庐山森林景观格局变化的长期动态模拟
2013-12-21梁艳艳周年兴谢慧玮蒋铭萍
梁艳艳,周年兴,谢慧玮,蒋铭萍
(南京师范大学地理科学学院,南京 210023)
森林演替是森林生态系统的自组织过程,是指在某一地段上一个植物群落被另一个植物群落代替,直到形成顶极群落的生态过程[1],其核心在于自然和人为干扰条件下植被的动态演替过程及其演替规律。基于森林演替的研究可以为景观的动态变化提供基础,在以植被格局为基础的森林景观动态分析中,可通过森林演替推断景观格局的动态变化以及相应的景观生态过程[2]。森林景观格局的变化会对森林生态系统内的物质循环和能量流动产生影响,制约多种生态过程,进而影响森林的演替,如斑块的大小和形状会影响种群的生存能力和抗干扰能力[3-5]。因此,关于森林景观格局的研究一直是学术界研究的重点与热点。目前国内学者多集中于遥感、地理信息系统技术支持下对景观格局的控制要素和时空变化特征的研究[6-11],而关于在森林演替的内在机理和规律的基础上进行景观格局动态变化分析方面的研究较少。
从科学方法论的角度看,方法的研究要完成4个连续的层次,即:描述研究、解释研究、预测研究和规范研究[12]。近年来,伴随着景观生态学研究的不断深入,生态学者逐渐克服直接调查和观测的局限,在已有的科学发现和机理认识的基础上,逐步关注复杂系统的模拟和科学预测,以便为调控生态系统奠定科学基础[13]。因此,景观空间动态模型应运而生。典型的景观空间动态模型主要包括林窗模型、马尔科夫模型和LANDIS模型。林窗模型主要用来探讨森林长期动态变化问题[14-15],但不包括种子传播和人为干扰等大的空间范围上的生态过程;马尔科夫模型则通过确定景观转移概率来模拟景观空间格局动态,一般不考虑景观的变化机制和动态过程[16-17];LANDIS模型用来模拟空间范围内的多种生态过程,包括森林演替、种子传播、物种定植、各种干扰以及它们之间的相互作用,并被广泛应用于森林景观变化的研究中[18-26]。景观变化的模拟不单要了解一种景观到另一种景观的变化,更重要的是要清楚发生变化的内在机理,林窗模型和马尔科夫模型的不足之处在于缺乏基于植被演替机理上的生态过程的表达,而LANDIS模型克服了这些弊端,增加对森林景观格局和生态过程关系的理解。
庐山是研究植被顺向演替和恢复生态学的“天然实验室”[27],模拟研究未来较长时间尺度内森林群落的演替,实现对森林景观格局的动态监测,对于我国南方开展亚热带山地次生植被的生态恢复工程具有典型的示范意义。鉴于此,本文以庐山风景区为案例地,尝试在LANDIS模型模拟森林植被未来300 a的演替动态基础上,分析森林景观格局随时间推移的动态变化特征,揭示森林景观演替的规律,预测景观的未来变化趋势,以期为森林景观的可持续发展提供理论参考和应用借鉴。
1 研究区概况
庐山地处江西省九江市东南,北临长江,东濒鄱阳湖,介于东经 115°52'—116°08'、北纬 29°26'—29°41'之间。庐山位于我国东部亚热带中部的北缘,与长江以北的亚热带北部相邻,气候上具有亚热带中部过渡到亚热带北部的特点。全年平均温度为11.4℃,最高只有32℃,最低-16.8℃。庐山雨量丰沛,全年平均降雨量1917 mm,年平均有雨日达168 d。庐山土壤的垂直结构类型共有5个:自山麓至山顶,依次分布着红壤和黄壤(400 m以下)、山地黄壤(400—800 m)、山地黄棕壤(800—1200 m)、山地棕壤(1200 m以上)[28]。考虑到庐山复杂的管理体制和空间上的连续性,本文选取庐山风景名胜区管理局管辖范围作为研究区(图1),面积为 121 km2,海拔71—1446 m。
庐山的自然环境复杂,生态系统完整,森林覆盖率高,地带性植被是亚热带常绿阔叶林,其植被受人为干扰影响剧烈,现状植被属于在自然恢复中的次生植被,呈现出较为明显的垂直分布特征。本文结合2010年完成的庐山林相调查图,根据《江西森林》的植被分类系统[29],在野外调查的基础上,确认庐山森林植被类型中的优势种主要包括樟树(Cinnamomum camphora)、苦槠(Castanopsis sclerophylla)、锥栗(Castanea henryi)、枫香(Liquidamba formosana Hance)、马尾松 (Pinus massoniana Lamb)、台湾松 (Pinus taiwanensis)、杉木(Cunninghamia lanceolata)、日本柳杉(Cryptomeria japonica)、日本扁柏(Chamaecyparis obtusa)和毛竹(phyllostachys heterocycla)等。由于这10个树种占据整个庐山森林植被面积的85%以上[27],为了研究方便,本文以这10个优势树种群落代表整个庐山森林植被,并将优势树种作为森林植被斑块组成的标志。
2 研究方法
2.1 数据来源与处理
定量化的景观格局分析必须建立在基础地理信息数据库之上。本研究中的地理信息数据来源主要是庐山林相调查图、1∶10000地形图、相关经验参数以及研究者实际调查的第一手资料。

图1 庐山风景区范围图Fig.1 Areal map of Mount Lushan
运用LANDIS模型进行模拟,将生成的结果在Arc GIS10.0软件中栅格化,并导入景观格局分析软件Fragstats 3.3,在斑块类型和景观水平上计算相关的景观格局指数。
2.2 LANDIS 模型
LANDIS模型是主要用于模拟森林演替、种子扩散、干扰和管理的空间直观景观模型。该模型把景观看作有相同大小的样地(像元)组成的网格,通过跟踪样地上物种的存在与否来模拟风火、种子扩散、采伐等自然和人为干扰下景观尺度上森林的动态变化。每个像元都记录树种的组成信息和干扰特征,并不记录每个物种的实际年龄,而是记录以10 a为间隔的年龄组,通过跟踪年龄组的缺失来模拟在自然和人为干扰条件下森林的动态变化[18]。
运行LANDIS模型所必需的参数包括两大类:GIS图形参数文件和属性参数文件。GIS图形参数包括森林植被图、立地类型图;属性参数主要包括物种生活史参数、物种建群系数和各种自然干扰参数。
2.2.1 森林植被图
森林植被图以林相图为基础,并对每个像元输入物种分布以及年龄信息。采用基于小班的随机赋值法,对每个像元进行赋值[30]。根据林相调查图,考虑到计算机模拟能力,将整个研究区重采样到30 m×30 m分辨率,得到一幅553行 ×531列的初始物种/年龄级分布图。
2.2.2 立地类型图
LANDIS模型将异质的景观分成相对均质的土地类型单元,同一种类型的土地具有相同的环境条件。庐山植被具有典型的垂直地带性分布规律,本研究中主要根据庐山的地貌将研究区划分为7种立地类型:海拔≤400 m、400 m<海拔<800 m、800 m≤海拔≤1200 m、海拔>1200 m、水域、建设用地和农耕地(图2)。所有立地类型都是根据庐山林相调查图、1∶10000地形图和研究者实地调查获得。
LANDIS模型将立地类型划分为无效立地类型(不模拟)和有效立地类型(模拟)。本研究中,无效立地类型包括水域、建设用地和农耕地,占整个研究区的8.18%,有效立地类型包括海拔≤400 m、400 m<海拔<800 m、800 m≤海拔≤1200 m、海拔>1200 m区域,分别占整个研究区面积的 21.15%,29.23%,32.89% 和8.53%。
2.2.3 物种生活史参数
物种演替是由物种不同的生理特性驱动的。在LANDIS模型中,物种的竞争能力主要由寿命、耐阴性、耐火性、成熟年龄、萌发能力和种子传播距离物种等特性决定。本研究区中10个优势树种的生活史参数通过查阅相关文献和实地调查获得[27,31-35],具体的参数值见表1。
2.2.4 物种建群系数
LANDIS模型只跟踪模拟有效立地利用类型上的物种变化。当种子扩散到某地后,由于受到该地区所处的环境因素(水、光、土壤、气候等)以及物种自身的生物学特性等因素的影响,不可能都存活。LANDIS模型通过物种的建群系数来反映物种在各立地类型上是否能够存活并且正常生长的能力。在同一立地类型内,每一个物种的建群系数相对一致,建群系数最敏感的范围为0.05—0.3[36]。通过查阅参考文献和咨询相关林业专家[27,32,37-39],得到物种在不同立地类型中的建群系数(表 2)。

图2 研究区立地类型图Fig.2 Land type map of study area

表1 庐山优势树种生活史参数Table 1 Dominant species attributes for Mount Lushan

表2 物种在各立地类型上的建群系数Table 2 Establishment coefficient for each species in all land types
值得注意的是,庐山是世界文化景观遗产所在地,整个研究区范围全部为生态公益林,禁止采伐;整个风景区火干扰发生的次数较少,而且主要是雷击引起的自然火;景区所受风的干扰很小;近几年来病虫害的发生呈上升趋势,但是这方面资料比较匮乏。因此,本研究不考虑任何自然干扰参数和人为采伐,在没有外界大的干扰下完全依靠种子的传播来模拟森林的演替。
2.3 景观格局变化分析
基于森林演替的研究可以为景观格局的动态变化提供基础,景观格局变化的定量分析可以从景观指数的变化上反映出来。景观格局指数高度浓缩了景观格局和景观动态信息,能够很好地了解景观格局的组成成分、空间配置和动态变化过程,是景观生态学最常用的定量化方法。本文结合庐山实际情况,所选取的景观格局指数见表 3[40]。
3 结果及分析
由于LANDIS模型的随机性,它本身的不确定性需要重复模拟来解决。而对于模拟结果的验证,LANDIS模型已经在众多不同的环境区域得到了广泛应用,适用于多种森林类型,模拟结果理想,在国际上具有较高的可信度[41-43]。在自然演替的情况下,庐山森林植被演替系列为草丛—灌丛—针叶林—针阔混交林—常绿阔叶林或者是落叶阔叶林—常绿落叶阔叶混交林—常绿阔叶林[27]。从本文的模拟结果可以看出,各群落中的物种组成和优势树种与本地区的实际情况及其前人的研究相一致[9]。
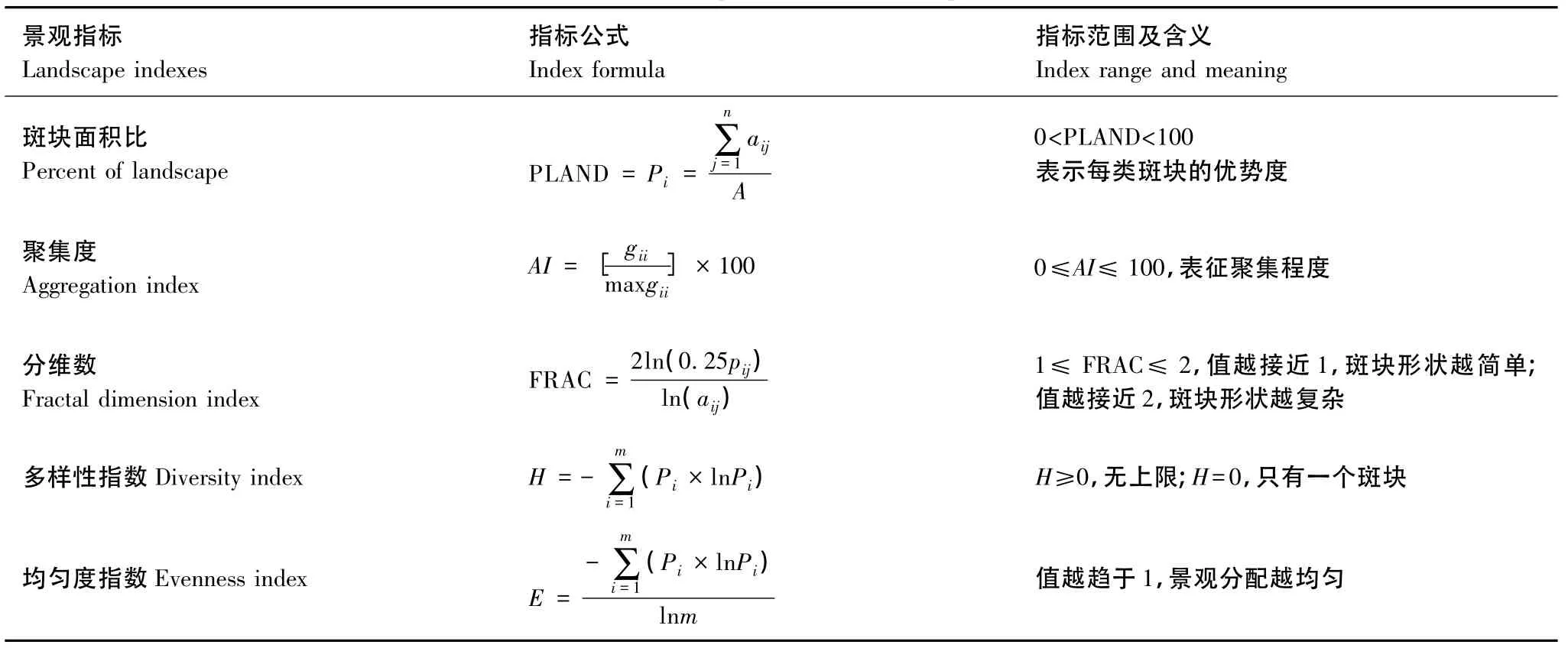
表3 本研究所采用的景观定量评价指数Table 3 Landscape indexes and its implication
3.1 斑块面积比变化
斑块面积比表示某一斑块类型占整个景观面积的相对比例,是帮助人们确定优势景观类型的依据之一。因为LANDIS模型只记录像元上树种的有无,同时一个像元上可能存在多个树种,所以树种所占的面积比之和可能会大于100%。图3是森林景观从当前环境开始模拟300 a以后的结果,它表现了在不同演替时期中各优势树种植被斑块所占的面积比例。
模拟的所有优势树种占庐山植被总面积比的变化趋势表明,未来300 a的时间里,庐山的森林植被将朝着地带性植被常绿阔叶林方向演替。整体来看,常绿阔叶林是研究区的景观基质,在控制景观整体的结构、功能和动态过程中起着主导作用。阔叶林树种樟树、苦槠、枫香和锥栗的面积比在300 a内增幅非常明显,保持在24%—55%之间。由于阔叶林的种子具有较强的传播和定居能力,属于增长型种群。阔叶林树种又具有极强的耐阴性,随着演替的进行会获得更大的生态位,分布面积将保持平稳增长趋势。
杉木林保持平稳的小幅度增长趋势。庐山的杉木林主要分布于海拔800 m以下的缓坡上,大多呈现纯林现象,有的地段混有一些阔叶树种,有的地段与毛竹混交。随着龄组的增加,并伴随着种群对资源的强烈竞争,逐步过渡到随机分布状态,所以呈现出平稳的面积比率增长态势。
先锋树种马尾松和台湾松林的面积在演替初始阶段都呈现小幅增长,并保持稳定趋势至100 a,然后突然锐减,直至200 a,随后保持较小的面积比例直至演替结束。马尾松和台湾松都是阳性喜光的树种,在土壤肥沃的地方生长良好,多与阔叶树混生,组成针阔混交林。随着演替的进行,它们的幼苗不能在自身林冠下更新,逐渐被阔叶树种入侵,面积比例将逐渐下降,与图3中的曲线相吻合。
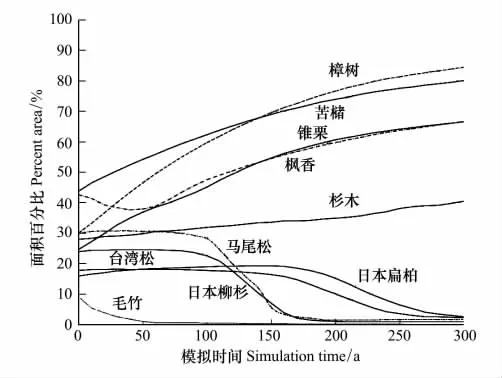
图3 庐山森林植被演替的模拟结果Fig.3 The result of forest vegetation succession in Mount Lushan
日本柳杉和日本扁柏林在演替的前150 a呈现平稳的小幅增长趋势,后150 a呈现缓慢下降的趋势。日本柳杉和日本扁柏是20世纪60年代人工引进的外来树种,寿命长,适生环境大致相同,如今这些人工针叶林已经遍布庐山的山顶[44]。由于这些人工群落成熟林逐渐增多,没有人为影响会在2160年前后保持相对稳定,但是由于结构单一,林冠郁闭度高,林下植物生长稀疏,天然更新困难,待群落衰退以后将逐渐为其它相应类型的植被所取代。
毛竹林的面积比例一直在不断下降。现有毛竹林多为人工栽植,由于庐山所处纬度偏高,又由于海拔较高,在与其它树种的竞争中处于劣势,其从幼龄林至衰老的时间大约为60 a。从长时间看,在没有人为因素影响下,最终将会逐渐衰老,为阔叶树种所更替。
3.2 景观聚集度变化
聚集度指数用于衡量景观尺度内物种空间分布格局的聚集水平。根据林相调查图资料,在2010年,毛竹林的聚集度最高,为98.41%,其次为马尾松和日本扁柏林,分别为98.29%、98.14%,苦槠林的聚集度最低,为95.76%,各优势树种植被斑块的聚集度都处于较高水平。通过LANDIS模型模拟,聚集度指数发生了变化(表4)。
从表4可以看出阔叶林树种聚集度都呈现出在前150 a缓慢增长,后150 a保持相对稳定的特征;杉木林的聚集度一直保持平稳的状态;马尾松和台湾松的寿命都在300 a左右,不会在短时间内退出演替过程,并在一定程度上发挥建群种的主要优势,在2210年左右,马尾松和台湾松林都进入老龄林阶段,聚集度迅速下降直至演替终期;日本柳杉和日本扁柏林的聚集度在前150 a相对稳定,后150 a表现为缓慢降低的现象;毛竹林的聚集度在整个模拟阶段一直在不断下降,直至演替结束。
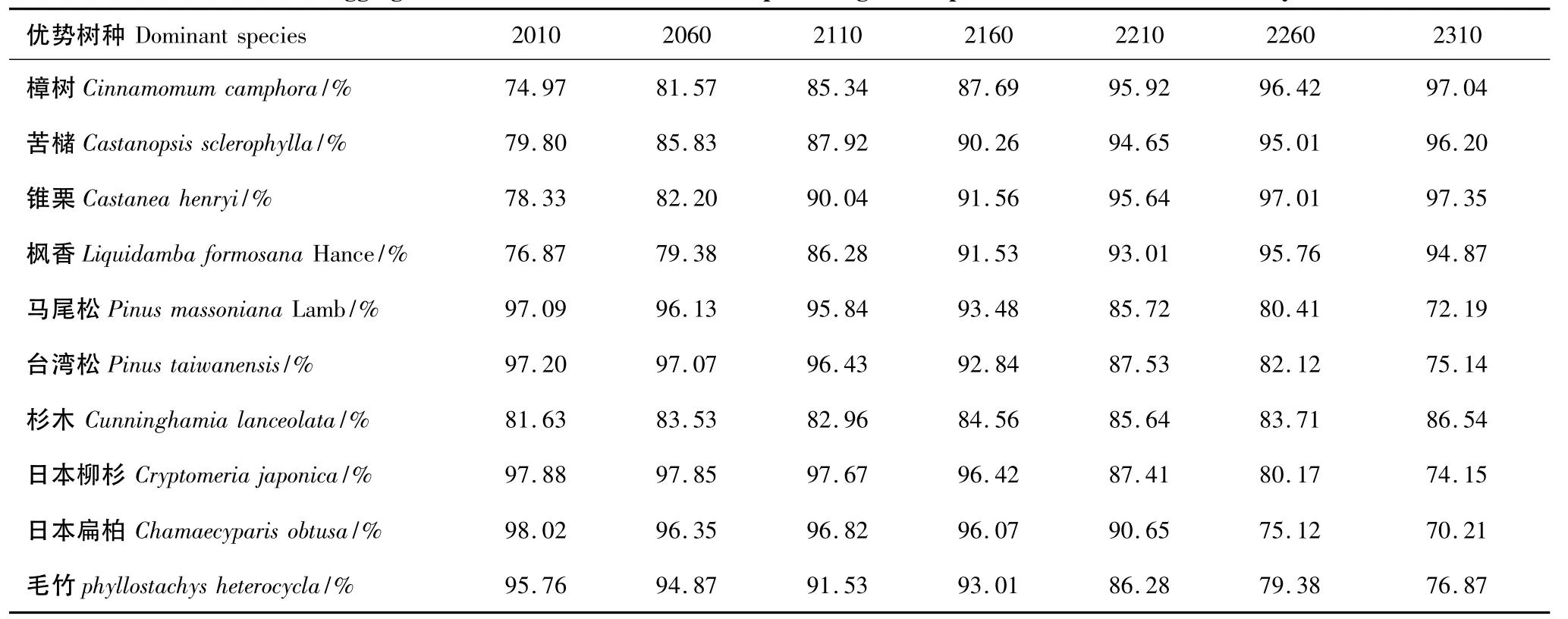
表4 不同模拟年份各优势树种植被斑块的分布聚集度Table 4 Aggregation index of various dominant species vegetation patches in different simulation years
3.3 景观分维数变化
分维数直接反映一定程度上斑块边界形状复杂性及破碎化程度。整体来看,在模拟的300 a内,各优势树种植被斑块的分维数都保持在1—1.1之间,说明各景观斑块的边缘相对比较规则且变化较小(表5)。2010年枫香林的分维数最高,为1.0691,而毛竹林最低,表明枫香林的景观形状最复杂,毛竹林的景观形状最简单,连通性最好。分维数随着时间推移呈现不断下降的趋势,到2160年,台湾松林的分维数保持最高水平1.0389,毛竹林保持在最低水平1.0052;到2310年,分维数的排列出现了变化,樟树林最高,为1.0311,苦槠林次之(1.0283),最低的仍然是毛竹林(1.0063)。毛竹林一直都处于最低水平,主要是由于毛竹林多为纯林景观,可以在短期内迅速扩张,并保持较好连通性。

表5 不同模拟年份各优势树种植被斑块的分维数指标Table 5 Fractal dimension index of various dominant species vegetation patches in different simulation years
3.4 景观多样性变化
景观多样性指数能反映一个区域内不同景观类型分布的均匀度和复杂化程度。本文选取多样性指数和均匀度指数两个指标来反映景观多样性特征。随着时间的推移,研究区景观多样性指数呈现先上升后保持缓慢下降的趋势(图4),均匀度指数则呈现先下降后上升再缓慢下降的变化态势(图5)。
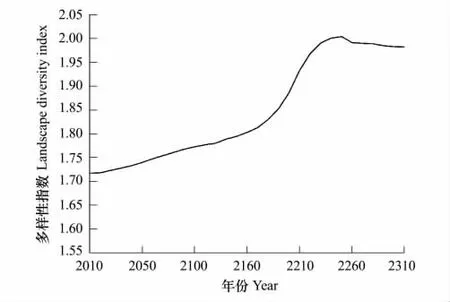
图4 不同模拟年份景观多样性指数变化图Fig.4 Change chart of landscape diversity in different simulation years
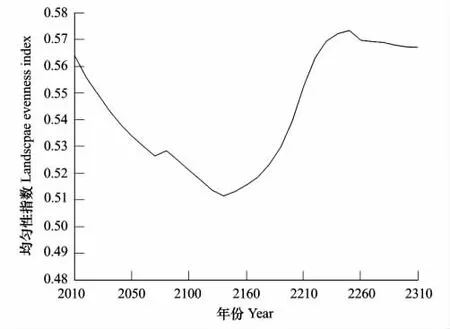
图5 不同模拟年份景观均匀度指数变化图Fig.5 Change chart of landscape evenness in different simulation years
庐山森林植被景观多样性指数由2010年的1.717上升到2.0047(2250年),此后的60 a又保持缓慢下降趋势,至2310年为1.9827;均匀度指数由2010年的0.564下降到0.5132(2150年),此后逐渐上升到0.5734(2250年),随后又呈缓慢下降趋势,到2310年降为0.5671。在一个景观系统中,土地利用类型越丰富,破坏化程度越高,其景观多样性指数越高。在模拟的前140 a里,景观的多样性指数增加,均匀度却下降,两者呈现负相关;随着演替的进行,140 a后景观多样性指数和均匀度指数变化趋势相同,都呈现先上升后缓慢下降的趋势,这与Cusack和Montagnini所做的研究一致[45]。到2250年,森林群落中人工针叶林的成熟林增至最多,多样性和均匀度指数上升至最高。此后随着人工群落的逐渐衰退,加上马尾松和台湾松的天然更新困难,阔叶林等乡土物种通过改善林地小气候,吸引种子传播及促进乔木树种的天然更新,进而促进森林植被逐渐恢复成稳定的地带性植被[27,35,39],使得森林的多样性和均匀度指数又不断下降,景观破碎化程度逐渐降低。总体来看,景观多样性指数变化的特征与整个森林朝着顶极群落常绿阔叶林方向演替的趋势相吻合。
4 结论
模拟研究庐山未来较长时间尺度内森林群落的演替,对于我国南方开展亚热带山地次生植被的生态恢复工程具有典型的示范意义。通过LANDIS模型对庐山森林景观的长期动态模拟的结果,可以得出以下几点结论:
(1)庐山森林群落的演替,依据一定的客观规律向最优化的顶极群落演变。阔叶林树种的绝对优势地位保证了森林植被的天然更新。分析发现,森林群落演替过程中各个阶段的表现特征是通过优势树种的数量增长和消亡来表达的。按照这样的规律,到群落相对稳定时,研究区内绝大部分为常绿阔叶林及常绿落叶阔叶混交林。从长远来看,应该继续实行严格的封山育林政策。
(2)景观格局指数的变化特征与植被向顶极群落演替的趋势相吻合。斑块面积百分比特征方面,阔叶林树种呈现持续增长的稳定趋势;景观聚集度特征方面,阔叶林树种在前150 a缓慢增长,而后150 a保持相对稳定,杉木林一直保持平稳,毛竹林在整个模拟阶段一直在不断下降直至演替结束;分维数特征方面,各优势树种植被斑块都保持在1—1.1之间,说明各景观斑块的边缘相对较规则且变化较小;景观多样性特征方面,多样性指数呈现出先上升后缓慢下降的趋势,而均匀度指数则呈现出先下降后上升再缓慢下降的变化态势。
(3)未来300 a的时间内,20世纪60年代实施的人工造林方案未对庐山森林景观格局产生明显影响。虽然人工林会造成森林林分简单,形成单优群落,但是由于针叶林树种的生活周期漫长,有的甚至达到几百年,它们的繁殖更新一般不会太快,对森林群落的入侵性要比草本植物小很多[46]。未来较长时间尺度内,外来树种只是扮演一个过渡的角色,对庐山的森林景观格局影响较小。
5 讨论
本文运用LANDIS模型来模拟森林自然演替状态下庐山景观格局的动态变化,还存在以下几个方面的局限性需要探讨:
(1)由于缺乏过去完整的植被图,本文的研究仅以2010年的林相图作为基础,未能考虑到之前的森林植被的变化历史对未来的影响。300 a的模拟过程只是漫长恢复演替过程中的一部分,更多的演替规律和过程还需要进一步的研究,此外模型是否需要修正还需要进一步的研究,未来可尝试与其它方法相结合,提高模拟结果的可信度。
(2)本研究是基于庐山森林植被演替得到的模拟结果,尚未考虑一些自然干扰和人为干扰因素,比如全球变暖、虫害以及森林管理方案的影响。如能对这些因素的影响进行模拟,则可从理论上阐明影响亚热带地区山地次生植被演替的驱动机制,为退化植被的生态恢复和森林景观的可持续发展奠定科学基础,这也是今后模型模拟工作的一个重要方向。
(3)本研究的景观格局指数受到空间分辨率和粒度的影响,由于聚集度指数和分维数指数的粒度效应比较明显[47],计算的精确性仍然值得进一步探讨。未来可尝试研究不同空间分辨率和粒度影响下景观格局指数间的比较,分析其变化特征。
尽管LANDIS模型存在上述局限性,但从机理上认清景观格局与森林演替之间的关系,对于森林景观格局而言,有助于其研究向生态背景的深层次内涵拓展,丰富景观格局研究的方法,为森林景观格局动态变化的研究提供新的思路;对恢复生态学而言,使森林的演替赋予了定量化的分析,丰富了恢复生态学的实践价值。致谢:感谢中国科学院沈阳应用生态研究所景观过程小组为本文在数据处理和模型调试方面提供的帮助,感谢庐山植物园相关专家提供的庐山森林植被的相关经验参数。
[1]Peng SL.Restoration Ecology.Beijing:China Meteorological Press,2007:39-39.
[2]Xu L H,Liu H Y,Chu X Z,Su K.Indication of desert vegetation to landscape patterns and dynamics at the northern edge of Tianshan Mountains.Acta Ecologica Sinica,2004,24(9):1966-1973.
[3]Fu B J,Chen L D,Ma K M,Wang Y L.The Principles and Application on Landscape Ecology.Beijing:Science Press,2001:126-133.
[4]Fahrig L,Merriam G.Habitat patch connectivity and population survival.Ecology,1985,66(6):1762-1768.
[5]Kratz T K,Benson B J,Blood E R,Cunningham G L,Dahlgren R A.The influence of landscape position on temporal variability in four North American ecosystems.The American Naturalist,1991,138(2):355-378.
[6]Tang L J.The spatial pattern analysis of landscape and the landscape planning in Sheshan Scenic Spot.Acta Geographica Sinica,1998,53(5):429-437.
[7]Ding SY,Qian L X,Cao X X,Li S,Li H M.Forest landscape patterns dynamic of Yihe-louhe River Basin.Acta Geographica Sinica,2003,58(3):354-362.
[8]He D J,Hong W,Hu H Q,Wu CZ,Chen X Z.Modeling landscape pattern dynamics and their effects under different disturbances in Wuyishan Scenery District.Acta Ecologica Sinica,2004,24(8):1602-1610.
[9]Hu H S,Wei M C,Tang JG,Zhang F Q,Zheng Y P.The landscape pattern changes and simulation in Lushan Mountain national Park.Acta Ecologica Sinica,2007,27(11):4696-4706.
[10]Zhao Q,Zheng G Q,Huang Q H.Characteristics of urban forest landscape pattern and optimization of urban forest spatial structure:A case study of Nanjing city.Acta Geographica Sinica,2007,62(8):870-878.
[11]Zhao F Q,Dai L M,Yu D P,Zhou L.Dynamic changes of forest landscape pattern in Lushuihe Forest Bureau of Changbai Mountains.Northeast China.Chinese Journal of Applied Ecology,2010,21(5):1180-1184.
[12]Guo L X,Bao JG.Review on Chinese tourist geography and its prospect.Geographical Research,1990,9(1):78-87.
[13]Ecological Society of China.Report on Advances in Ecology.Beijing:Chinese Science and Technology Press,2010:6-6.
[14]Botkin D B.Forest Dynamics:An Ecological Model.Oxford:Oxford University Press,1993.
[15]Yan X D.Several basic issues of forest gap model Ⅰ.Effect of simulated plot area.Chinese Journal of Applied Ecology,2001,12(1):17-22.
[16]Horn H S.Forest succession.Scientific American,1975,232(5):90-98.
[17]Patil GP,Taillie C.A multiscale hierarchical Markov transition matrix model for generating and analyzing thematic raster maps.Environmental and Ecological Statistics,2001,8(1):71-84.
[18]He H S,Mladenoff D J.Spatially explicit and stochastic simulation of forest landscape fire disturbance and succession.Ecology,1999,80(1):81-99.
[19]He H S,Hao Z Q,Larsen D R,Dai L M,Hu Y M,Chang Y.A simulation study of landscape scale forest succession in northeastern China.Ecological Modelling,2002,156(2/3):153-166.
[20]Hu Y M,Xu C G,Chang Y,Li X Z,Bu R C,He H S,Leng W F.Application of spatially explicit landscape model(LANDIS):A case researches in Huzhong area,Mt Daxing'anling.Acta Ecologica Sinica,2004,24(9):1846-1856.
[21]Wang X G,Li X Z,He H S,Xie F J.Evaluation of landscape restoration in the northern slopes of Great Xing'an Mountains after the 1987 catastrophic fire.Acta Ecologica Sinica,2005,25(11):3098-3106.
[22]Wang X G,Li X Z,He H S.Long-term effects of different management strategies on Larix gmelinii forests in Great Hing'an Mountains after the catastrophic fire in 1987.Chinese Journal of Applied Ecology,2008,19(4):915-921.
[23]Gong X,Chang Y,Bu R C,Li X Z,Xu C G,Yu Q H.Long-term effects of different forest harvesting modes on forest landscape pattern in Huzhong Forestry Bureau.Chinese Journal of Ecology,2006,25(7):805-812.
[24]Zhou Y F,He H S,Bu R C,Jin L R,Li X Z.Modeling of forest landscape change in Xiaoxing'anling Mountains under different planting proportions of coniferous and broadleaved species.Chinese Journal of Applied Ecology,2008,19(8):1775-1781.
[25]Guo R,Bu R C,Hu Y M,Chang Y,He H S,Liu X M,Zhang Z Q.Simulation of timber-harvesting area in Xiaoxing'anling Mountains under climate change.Chinese Journal of Applied Ecology,2010,21(7):1681-1688.
[26]Chen H W,Hu Y M,Chang Y,Bu R C,Li Y H,Liu M.Simulating long-term effects of different harvesting modes on forest fire in Huzhong Forest Region,northeastern China.Journal of Beijing Forestry University,2011,33(5):13-19.
[27]Liu X Z,Wang L.Scientific Survey and Study of Biodiversity on the Lushan Nature Reserve in Jiangxi Province.Beijing:Science Press,2010:3-293.
[28]Huang R C,Dai Z H,Chen B B,Lu B S.Characteristics of the soils of the Lushan area,central China.Acta Pedologica Sinica,1957,5(2):117-135.
[29]Jiangxi Forest Editorial Board.Jiangxi Forest.Beijing:China Forestry Press,1986.
[30]Xu C G,Hu Y M,Chang Y,Li X Z,Bu R C,He H S,Leng W F.The effect of cell-level uncertainty on spatially explicit landscape model simulation.Acta Ecologica Sinica,2004,24(9):1938-1949.
[31]Guo Z H,Zhuo Z D.A study on flora geography in Lushan Mountain.Tropical Geography,1996,16(1):72-81.
[32]Zhang Y.Animals and Plants in Jiangxi Province.Beijing:The Central Party School Press,1994:45-57.
[33]Yang JG.The distribution and environmental research of Cryptomeria in China.Jiangxi Forestry Science and Technology,1980,(2):11-14.
[34]Peng SL,Fang W.Studies on dynamics of Castanopsis Chinensis and Schima Superba population in forest succession of Dinghushan Mountain.Acta Phytoecologica Sinica,1995,19(4):311-318.
[35]Peng SL.Comparison of growth dynamics between first generation and regeneration of Pinus mossomiana.Chinese Journal of Applied Ecology,1995,6(1):11-13.
[36]He H S,Mladenoff D J,Gustafson E J.Study of landscape change under forest harvesting and climate warming-induced fire disturbance.Forest Ecology and Management,2002,155(1/3):257-270.
[37]Lin X,Hong C D,Si JP,Zeng F L,Ji P F.The preliminary study of Chamaecyparispisifera and Chamaecyparis'growth rule.Forest Science and Technology,1987,(12):10-13.
[38]Luo S J,Yi Y M,Zou H Y,Liang S W,Wu F,Wu W J.Germination percentage and complex stressing vigor test(CSVT)of Pinus hwangshanensis.Hubei Forestry Science and Technology,2001,(1):9-11.
[39]Chen L C,Wang S L,Chen C Y.Degradation mechanism of Chinese fir plantation.Chinese Journal of Applied Ecology,2004,15(10):1953-1957.
[40]Wu J G.Landscape Ecology:Pattern,Process,Scale and Hierarchy.2nd ed.Beijing:Higher Education Press,2007:106-115.
[41]Loehle C.A hypothesis testing framework for evaluating ecosystem model performance.Ecological Modelling,1997,97(3):153-165.
[42]Xu C G,Hu Y M,Jiang Y,Chang Y,Li X Z,Bu R C,He H S.Validation of spatially explicit landscape model.Chinese Journal of Ecology,2003,22(6):127-131.
[43]Wang X G,Li X Z,He H S.Spatial simulation of forest succession under different fire disturbances and planting strategies in northern slopes of Great Xing'anling Mountains.Journal of Beijing Forestry University,2006,28(1):14-22.
[44]Wan H L,Feng Z W,Pang H D.On the exotic plants in Lushan,Jiangxi Province,China.Acta Ecologica Sinica,2008,28(1):103-110.
[45]Cusack D,Montagnini F.The role of native species plantations in recovery of understory woody diversity in degraded pasturelands of Costa Rica.Forest Ecology and Management,2004,188(1/3):1-15.
[46]Zheng Y Q,Zhang C H.Current status and progress of studies in biological invasion of exotic trees.Scientia Silvae Sinicae,2006,42(11):114-122.
[47]Shen WJ,Wu JG,Lin Y B,Ren H,Li QF.Effects of changing grain size on landscape pattern analysis.Acta Ecologica Sinica,2003,23(12):2506-2519.
参考文献:
[1]彭少麟.恢复生态学.北京:气象出版社,2007:39-39.
[2]徐丽宏,刘鸿雁,楚新正,苏凯.天山北麓典型地段植被对景观格局和动态的指示意义.生态学报,2004,24(9):1966-1973.
[3]傅伯杰,陈利顶,马克明,王仰麟.景观生态学原理及应用.北京:科学出版社,2001:126-133.
[6]唐礼俊.佘山风景区景观空间格局分析及其规划初探.地理学报,1998,53(5):429-437.
[7]丁圣彦,钱乐祥,曹新向,李爽,李昊民.伊洛河流域典型地区森林景观格局动态.地理学报,2003,58(3):354-362.
[8]何东进,洪伟,胡海清,吴承祯,陈先珍.武夷山风景名胜区景观空间格局变化及其干扰效应模拟.生态学报,2004,24(8):1602-1610.
[9]胡海胜,魏美才,唐继刚,张福庆,郑艳萍.庐山风景名胜区景观格局动态及其模拟.生态学报,2007,27(11):4696-4706.
[10]赵清,郑国强,黄巧华.南京城市森林景观格局特征与空间结构优化.地理学报,2007,62(8):870-878.
[11]赵福强,代力民,于大炮,周莉.长白山露水河林业局森林景观格局动态.应用生态学报,2010,21(5):1180-1184.
[12]郭来喜,保继刚.中国旅游地理学的回顾与展望.地理研究,1990,9(1):78-87.
[13]中国生态学学会.生态学学科发展报告.北京:中国科学技术出版社,2010:6-6.
[15]延晓冬.林窗模型的几个基本问题的研究 Ⅰ.模拟样地面积的效应.应用生态学报,2001,12(1):17-22.
[20]胡远满,徐崇刚,常禹,李秀珍,布仁仓,贺红士,冷文芳.空间直观景观模型LANDIS在大兴安岭呼中林区的应用.生态学报,2004,24(9):1846-1856.
[21]王绪高,李秀珍,贺红士,解伏菊.1987年大兴安岭特大火灾后北坡森林景观生态恢复评价.生态学报,2005,25(11):3098-3106.
[22]王绪高,李秀珍,贺红士.1987年大兴安岭特大火灾后不同管理措施对落叶松林的长期影响.应用生态学报,2008,19(4):915-921.
[23]公霞,常禹,布仁仓,李秀珍,徐崇刚,于庆和.呼中林业局森林采伐方式对森林景观格局的长期影响.生态学杂志,2006,25(7):805-812.
[24]周宇飞,贺红士,布仁仓,金龙如,李秀珍.不同针阔树种造林比例下小兴安岭森林景观的动态模拟.应用生态学报,2008,19(8):1775-1781.
[25]郭锐,布仁仓,胡远满,常禹,贺红士,刘晓梅,张志全.气候变化条件下小兴安岭林区森林采伐面积模拟.应用生态学报,2010,(7):1681-1688.
[26]陈宏伟,胡远满,常禹,布仁仓,李月辉,刘淼.呼中林区不同森林采伐方式对林火的长期影响模拟.北京林业大学学报,2011,33(5):13-19.
[27]刘信中,王琅.江西省庐山自然保护区生物多样性考察与研究.北京:科学出版社,2010:3-293.
[28]黄瑞采,戴朱恒,陈邦本,陆宝树.庐山区土壤的特征.土壤学报,1957,5(2):117-135.
[29]江西森林编委会.江西森林.北京:中国林业出版社,1986.
[30]徐崇刚,胡远满,常禹,李秀珍,布仁仓,贺红士,冷文芳.像元尺度上不确定性对空间景观直观模型模拟的影响.生态学报,2004,24(9):1938-1949.
[31]郭志华,卓正大.庐山常绿阔叶、落叶阔叶混交林的植物区系地理研究.热带地理,1996,16(1):72-81.
[32]张伊.江西省志——动植物志.北京:中央党校出版社,1994:45-57.
[33]杨建国.日本柳杉在我国的地理分布和环境的研究.江西林业科技,1980,(2):11-14.
[34]彭少麟,方炜.鼎湖山植被演替过程中椎栗和荷木种群的动态.植物生态学报,1995,19(4):311-318.
[35]彭少麟.鼎湖山人工马尾松第1代与自然更新代生长动态比较.应用生态学报,1995,6(1):11-13.
[37]林协,洪昌端,斯金平,曾逢良,纪品芬.日本花柏、日本扁柏生长发育规律的初步研究.林业科技通讯,1987,(12):10-13.
[38]罗世家,易咏梅,邹惠渝,梁师文,吴峰,吴旺杰.黄山松种子发芽率及活力测定.湖北林业科技,2001,(1):9-11.
[39]陈龙池,汪思龙,陈楚莹.杉木人工林衰退机理探讨.应用生态学报,2004,15(10):1953-1957.
[40]邬建国.景观生态学——格局、过程、尺度与等级 (第二版).北京:高等教育出版社,2007:106-115.
[42]徐崇刚,胡远满,姜艳,常禹,李秀珍,布仁仓,贺红士.空间直观景观模型的验证方法.生态学杂志,2003,22(6):127-131.
[43]王绪高,李秀珍,贺红士.大兴安岭森林景观在不同火干扰及人工更新下的演替动态.北京林业大学学报,2006,28(1):14-22.
[44]万慧霖,冯宗炜,庞宏东.庐山外来植物物种.生态学报,2008,28(1):103-110.
[46]郑勇奇,张川红.外来树种生物入侵研究现状与进展.林业科学,2006,42(11):114-122.
[47]申卫军,邬建国,林永标,任海,李勤奋.空间粒度变化对景观格局分析的影响.生态学报,2003,23(12):2506-2519.
