物量的野外调查工作量比较大,因此对与根系生物量的计算,可以依据地上部分生物量和地下部分生物量呈一定关系进行换算,减轻野外工作量。进行相关分析发现:杉木乔木层地上部分生物量与地下部分生物量呈乘幂关系,拟合模型为:
y=0.221x1.071,(R2=0.885)。
3.3 杉木林生态系统乔木层年碳储量的估算
图3为16年生杉木林乔木层总碳储量和年碳固定量情况,总碳储量随林龄的变化,呈增长趋势,用一元线性回归拟合,相关性很高。16 a乔木层总碳储量达到73.905 t·hm-2,随着年龄的增长,碳储量还会出现更多的积累,达到一定的年龄后会出现拐点,增长速率放慢[4]。
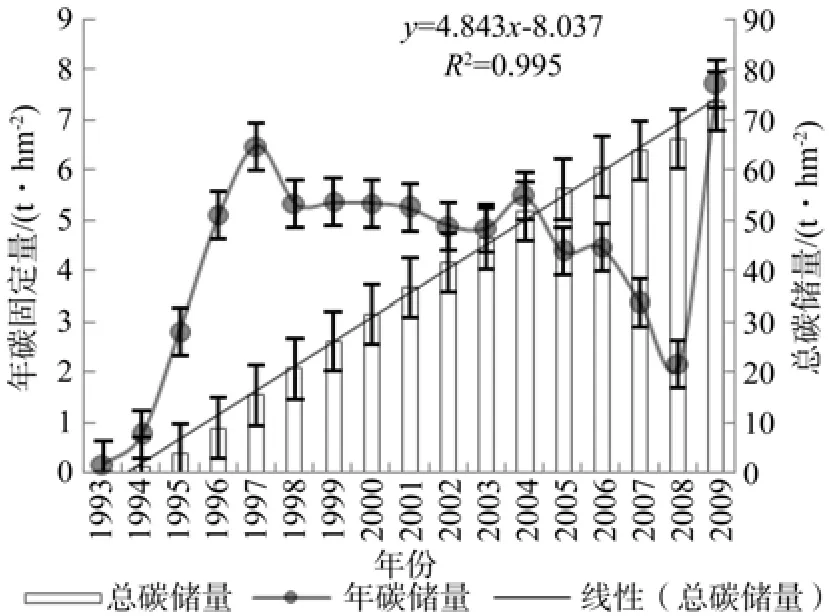
图 3 杉木乔木层碳储量及年固定量Fig.3 Carbon storage and annual storage in tree story of Chinese fi r
树木年轮是树木对环境因素的生理响应,可以估计树木的生长格局[25]。依据年轮宽度和相对生长模型估算的杉木乔木层年碳固定量来看,其每年碳固定量是不近相同的,到第5年时杉木年碳固定量达到6.49 t·hm-2a-1后, 而后基本维持在5 t·hm-2a-1左右。 2008年碳固定量最低,仅为2.152 t·hm-2a-1,与当年的气候变化有很大的关系,2008年的冰雪灾害严重影响了林木的生长,净生产力低。第二快速生长期出现在第16年,达到7.713 t·hm-2a-1。杉木人工林乔木层平均年碳固定量为4.347(±1.95) t·hm-2a-1,杉木林的年碳固定量随着年龄的增长呈现先增大后减少的趋势[26-27],与传统生态学林龄和生物量关系的Logistic模型一致[28],在幼龄和中龄阶段,随着林龄的增大生物量快速增长,之后生物量增长逐步变小直至达到近似平衡状态[29]。从杉木的连年生长曲线来看,该区域的林木处于高生长期,还没有达到生长拐点,碳蓄积潜力巨大。
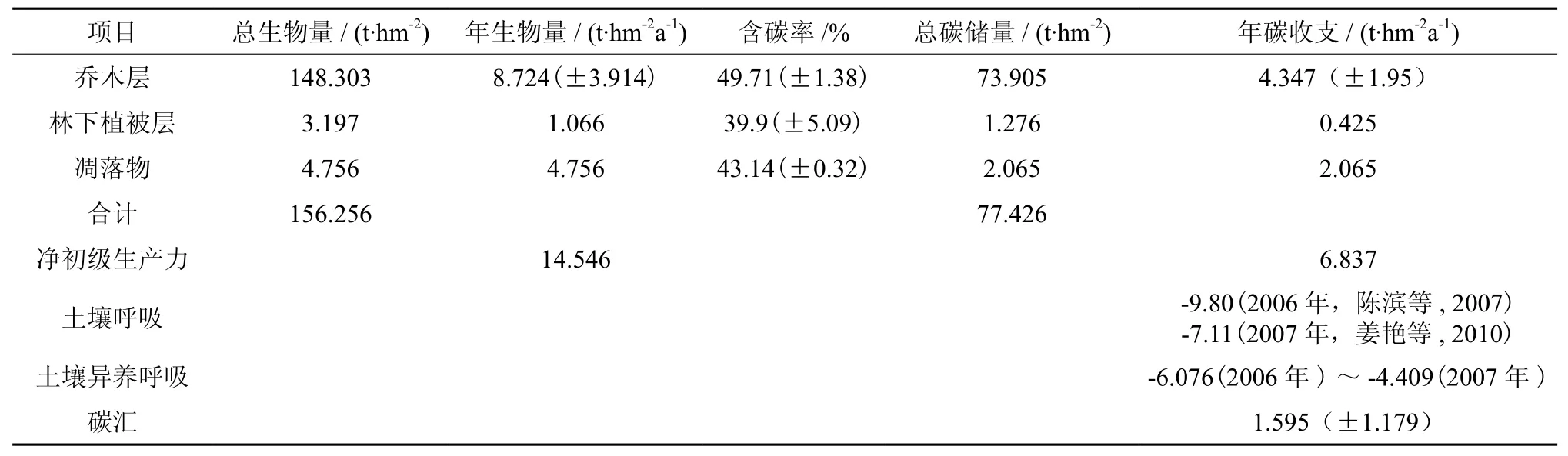
表 2 大岗山杉木人工林生态系统碳平衡(平均值±标准差)†Table 2 Carbon balance of ecosystem of Chinese fir plantation in Dagangshan (Mean±SD)
3.4 杉木林生态系统碳汇能力估算
16年生杉木人工林生态系统净初级生产力即生物碳库为6.837 t·hm-2a-1。土壤呼吸是土壤与大气交换CO2的过程,是土壤碳素同化和异化平衡的结果[3,30,41]。土壤异养呼吸碳通量占土壤呼吸碳通量的59%~65%[31],据姜艳、陈滨等在同一研究区域测定的土壤呼吸通量结果,杉木林生态系统土壤异养呼吸碳年支出量为-5.88~-4.266 t·hm-2a-1[32-33]。森林碳汇量为净初级生产力减去异养生物(土壤)的呼吸碳通量[23],杉木林生态系统的碳汇量为1.595(±1.179) t·hm-2a-1,杉木人工林生态系统是大气CO2的一个重要的汇,由此可见森林与全球大气CO2浓度的变化有着密切的关系,成为影响全球气候变化的一个重要因子。
4 结论与讨论
植物通过光合作用生产有机物储存在地上植物和地下植物中,固定大气中的CO2,形成生物量碳库,可以直接反映森林植被的固碳现状。16年生杉木人工林总生物量为156.256 t·hm-2,总碳储量为77.246 t·hm-2,高于会同15年生的杉木人工林总碳储量(57.34 t·hm-2)[34]。乔木层总生物量为148.303 t·hm-2,与江西千烟洲20年左右的杉木人工林的乔木层总生物量(140.88 t·hm-2)相差不大[15],也与其他地区比较接近[35,40]。杉木林还处于生长旺盛期,碳储存量潜力巨大,福建33年生杉木人工林总生物量达到236.069 t·hm-2,碳储量为 131.204 t·hm-2[36]。
杉木林地上部分总生物量(含林下植被层和凋落物)为 121.606 t·hm-2,碳储量为 60.556 t·hm-2,占77.83%,地下根系生物量为34.65 t·hm-2,碳储量为16.69 t·hm-2,占22.17%,两者比例为3.50,地上植被层碳储量最重要的和可见的碳库,也是人工林生产项目中测量和估算的唯一碳库[37]。地下生物量是也是总生物量碳库的重要组成部分,本研究没有考虑到死细根的碳储量,细根的碳周转率迅速,因此根系生物量碳的测定和估算相对低些,以后估算研究工作中要多加考虑。
年凋落物是地被层碳库和土壤碳库的主要来源,也是NPP的重要组成部分,大岗山杉木林2009年年凋落物量为4.756 t·hm-2a-1,碳储量为2.065 t·hm-2a-1,低于会同15年生杉木人工林的凋落物归还碳量(3.43 t·hm-2a-1)[34],高于大青山杉木人工林的凋落物归还碳量(1.37 t·hm-2a-1)[2],和福建33年生杉木人工林枯枝落叶层碳储量(2.28 t·hm-2a-1)相当[24]。本研究的凋落物量是2009年5月~2010年4月每月收集的新鲜枯落物总量,没有任何分解,枯落物碳储量相对于要高些,从凋落物角度分析,该杉木林年凋落量是较少的,对土壤自肥、保水与改良土质不利,其生态效益低。
森林生态系统生物量和生产力研究的主要内容之一是研究林木通过光合作用同化大气中的CO2,将其合成积累为有机物[42],林分年净生产力即碳积累速率决定森林生态系统的碳汇潜力[26]。在林分的生长过程中,年碳积累速率是不同的,幼龄林和中龄林碳积累速率大,到一定林龄碳积累速率就会趋于稳定。杉木林生态系统乔木层碳固定量为4.347(±1.95) t·hm-2a-1,与田大伦等(1998)用生物量法估算会同杉木人工林的净初级生产力为3.448~6.905 t·hm-2a-1一致[17],高于陈青青等(2012)估算了杉木林地上乔木层碳积累速率为0.107~0.618 t·hm-2a-1[26],与肖复明等(2010)估算15年生的杉木林为7.36 t·hm-2a-1相差较大[34]。整个杉木林生态系统净生产力为6.837 t·hm-2a-1,杨玉盛等(2007)估算33年生的杉木林为6.599 t·hm-2a-1[24],估算结果相差不大。
生态系统碳汇/源产生的根源是生态系统碳循环过程中的收支不平衡[29],大岗山杉木林人工林生态系统的年碳汇量为 1.595(±1.179)t·hm-2a-1。本研究采用土壤异养呼吸占土壤呼吸总量的59%~65%是适宜的,Raich和Schlesinger(1992)在全球尺度上对陆地生态系统土壤呼吸的研究认为植物的根系呼吸占土壤呼吸总量的30%~70%[38]。赵仲辉等(2011)用涡动相关通量法估算出13年生杉木人工林NEE为2.553 t·hm-2a-1[20],田大伦等(1998)用生物量法估算会同杉木人工林的NEE为1.922~3.858 t·hm-2a-1,所有研究表明杉木人工林具有碳汇功能[17]。方晰等(2002)认为杉木林生态系统10年生以前主要向大气释放CO2,10 年生后才具有CO2汇的功能[9],本研究也有同样发现杉木人工林林生态系统在幼龄阶段(前5 a)时净生产力较低,表现为碳源。
森林生态系统的碳汇量主要取决于NPP增长驱动的强度和长期趋势,导致更多的大气CO2进入到生态系统各碳库,使生态系统碳净吸收成为可能。2008年的灾害天气使得杉木生态系统乔木层的年净生产力低下,导致出现碳释放,形成碳源。王灿等用TRIPLEX1.6模型模拟了湖南杉木的林生产力NPP最大值为8.09 t·hm-2a-1[39],从NPP增长长期趋势来看,区域杉木人工林生态系统处于中龄林,净生产力呈强有力的上升趋势,随着年龄的增长,碳汇潜力巨大。
[1] 方精云,陈安平.中国森林植被碳库的动态变化及意义[J].植物学报,2001,43(9):967-973.
[2] 康 冰,刘世荣,蔡道雄, 等.南亚热带杉木生态系统生物量和碳素积累及其空间分布特征[J].林业科学,2009,45(8):147-152.
[3] Tianshan Zha, Zisheng Xing, Kai-Yun Wang, et al. Total and Component Carbon Fluxes of a Scots Pine Ecosystem from Chamber Measurements and Eddy Covariance[J]. Annals of Botany, 2007,99: 345-353.
[4] 邵全琴,杨海军,刘级远,等.基于树木年轮信息的江西千烟洲人工林碳蓄积分析[J]. 地理学报,2009,64(1):69-83.
[5] 方精云,郭兆迪,朴世龙, 等.1981-2000年中国陆地植被碳汇的估算[J].中国科学D辑:地球科学,2007,37(6):804-812.
[6] Fang J Y, Chen A P, Peng C H, et a1. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science,2001,292:2320-2322.
[7] NOPHEA SASAKI, SOPHANARITH KIM. Biomass carbon sinks in Japanese forests: 1966-2012[J]. Forestry, 2009, 82(1):113-123.
[8] Piao S L, Fang J Y, Zhu B, et a1. Forest biomass carbon stocks in China over the past 2 decades: estimation based on integrated inventory and satellite data[J]. Geophys Res,2005,110, G01006,doi: 10. 1029/2005JG00001.
[9] 方 晰,田大伦,项文化, 等.第二代杉木中幼林生态系统碳动态与平衡[J].中南林学院学报,2002,22(1):1-6.
[10] 方精云,刘国华,朱 彪,等.北京东灵山三种温带森林生态系统的碳循环[J]. 中国科学. D辑, 地球科学,2006,36(6):533-543.
[11] 贺金生.中国森林生态系统的碳循环:从储量、动态到模式[J].中国科学:生命科学,2012,42(3):252-254.
[12] 陈德祥,李意德, Liu H P,等.尖峰岭热带山地雨林生物量及碳库动态[J].中国科学:生命科学,2010,40:596-609.
[13] 杨同辉,宋 坤,达良俊,等.中国东部木荷-米楮林的生物量和地上净初级生产力[J].中国科学:生命科学,2010,40:610-619.
[14] 张全智,王传宽.6种温带森林碳密度与分配[J].中国科学:生命科学,2010,40:621-631.
[15] 马泽清,刘琪璟,徐雯佳,等.江西千烟洲人工林生态系统的碳蓄积特征[J].林业科学,2007,43(11):1-7.
[16] 雷加富.中国森林资源[M].北京,中国林业出版社,2005:172.
[17] 田大伦,盘宏华,康文星,等.第二代杉木人工林生物量的研究[J].中南林学院学报,1998,18:11-16.
[18] Zhao M F, Xiang W H, Peng C H, et al. Simulating age-related changes in carbon storage and allocation in a Chinese fir plantation growing in southern China using the 3-PG model[J].Forest Ecology and Management, 2009,257(6):1520-1531.
[19] 侯振宏,张小全,徐德应,等.杉木人工林生物量和生产力研究[J].中国农学通报,2009,25(5):97-103.
[20] 赵仲辉,张利平,康文星,等.湖南会同杉木人工林生态系统CO2通量特征[J].林业科学,2011,47(11):6-12.
[21] 王 丹,王 兵,戴 伟,等.不同发育阶段杉木林土壤有机碳变化特征及影响因素[J].林业科学研究,2009,22(5):667-671.
[22] 乔 磊,王 兵,郭 浩,等.江西大岗山地区7-9月降水量的重建与分析[J].生态学报,2011,31(8):2272-2280.
[23] 于贵瑞,伏玉玲,孙晓敏, 等.中国陆地生态系统通量观测研究网络的研究进展及其发展思路[J].中国科学D辑:地球科学,2006,36:1-21.
[24] 杨玉盛,陈光水,王义祥,等.格氏栲人工林和杉木人工林碳吸存与碳平衡[J].林业科学,2007,43(3):113-117.
[25] Adrien N. Djomo, Alexander Knohl, Gode Gravenhorst.Estimations of total ecosystem carbon pools distribution and carbon biomass current annual increment of a moist topical forest[J]. Forest Ecology and Management,2011,261:1448-1459.
[26] 陈青青,徐伟强,李胜功,等.中国南方4种林型乔木层地上生物量及其碳汇潜力[J].科学通报,2012,57(13):1119-1125.
[27] Chapin III F S, Matson P A, Mooney H A. Principles of Terrestrial Ecosystems Ecology[M]. New York: Springer-Verlag.2002:285-287.
[28] 徐 冰,郭兆迪,朴世龙,等.2000-2050年中国森林生物量碳库基于生物量密度与林龄关系的预测[J].中国科学:生命科学,2010,(40):587-594.
[29] 刘双娜,周 涛,魏林艳,等.中国森林植被的碳汇/源空间分布格局[J].科学通报,2012,57(11):943-950.
[30] 王 兵,姜 艳,郭 浩,等.土壤呼吸及其三个生物学过程研究[J].土壤通报,2011,42(2):438-490.
[31] 姜 艳,王 兵,汪玉如,等.亚热带林分土壤呼吸及其与土壤温湿度关系的模型模拟[J].应用生态学报,2010,21(7):1641-1648.
[32] 姜 艳.毛竹林土壤呼吸及其三个生物学过程的时空格局变化研究[D]. 北京:中国林业科学研究院,2010.
[33] 陈 滨,赵广东,冷 泠,等.江西大岗山杉木人工林生态系统土壤呼吸研究[J].气象与减灾研究,2007,30(3):12-16.
[34] 肖复明, 范少辉, 汪思龙, 等.毛竹杉木人工林生态系统碳平衡估算[J].林业科学,2010,46(11):59-65.
[35] 田大伦,方 晰,项文化.湖南会同杉木人工林生态系统碳素密度[J].生态学报,2004,24(11): 2382-2386.
[36] 杨玉盛,陈光水,王义祥,等.格氏栲人工林和杉木人工林碳库及分配[J].林业科学,2006,42(10):43-47.
[37] 李怒云,等.林业碳汇计量[M].北京,中国林业出版社,2009:17.
[38] Raich J W, Schlesinger W H. The Global Carbon Dioxide Flux in Soil Respiration and Its Relationship to Vegetation and Climate[J]. Tellus, 199, 44B: 81-99.
[39] 王 灿,项文化,赵梅芳,等.基于TRIPLEX模型的湖南省杉木生产力模拟及预测[J].中南林业科技大学学报,2012,32(6): 104-109.
[40] 邓华平,李树战,何明山,等.豫南不同年龄杉木林生态系统碳储量及其空间动态特征[J].中南林业科技大学学报,2011,31(8): 83-95.
[41] 卢 锟,项文化,邓湘雯,等.湘中丘陵区4种不同森林土壤CO2排放量比较[J].中南林业科技大学学报,2012,32(3):104-108.
[42] 沈 燕,田大伦,项文化,等.天然次生檫木枫香混交林生物量及生产力研究[J].中南林业科技大学学报,2011,31(5):26-30.
Study on carbon sink of Chinese fi r plantation ecosystem in Dagangshan mountain, Jiangxi province
PAN Yong-jun1, WANG Bing2, CHEN Bu-feng1, PENG Qing-zhen3
(1. Research Institute of Tropical Forestry, Chinese Academy of Forestry, Guangzhou 510520, Guangdong, China;2. Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China;3. Guangzhou Favorview Palace Experiment School, Guangzhou 510060, Guangdong, China)
The accurate research on carbon storage and carbon sinks function of forest ecosystems and is a basic works of ecology and plays a important role to appraise forest vegetation’s action in carbon balance of ecosystems. The carbon storage and carbon balance of the 16-year-old Cunninghamia lanceolata plantation ecosystems in Dagangshan mountain, Jiangxi province were investigated based on the data collected from long-term located observation. The main results of the research are as followings: the biomass of Chinese fi r forest was 156.256 t·hm-2, that of tree layer accounted for the 94.9% of total carbon storage, that of under-story biomass and annual litter amount were 3.197 t·hm-2, 4.756 t·hm-2, respectively; the total carbon storage was 77.246 t·hm-2, which was the largest carbon pool, the mean net primary production was 6.837 t·hm-2according to annual tree ring width. The carbon sink function was 1.595 (±1.179)(t·hm-2a-1). It is signif i cant that Chinese fi r plantation was an effective approach to the fi xed carbon. The capacity for forests to aid in climate change mitigation efforts is substantial, they can further mitigate the increase in atmospheric CO2that results from fossil fuel combustion and deforestation.
Chinese fi r plantation; biomass;carbon storage; forest carbon sink
S718.51+2.1
A
1673-923X(2013)10-0120-06
2012-10-16
国家“973”项目课题“天然森林和草地土壤碳储量及时空格局”(2011CB403200);林业公益性行业科研专项“森林生态服务功能分布式定位观测与模型模拟”(201204101);中国林业科学研究院热带林业研究所基础科研业务费专项资金项目:珠三角区域森林气候水文变化特征观测研究(生态定位站研究项目)(RITFYWZX201107)
潘勇军(1971-),男,湖南张家界人,博士,主要从事森林生态方面的研究;E-mail:Pyjzsj@126.com
[本文编校:文凤鸣]
