喀斯特峰丛洼地土壤剖面微生物特性对植被和坡位的响应
2013-12-16冯书珍苏以荣秦新民葛云辉何寻阳
冯书珍,苏以荣,秦新民,肖 伟,葛云辉,何寻阳,*
(1.中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,长沙 410125;2.广西师范大学生命科学学院,桂林 541004;3.湖南农业大学,长沙 410125;4.中国科学院环江喀斯特生态系统观测研究站,环江 547100)
西南喀斯特地区是一种受地质背景制约的脆弱生态环境,巨大的人口压力下,高强度的人为活动导致土地质量的下降与石漠化。该地区的退化土地生态恢复重建,维系着我国长江、珠江中下游地区的生态安全。近二十年来,通过实行退耕还林、封山育林、生态移民等措施,石漠化治理工作取得了一定成效,但石漠化快速扩展的总体趋势并没有得到有效的遏止[1],其原因除了生态恢复与农村脱贫致富结合不够外,更重要的原因是对该区生态恢复的内在机制认识不足:只注重地上植被的种植与培育,忽视了对生态系统功能维持和提升的土壤微生物的重要作用。
土壤微生物在养分循环与平衡、土壤理化性质的改善中起着重要作用[2-3],对环境变化敏感,其中,微生物呼吸强度是衡量土壤微生物总的活性指标[4],而微生物量的任何变化,都会影响土壤养分的循环和有效性[5],能够较早地指示生态系统功能的变化。土壤微生物受土壤理化性状、土壤生态系统内生物因素、不同经营措施等的影响[6]。有研究显示地上植被和地下微生物间有正负反馈的互作机制[7];坡位对土壤养分的剖面分布有着重要影响[8-9],进而影响土壤微生物的生长;栖居在深层的土壤微生物群落受深层土壤环境的制约,但对其随深度变化的主要影响因子看法不一。以往对土壤微生物的研究主要集中于表层土壤,人们对退化喀斯特生境恢复过程中土壤剖面微生物随植被恢复、坡位等的变化系统研究缺乏。
因此,本研究通过选取喀斯特峰丛洼地3种典型植被类型(草丛、灌丛、原生林:中坡位)、3种坡位(原生林:上、中和下坡位)的剖面土壤作为研究对象,研究土壤微生物量碳、氮、微生物熵、基础呼吸与代谢熵的剖面分异特征,阐明土壤剖面微生物特性对植被恢复与立地条件的响应规律,揭示影响土壤微生物特性变化的主要因子,以期深化对喀斯特峰丛洼地土壤剖面微生物及其功能的认识,为区域的可持续发展和生态恢复措施的制订提供理论依据。
1 研究地区与研究方法
1.1 研究区自然概况
本研究的研究区选为广西环江毛南族自治县下南乡西南部的古周示范区(24°50′N,107°55′E)以及木论国家自然保护区(25°06′—25°12′N,107°53′—108°05′E),同属于典型喀斯特峰丛洼地景观以及亚热带季风气候区。古周示范区最低点海拔为376 m,与最高点海拔相差440 m;多年平均气温16.5—20.5℃,多年平均降雨量为1389.1 mm;1a内日照时数以2月份最低,7月份最高,日照率以3月份最低,9月份最高;研究区的土壤主要为碳酸盐岩发育的石灰岩,自然植被以草丛和灌木群落为主,森林覆盖率仅为13%。木论国家自然保护区与古周研究区相距30 km,地形、气候条件与古周研究区大致相同,海拔400—1000 m,年平均气温15.0—18.7℃,极端高温36℃,极端低温-5℃;年均降雨量1530—1820 mm,林内相对湿度一般在80%—90%,夏季可接近饱和;林区土壤类型简单,主要为石灰土和零星分布的硅质土,均属非地带性土壤。样地的基本信息与优势种如表1。

表1 样地基本信息及植物优势物种Table 1 Site descriptions and dominant floral species in the studied communities
1.2 土壤样品采集与处理
2008年12月至2009年1月,选取草丛、灌丛(古周示范区)、原生林(木论自然保护区)从山顶到山脚各3条样带(1条主样带和2条辅助样带)。选取中坡位各3个大样方,分析土壤剖面微生物对不同植被类型的响应;同时,为考虑坡位的影响,在几无人为干扰的原生林3条样带上选取上、下坡位各3个大样方。样方面积为20 m×30 m,土样按土壤发生层采集淋溶层(A层,0—10 cm)、过渡层(AB层,20—30 cm,草丛和灌丛;30—50 cm,原生林)、淀积层(B层,70—100 cm)样品,共45个样品。环刀法测定各层土壤容重。
四分法取约500 g新鲜土壤置于灭菌后的自封袋中,封口后于随身携带的冰盒带回,在实验室去除土壤中可见的动植物残体,过2 mm筛,一份保存在4℃冰箱中供土壤微生物指标分析;另一份自然风干保存用于土壤理化性质分析。供试土壤的基本理化性质如表2。
1.3 样品分析方法
土壤微生物量碳(SMBC)采用氯仿熏蒸浸提-有机碳自动分析仪器法(Phoenix 8000),土壤微生物量氮(SMBN)采用氯仿熏蒸浸提-氮素流动注射分析仪器法(Fiastar 5000)测定[10]。
土壤基础呼吸(SBR)采用碱液吸收-TOC仪(Phoenix 8000)测定法,对龙健等的方法[11]稍作改进:将供试土壤20 g置于密闭1 L广口瓶中(内置一盛有1 mol/L NaOH溶液20 mL小广口瓶),广口瓶底部有10 mL的去离子水,以维持瓶内的空气饱和湿度,于28℃的恒温箱中培养24 h,于TOC仪上检测并计算土壤呼吸放出CO2的量,同时设空白处理作为对照。

1.4 数据处理与统计分析
土壤微生物量碳:

式中,Ec=熏蒸土壤提取的有机碳-未熏蒸土壤提取的有机碳;kEC为转换系数,取值0.45。
土壤微生物量氮:

式中,EN=熏蒸土壤提取的全氮-未熏蒸土壤提取的全氮;kEN为转换系数,取值0.45。

用SPSS 16.0和Excel 2007做统计分析和制图。以植被类型(草丛、灌丛和原生林)和土层深度以及坡位(上、中、下坡位)和土层深度作为处理因子分别对土壤微生物特性进行双因素方差分析,处理间的多重比较用S-N-K法。
2 结果与分析
2.1 植被类型和坡位对土壤剖面微生物特性的影响
2.1.1 土壤微生物量碳及碳熵的剖面分异特征
喀斯特峰丛洼地同一植被类型,SMBC随土层深度的增加而减少;同一坡位,原生林SMBC均在A层、AB层、B 层((1216.57±859.05)mg/kg、(1098.59±48.81)mg/kg、(531.60±19.67)mg/kg)表现最高,依次为:PF>S>T,灌丛SMBC在B层(160.62±16.65)mg/kg—A层(639.76±54.01)mg/kg之间;草丛SMBC的3个剖面均为最低,依次为A层((367.12±59.05)mg/kg)>AB层((134.41±48.81)mg/kg)>B层((56.66±15.00)mg/kg)。原生林3种坡位(上、中、下坡位),A层SMBC在下坡位表现最高,均与中、上坡位呈现显著差异;AB层SMBC则在下坡位最低,中、上坡位间无显著差异;B层上、下坡位无显著差异,中坡位最高(图1)。
草丛qMBC在0.96%—2.50%之间;灌丛qMBC在0.99%—2.77%之间;原生林qMBC在1.40%—4.67%之间。qMBC在3种植被类型间差异不同,整体表现为原生林最高,并随土层深度的增加而减小;原生林A层下坡位/中坡位>上坡位,AB层无显著差异,B层则呈现中坡位>下坡位/上坡位的趋势(图1右)。
2.1.2 土壤微生物量氮及氮熵的剖面分异特征
喀斯特峰丛洼地不同植被类型下SMBN与SMBC变化趋势类似,由草丛、灌丛、原生林呈现上升趋势(P<0.05);同一植被类型,SMBN均随土层深度的增加而降低:草丛SMBN含量介于B层(18.06±0.52)mg/kg—A层(48.60±3.16)mg/kg;灌丛SMBN含量介于B层 (18.60±0.82)mg/kg—A层(65.23±3.73)mg/kg之间;原生林均在A层、AB层、B层表现最高,依次为(194.42±2.31)mg/kg、(54.43±3.67)mg/kg、(31.81±5.87)mg/kg。A层B层SMBN在不同坡位间呈现下坡位>中坡位/上坡位的变化,AB层SMBN则在3种坡位间差异显著(图2)。
在草、灌丛阶段qMBN随土层深度的增加无显著变化;A层qMBN随植被恢复依次增加,AB层、B层表现为PF>T/S;原生林3种土层深度下,下坡位的qMBN均显著低于中坡位/上坡位(图2)。
2.1.3 土壤基础呼吸与代谢熵的剖面分异特征
3种植被类型,SBR均表现为:PF>S>T,并随土层深度的加深而逐渐减小;原生林3种坡位(上、中、下坡位)均表现为A层>AB层>B层,其中,A层土壤受坡位影响显著,表现为下坡位>中坡位>上坡位;AB层、B层土壤上、下坡位间无显著差异(图3)。
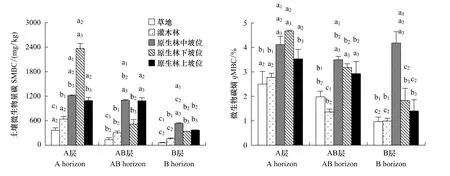
图1 土壤微生物量碳及微生物碳熵的剖面分异特征*Fig.1 The profile distribution on soil MBC and the ratio of MBC to SOC*

图2 土壤微生物量氮及微生物氮熵的剖面分异特征Fig.2 The profile distribution on soil MBN and the ratio of MBN to soil total nitrogen
qCO2在3种植被类型间差异显著:T>PF>S,在灌丛最低,草丛最高,表明土壤微生物对土壤碳的利用效率灌丛较高,其次为原生林,草丛较低。同一植被类型随土层深度的增加而减少,坡位则对qCO2无显著影响(图3)。
2.1.4 植被类型、坡位和土层深度对土壤微生物特性的影响
在喀斯特峰丛洼地,植被类型、土层深度对SMBC、SMBN与SBR的影响都达到了极显著水平(P<0.01);坡位对土壤微生物指标影响不一,对qCO2无显著影响(P>0.05);除坡位和土层深度的交互作用对SMBC和SMBN有极显著影响外,其余两两交互作用对土壤微生物指标影响不显著(表3)。
2.2 土壤微生物特性之间及与土壤养分的相关性
土壤养分与土壤微生物特性的相关性分析结果表明 (表4),SMBC、SMBN、qMBC和SBR均与有机碳、全氮、碱解氮呈显著正相关(P<0.01),SMBC、SMBN和SBR与全磷也呈现正相关关系(P<0.01),而qCO2则与土壤基本养分无明显的相关性;土壤微生物特性之间,SMBC和SMBN,qMBC和qMBN,SBR分别与SMBC、SMBN呈显著正相关关系(P<0.01);土壤微生物特性均与pH无显著相关关系。
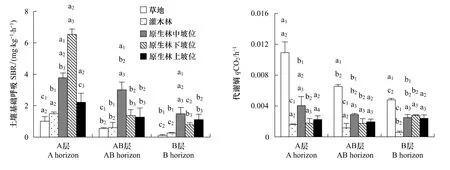
图3 土壤基础呼吸与代谢熵的剖面分异特征Fig.3 The profile distribution on soil basic respiration and microbial metabolic quotient

表3 不同因素对土壤微生物特性的方差分析结果Table 3 Results of different factors and their interactions on soil microbial properties by Two-way ANOVA

表4 土壤微生物特性之间及其与土壤基本理化性质的相关系数(n=45)Table 4 Correlation coefficients among soil microbial and basic physicochemical properties(n=45)
3 讨论
3.1 土壤剖面微生物量碳氮及微生物熵对植被类型及坡位的响应
不同植被类型,微生物量碳(SMBC)和微生物氮(SMBN)均随着土层深度的加深而减少,这与微生物利用底物的量直接相关(表4),研究表明SMBC的高低主要受土壤中有机碳源的制约[12],并与随土壤剖面的加深土壤活性碳库的减少有关[13]。随植被恢复,3种土层深度下,SMBC由草丛、灌丛、原生林依次上升,SMBN也表现出类似的趋势(A层:PF>S>T;AB/B层:PF>S/T),这主要归因于其地上部分生物量的差异使输入到土壤中的有机碳量明显不同[14],植被的种类不同其枯落物的质量也不同,造成进入土壤的微生物可利用底物的质和量的变化,从而影响土壤微生物量的变化[15-16]。一般认为,小尺度的坡位因子能在一定程度上引起坡面土壤理化性质的变化(表1),从而导致不同坡位的土壤微生物量分异和微生物区系的改变[7,17]。在喀斯特地区,土壤微生物量对土层深度的响应比坡位更为强烈(表3),人类干扰较少的林地坡地土壤侵蚀沿中上坡向坡脚逐渐加剧,下坡位较中/上坡位其侵蚀程度较为严重[18],而土壤养分在各土地利用类型剖面中遵循表聚特征[19],致使原生林A层SMBC表现为由下坡位到上坡位递减的趋势,而AB层、B层SMBC则表现为下坡位<中坡位/上坡位。
土壤微生物碳熵(qMBC)的变化可以反映土壤有机碳的变化[20],在指示土壤过程或者健康状况时,被用作土壤性质以及健康的指标[21]。研究发现,植被类型、坡位均对qMBC有显著影响(表3),qMBC在3种植被类型下原生林表现为最高,表明土壤中的有机碳在植被恢复过程中得以累积;在3种坡位下(原生林:上、中、下坡位),下坡位显著高于上坡位,表明下坡位表层土壤沉淀富集,结构性好,土壤肥力水平较中、上坡位高。土壤微生物氮熵(qMBN)反映微生物对全氮的利用效率,3种土层深度下,qMBN均表现为下坡位<中/上坡位,这可能是由于下坡位是坡面养分的汇集处[7],其中碱解氮显著高于中、上坡位(表1),反映近期内维持土壤微生物生长利用所需氮素供应丰富,下坡位土壤微生物对全氮的利用效率较中/上坡位低,这和qMBN与土壤全氮含量显著正相关(表4)相悖,推测其可能的原因是土壤qMBN不仅与土壤氮素水平相关,而且与土壤微生物群落结构利用氮素能力和微生物可利用的氮素形态[22]等土壤微环境相关。
3.2 土壤基础呼吸及代谢熵对植被类型及坡位的响应
土壤基础呼吸(SBR)是表征土壤质量和肥力的重要生物学指标,尤其是反映了土壤的生物活性和土壤物质代谢的强度[23-24],是非常敏感的指标,对于土壤环境甚至微环境都有敏感的反应[25]。植被恢复过程中SBR均随着土层的加深而减少,这可能是由于A层土壤接收较多的植物凋落物和有相对较多的根系分布,有机质含量高,有较充分的营养促进微生物的生长[26],而随着土壤剖面的加深,有机质等条件变差(表1),不利于土壤微生物的生长与活动,基础呼吸减弱。本研究表明,不管是A层土壤还是B层土壤,SBR均随植被的恢复而增大,差异达到显著水平(P<0.05),与土壤微生物量的变化一致,说明土壤有机残体的分解速度和强度:PF>S>T,这是由于植物较高的多样性会产生较高的植物生物量[27],从而导致其进入土壤系统的有机质成分较多,同时,较高的植物物种丰度及多样性也增加了微生物利用资源的异质性,有利于有机质的分解及转化,使得土壤保肥、供肥能力增加(表1)。SBR对坡位的响应,A层土壤较AB层、B层更为敏感,呈现由下坡位至上坡位依次递减的趋势,这种变化的主要原因可能是降雨侵蚀过程中的再分配造成不同坡位土壤养分含量的差异[6-7]。
代谢熵(qCO2)是土壤基础呼吸与土壤微生物量的比值,是衡量土壤微生物对土壤碳的利用效率高低的指标[28],也代表了微生物群落维持力大小和对有机质的利用效率[29]。研究结果表明,qCO2表现为灌丛最低,草丛最高,在植被恢复后期表现出增加的趋势,说明灌丛的碳利用率最高,草丛的碳利用率最低,这与杨刚研究结果一致[31]。这可能是由于在草丛、灌丛阶段土壤微生物的qCO2与土壤的熟化程度呈负相关[30],到原生林阶段,土壤微生物群落及功能发生改变,从而引起了qCO2值的增大[31],其内在的机制可能是原生林阶段受到更为明显的矿质养分P的胁迫,在这种条件下,微生物必须从维持生长和繁殖的能量中分流出一部分去补偿由于胁迫所需要付出的额外能量[13,32]。本研究发现qCO2与坡位、土壤基本理化性质无明显相关性(表3,表4),但坡位对SBR、SMBC、SMBN的变化均有显著影响,说明在反映喀斯特峰丛洼地坡位对土壤质量变化时,qCO2比SBR、SMBC、SMBN更加稳定,指示qCO2在评价土壤质量变化随坡位的改变存在局限性。
4 结论
(1)土壤微生物量碳、基础呼吸随植被的恢复而增大,同样适用于深层土壤,坡位则对土壤剖面微生物特性影响不一致:代谢熵受坡位影响不显著。
(2)深层土壤微生物受深层土壤环境的制约,A层各微生物特性对植被类型及坡位的响应比AB层B层的更为敏感。
(3)喀斯特峰丛洼地中,土壤有机碳、全氮、碱解氮与土壤微生物量碳、氮、微生物熵及代谢熵关系密切,因此,调控土壤氮素营养与土壤有机质对于维持土壤微生物功能同等重要。
本研究并没有涉及土壤微生物多样性的剖面分异特性对植被类型及坡位的响应,而这对于土壤碳氮周转及土壤肥力维持同样重要,因此未来将纳入土壤微生物多样性的研究,为进一步寻求反映土壤退化或土壤肥力保持的土壤生物学指标提供证据,并为指导喀斯特石漠化治理提供科学依据。
致谢:感谢中国科学院亚热带农业生态研究所陈彩艳研究员对写作的帮助。
[1] Yuan D X.Global view on Karst rock desertification and integrating control measures and experiences of China.Pratacultural Science,2008,25(9):19-25.
[2] Doran J W,Zeiss M R.Soil health and sustainability:managing the biotic component of soil quality.Applied Soil Ecology,2000,15(1):3-11.
[3] Wei Y,Zhang J C,Yu Y C,Yu L F.Changes of soil basal respiration and qCO2along successional processes of degraded Karst vegetation.Chinese Journal of Soil Science,2010,41(4):797-801.
[4] Huang Y M,An S S,Xue H.Responses of soil microbial biomass C and N and respiratory quotient(qCO2)to revegetation on the Loess Hilly-Gully region.Acta Ecologica Sinica,2009,29(6):2811-2818.
[5] Kuzyakov Y,Subbotina I,Chen H Q,Bogomolova I,Xu X L.Black carbon decomposition and incorporation into soil microbial biomass estimated by14C labeling.Soil Biology and Biochemistry,2009,41(2):210-219.
[6] Hu Y L,Wang S L,Yan S K.Research advances on the factors influencing the activity and community structure of soil microorganism.Chinese Journal of Soil Science,2006,37(1):170-176.
[7] Wsrdle D A,Bardgett R D,Klironomos J N,Setälä H,van der Putten W H,Wall D H.Ecological linkages between aboveground and Belowground biota.Science,2004,304(5677):1629-1633.
[8] Gao X S,Deng L J,Zhang S R.Soil physical properties and nutrient properties under different utilization styles and slope position.Journal of Soil and Water Conservation,2005,19(2):53-56,60-60,79-79.
[9] Harris R F,Chesters G,Allen O N.Dynamics of soil aggregation.Advance in Agronomy,1996,18:107-169.
[10] Wu J,Jogensen R G,Pommerening B,Chaussod R,Brookes P C.Measurement of soil microbial C by fumigation:An automated procedure.Soil Biology and Biochemistry,1990,22(8):1167-1169.
[11] Long J,Deng Q Q,Jiang X R,Liu F.Effects of different de-farming and reforestation patterns on changes of soil fertility quality in karst region of southwestern China.Chinese Journal of Applied Ecology,2005,16(7):1279-1284.
[12] Witter E,Mårtensson A M,Garcia F V.Size of the soil microbial biomass in a long-term field experiment as affected by different N-fertilizers and organic manures.Soil Biology and Biochemistry,1993,25(6):659-669.
[13] Agnelli A,Ascher J,Corti G,Ceccherini M T,Nannipieri P,Pietramellara G.Distribution of microbial communities in a forest soil profile investigated by microbial biomass,soil respiration and DGGE of total and extracellular DNA.Soil Biology and Biochemistry,2004,36(5):859-868.
[14] Pan F J,Zhang W,Wang K L,He X Y,Liang S C,Wei G F.Litter C:N:P ecological stoichiometry character of plant communities in typical Karst Peak-Cluster Depression.Acta Ecologica Sinica,2011,31(2):335-343.
[15] Xu H Q,Zhang J E,Feng L F,Quan G M,Mao D J,Qin Z.Effects of different land use patterns on microbial biomass carbon and nitrogen in Guangdong Province.Acta Ecologica Sinica,2009,29(8):4112-4119.
[16] Jobbágy E G,Jackson R B.The vertical distribution of soil organic carbon and its relation to climate and vegetation.Ecological Application,2000,10(2):423-436.
[17] Huang Y M,An S S,Liu L J,Xue H.Soil basal respiration response to grass vegetation restoration and it′s affecting factors in the Loess Hilly-Gully Region.Chinese Journal of Eco-Agriculture,2009,17(5):862-869.
[18] Feng T,Chen H S,Zhang W,Nie Y P,Wang K L.137Cs profile distribution character and its implication for soil erosion on Karst slopes of Northwest Guangxi.Chinese Journal of Applied Ecology,2011,22(3):593-599.
[19] Wei X P,Yuan D X,Xie X Y.Study on soil erosion and loss on slope in karst mountain valley area of Chongqing valley with137Cs and soil nutrient elements.Journal of Soil and Water Conservation,2010,24(6):16-23.
[20] Liu S L,Su Y R,Huang D Y,Xiao H A,Wu J S.Response of Cmic-to-Corgto land use and fertilization in subtropical region of China.Scientia Agricultura Sinica,2006,39(7):1411-1418.
[21] He X Y,Wang K L,Chen Z H,Yang G,Chen H S.Responses of soil microbial characters to farm land-use types in peak-cluster depression of karst region.Chinese Journal of Soil Science,2008,39(3):409-513.
[22] Qiu S J,Peng P Q,Liu Q,Rong X M.Soil microbial biomass nitrogen and its role in nitrogen cycling.Chinese Journal of Ecology,2006,25(4):443-448.
[23] Cookson W R,Abaye D A,Marschner P,Murphy D V,Stockdale E A,Goulding K W T.The contribution of soil organic matter fractions to carbon and nitrogen mineralization and microbial community size and structure.Soil Biology and Biochemistry,2005,37(9):1726-1737.
[24] Ming X,Ye Q.Soil-surface CO2efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California.Global Change Biology,2001,7(6):667-677.
[25] Fang C,Moncrieff J B,Gholz H L,Clark K L.Soil CO2efflux and its spatial variation in a Florida slash pine plantation.Plant and Soil,1998,205(2):135-146.
[26] Zhou Y,Xu X G,Wang F,Ruan H H,Wang J S,Fang Y H,Wu Y Y,Xu Z K.Soil microbial biomass,respiration,and metabolic quotient along an altitudinal gradient in Wuyi Mountain of southeastern China.Chinese Journal of Ecology,2009,28(2):265-269.
[27] Cardinale B J,Wright J P,Cadotte M W,Carroll I T,Hector A,Srivastava D S,Loreau M,Weis J J.Impacts of plant diversity on biomass production increase through time because of species complementarity.Proceedings of the National Academy of Sciences of the United States of America,2007,104(46):18123-18128.
[28] Silvana A P F,Wagner B,Cerri C C.Effect of sewage sludge on microbial biomass,basal respiration,metabolic quotient and soil enzymatic activity.Applied Soil Ecology,2005,30(1):65-77.
[29] Grayston S J,Griffith G S,Mawdsley J L,Campbell C D,Bardgett R D.Accounting for variability in soil microbial communities of temperate upland grassland ecosystems.Soil Biology and Biochemistry,2001,33(4/5):533-551.
[30] Anderson T H.Microbial eco-physiological indicators to asses soil quality.Agriculture,Ecosystems and Environment,2003,98(1/3):285-293.
[31] Yang G,He X Y,Wang K L,Huang J S,Chen Z H,Li Y Z,Ai M R.Effects of vegetation types on soil micro-biomass carbon,nitrogen and soil respiration.Chinese Journal of Soil Science,2008,39(1):189-191.
[32] Odum E P.Trends expected in stressed ecosystems.Bioscience,1985,35(7):419-422.
参考文献:
[1] 袁道先.岩溶石漠化问题的全球视野和我国的治理对策与经验.草业科学,2008,25(9):19-25.
[3] 魏媛,张金池,余元春,喻理飞.退化喀斯特植被恢复过程中土壤基础呼吸及代谢熵的变化.土壤通报,2010,41(4):797-801.
[4] 黄懿梅,安韶山,薛虹.黄土丘陵区草地土壤微生物C、N及呼吸熵对植被恢复的响应.生态学报,2009,29(6):2811-2818.
[6] 胡亚林,汪思龙,颜绍馗.影响土壤微生物活性与群落结构因素研究进展.土壤通报.2006,37(1):170-176.
[8] 高雪松,邓良基,张世熔.不同利用方式与坡位土壤物理性质及养分特征分析.水土保持学报,2005,19(2):53-56,60-60,79-79.
[11] 龙健,邓启琼,江新荣,刘方.西南喀斯特地区退耕还林(草)模式对土壤肥力质量演变的影响.应用生态学报,2005,16(7):1279-1284.
[14] 潘复静,张伟,王克林,何寻阳,梁士楚,韦国富.典型喀斯特峰丛洼地植被群落凋落物C:N:P生态化学计量特征.生态学报,2011,31(2):335-343.
[15] 徐华勤,章家恩,冯丽芳,全国明,毛丹娟,秦钟.广东省不同土地利用方式对土壤微生物量碳氮的影响.生态学报,2009,29(8):4112-4119.
[17] 黄懿梅,安韶山,刘连杰,薛虹.黄土丘陵区土壤基础呼吸对草地植被恢复的响应及其影响因素.中国生态农业学报.2009,17(5):862-869.
[18] 冯腾,陈洪松,张伟,聂云鹏,王克林.桂西北喀斯特坡地土壤137Cs的剖面分布特征及其指示意义.应用生态学报,2011,22(3):593-599.
[19] 魏兴平,袁道先,谢世友.运用137Cs与土壤营养元素探讨重庆岩溶槽谷区山坡土壤的流失和漏失.水土保持学报,2010,24(6):16-23
[20] 刘守龙,苏以荣,黄道友,肖和艾,吴金水.微生物商对亚热带地区土地利用及施肥制度的响应.中国农业科学,2006,39(7):1411-1418.
[21] 何寻阳,王克林,陈志辉,杨刚,陈洪松.岩溶洼地土壤微生物指标对不同土地利用方式的响应.土壤通报,2008,39(3):409-513.
[22] 仇少君,彭佩钦,刘强,荣湘民.土壤微生物生物量氮及其在氮素循环中作用.生态学杂志,2006,25(4):443-448.
[26] 周焱,徐宪根,王丰,阮宏华,汪家社,方燕鸿,吴焰玉,徐自坤.武夷山不同海拔梯度土壤微生物生物量、微生物呼吸及其商值(qMB,qCO2).生态学杂志,2009,28(2):265-269.
[31] 杨刚,何寻阳,王克林,黄继山,陈志辉,李有志,艾美荣.不同植被类型对土壤微生物量碳氮及土壤呼吸的影响.土壤通报,2008,39(1):189-191.
