川西米亚罗林区不同海拔岷江冷杉生长对气候变化的响应
2013-12-16王晓春张远东刘世荣
徐 宁,王晓春,张远东,刘世荣
(1.东北林业大学生态研究中心,哈尔滨 150040;2.中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态环境重点实验室,北京 100091)
全球气候变化会影响地球生态系统的稳定性和持续性,并将威胁人类的生存空间和可持续发展,因此分析气候变化对生态系统的影响具有十分重要的意义[1]。森林是陆地生态系统的主体,且对气候变化敏感,尤其是亚高山森林森林生态系统响应更为敏感[2-4]。近年来,国内外学者在不同亚高山地区开展了树木年轮与气候关系研究,特别在对气候变化响应和历史气候重建方面进行了较多工作[5-10]。气候变化对不同树种的影响不同,例如,温度可能极大地影响某一树种的生长,却对另一树种影响不大[11-14];即使同一树种,在不同气候条件下甚至不同环境梯度上,其生长与气候的关系也存在差异[15-18]。我国学者近年来也开展了不同海拔树木生长与气候关系的研究,但研究结果之间存在一定的差异。天山中段雪岭云杉(Picea schrenkiana)树木生长主要受前一年7、8月高温和当年4、5月降水不足导致的干旱影响,只有在接近气候林线处,低温的限制作用才表现出来[19];山西芦芽山中低海拔华北落叶松(Larix principis-rupprechtii)的树木径向生长受到土壤干旱的限制,而高海拔华北落叶松受到低温的限制。随海拔升高,树木径向生长与生长季降水的关系从正相关逆转为负相关[20];贡嘎山不同海拔云冷杉树木生长与平均温度均呈正相关[21]等。这些研究表明在干旱地区,亚高山森林树木生长可能主要受降水限制;而在湿润地区,亚高山森林树木生长可能主要受温度限制。但也有研究发现了高山林线树木生长对气候变化并不敏感的现象,与高山林线主要受生长季温度限制的传统认识明显不同[22]。在川西半湿润地区,树木生长主要受哪些气候因子的影响?海拔及其所引起的水热变化对树木生长与气候的关系有哪些影响?除了气候对树木生长起着一定影响作用,树龄是否会对树木生长产生一定影响?这些问题仍需深入研究。
青藏高原东缘亚高山森林广布,主要树种包括云杉、冷杉、落叶松等,同一区域的不同海拔或立地,其生长限制因子会发生变化[1]。青藏高原由东南至西北降雨量逐渐减少,由湿润区变化至干旱区。川西米亚罗林区位于川西邛崃山区,属于半湿润地区,开展该地区不同海拔树木生长与气候关系研究,是对不同气候和不同海拔条件下树木生长-气候关系分析的有益补充。本文通过建立米亚罗地区岷江冷杉(Abies faxoniana)树轮年表,探讨海拔和树龄对岷江冷杉生长与气候间关系的影响,为进一步研究半湿润地区生态系统对气候变化的响应奠定基础。
1 研究区域概况与研究方法
1.1 研究区域概况
研究区位于川西米亚罗林区(31°24'—31°55'N,102°35'—103°4'E,海拔 2200—5500 m),属青藏高原东缘褶皱带外缘部分。地形以石质山地为主,坡度多在35°以上,属高山气候。以海拔2760 m的米亚罗镇为例,全年降水量700—1400 mm,年蒸发量1000—1900 mm。夏季温凉多雨,冬季干燥寒冷,最冷月为1月(-8℃),最热为7月(12.6℃),≥10℃的年积温为1200—1400℃。米亚罗林区植被垂直成带明显,其类型和生境随海拔及坡向而分异。原生森林分布于海拔2400—4200 m之间,以亚高山暗针叶林为主,优势树种为岷江冷杉和紫果云杉(Picea purpurea)。
1.2 研究方法
1.2.1 样品采集与年表建立
于2011年7月份,选择在受人为活动影响较小的区域,分高、中、低海拔在3个地点采样(表1)。基于国际树木年轮数据库标准,选择生长良好的岷江冷杉进行树芯取样。用内径为5.3 mm的生长锥在胸高处(1.3 m)钻取树芯,尽量取到髓心。将取到的样本放入塑料管内,并进行编号,每株树木采集1个年轮样芯。样本带回实验室后,剔除断裂或未钻取到髓芯等不适合本研究的部分样芯,最后共保留132根实验样芯,并全部交叉定年。

表1 川西米亚罗林区高、中、低3个海拔树轮采样点信息Table 1 Information for three sampling sites(High-altitude,Middle-altitude,Low-altitude)at Miyaluo,western Sichuan
年轮样品预处理基本按照Fritts[23]阐述的方法进行,经晾干、固定、打磨后,在双筒显微镜下进行目视交叉定年。然后用VELMAX年轮测量仪测量年轮宽度,该系统精度为0.001 mm。测量后的树轮样本序列用COFECHA程序[24]对定年和测量结果进行检验,并消除定年和宽度测量过程中出现的误差。经过交叉定年的年轮序列,利用ARSTAN程序采用负指数或线性函数进行去趋势和标准化[25],其目的是去除低频变化趋势,降低因年龄生长速率不同而带来的影响;用双权重平均法进行年轮曲线的标准化,最终建立3个海拔岷江冷杉年表。从年表统计特征可知,差值年表的各项统计特征均高于标准年表,例如:高海拔差值年表中,样本代表性为0.94,大于标准年表中的0.90;差值年表中的第一特征根方差解释量为34.77%,大于标准年表中的26.19%等,因此选用差值年表进行年轮-气候关系分析。
1.2.2 气象资料与数据分析
由于川西米亚罗地区缺乏长期气候数据,气候资料选用都江堰气象站(103°40'E,31°01'N,海拔698 m)1954—2011年的气候数据,其位于采样点东南部约100 km处(图1)。研究中主要用月均气温、月最低温、月最高温、月总降水量、帕尔默干旱指数(palmer drought severity index,PDSI)[26]5 项气象指标。由于树木生长不仅与当年的气候条件有关,也受上年气候因子的影响。因此,选取上年6—12月份和当年1—9月份的各月的气象因子以及春、夏、秋、冬4个季气候因子的平均值与岷江冷杉年表进行分析[27]。所取岷江冷杉最大年龄在170a左右,因此将3个海拔的样芯按树龄分成大于85a和小于85a两组,相同海拔不同年龄段的两组年表分别与气候因子进行相关分析。年表与气候要素之间的相关分析用SPSS 19.0软件来计算。

图1 四川都江堰市国家站月平均气温与月总降水量的变化Fig.1 Monthly mean temperature and total precipitation at Dujiangyan,Sichuan,China
3 结果与分析
3.1 年表特征
在米亚罗地区所取得的3个差值年表中,它们的年表统计特征值(表2)在海拔梯度上表现出了一定的规律性,平均相关系数在0.32—0.42之间,平均敏感度在0.17—0.21之间,其中低海拔最高,而且均表现出从低到高递减趋势。3个海拔岷江冷杉年表的平均敏感度、信噪比、样本代表性都相对较好,表明其适合进行年轮气候学分析。
从图2可以看出,3个岷江冷杉差值年表整体变化趋势基本相似。通过对3个年表的相关分析得出,低海拔和中海拔的年表相关性最强(0.55),中海拔和高海拔的相关性最低(0.32)的结论,但它们的相关都达到了95%的显著水平。这表明米亚罗地区不同海拔岷江冷杉的生长具有区域一致性。

表2 川西米亚罗林区岷江冷杉3个海拔差值年表的主要统计特征Table 2 Major statistic characteristics for residual chronologies of Abies faxoniana from three sampling sites at Miyaluo,western Sichuan
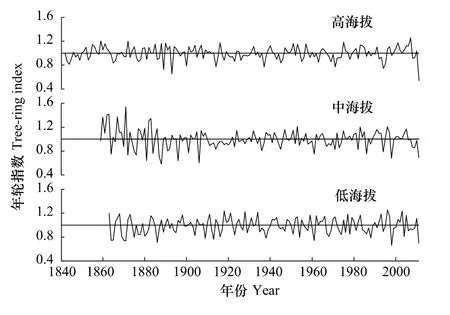
图2 川西米亚罗林区高、中、低3个海拔岷江冷杉的差值年表Fig.2 Residual chronologies of Abies faxoniana at three sampling sites at Miyaluo,western Sichuan
3.2 不同海拔岷江冷杉径向生长与气候因子的相关性
由表3可见,中低海拔2个采样点表现出相对一致的相关性,即都与当年春季和年均最高温呈极显著负相关关系,其中低海拔对春季最高温响应更显著,而中海拔与年均最高温响应最明显。同时,中海拔还与年均平均温度呈显著负相关,低海拔与当年春季平均温度呈显著负相关。高海拔岷江冷杉生长与前一年冬季最低温呈显著正相关,与均温、降水、最高温响应不显著。
从图3可以看出,低海拔年表与当年4、5月均温以及5、6月最高温显著负相关。中海拔年表与当年4月均温和4、5月份最高温以及前年8月最低温呈显著负相关。高海拔与2月份最低温的正相关显著,与1月均温正相关关系显著,另外还与7月降水量显著负相关。干旱指数方面,中低海拔均有较为明显的正相关关系,分别是低海拔在当年3、4、5月份、中海拔在当年4、5月份达到正相关显著,且低海拔显著性明显高于中海拔。高海拔年表与干旱指数的相关不显著。

表3 不同海拔岷江冷杉年表与季节气候变量的相关关系Table 3 Correlation coefficients between chronologies of Abies faxoniana and seasonal climatic variables
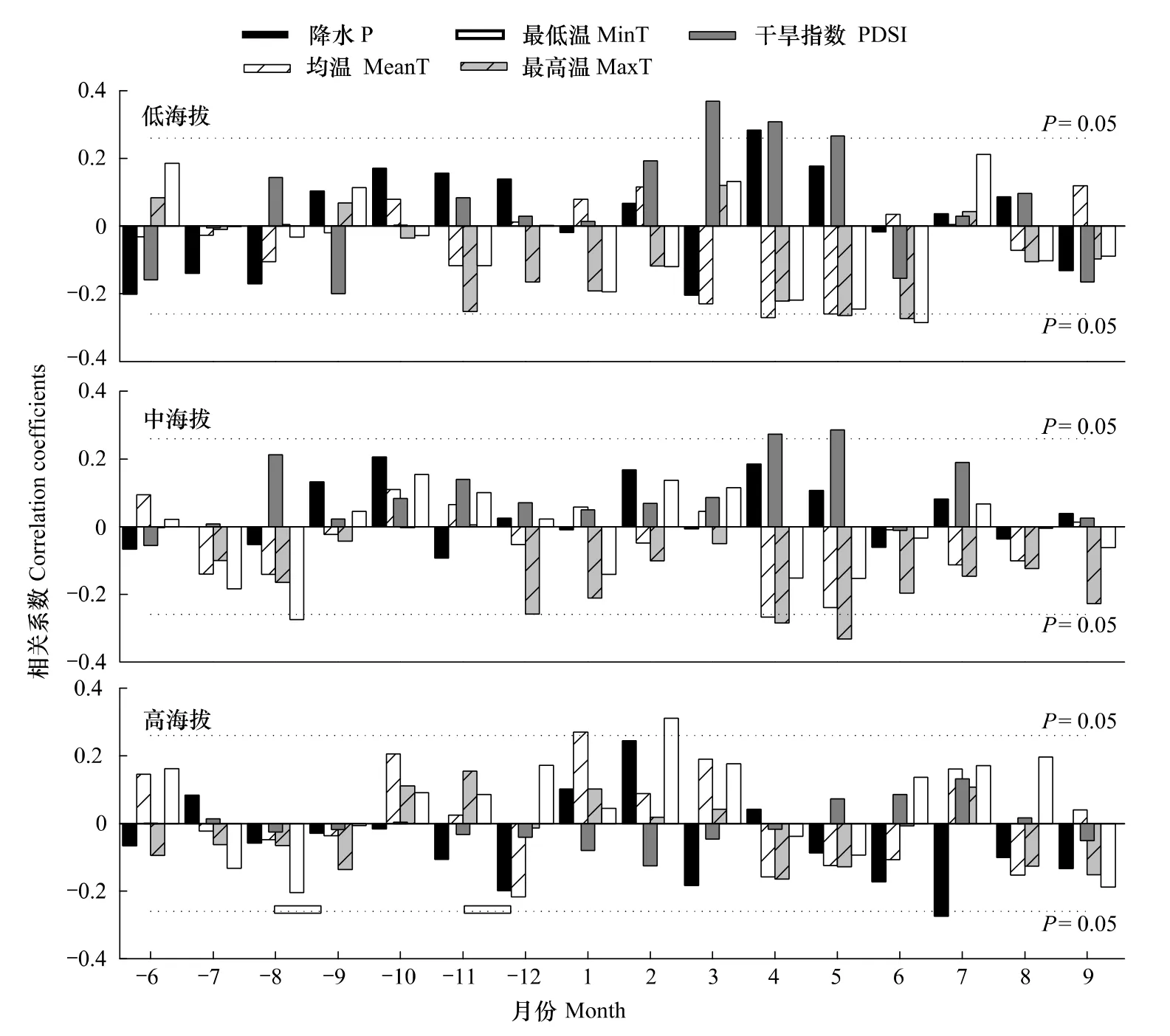
图3 不同海拔岷江冷杉年表与月气候数据的相关关系Fig.3 Correlation coefficients between chronologies of Abies faxoniana at different elevations and monthly climatic data
3.3 不同树龄岷江冷杉径向生长与气候因子的相关性
由表4可见,在高中低3个海拔,大龄岷江冷杉对气候因子的响应相对较为明显。中海拔大龄树木对春季和年均温度、最高温呈显著负相关,高海拔大龄树仅与夏季最低温呈显著正相关。3个海拔中,仅中海拔小龄树与年均最高温呈显著负相关。
由图4可以看出,总体上大龄树木比同海拔小龄树木生长对气候变化的响应较为敏感。低海拔小龄树与7月最低温呈显著正相关;中海拔小龄树与前1年8月的最低温和当年4、5月份的最高温显著负相关;高海拔小龄岷江冷杉与9月平均温度显著负相关,与2月最低温显著正相关。与小龄树木相比,低海拔的大龄树还与当年5、6月份的均温、最高温显著负相关;中海拔的大龄树木还与前一年12月最高温度呈显著负相关;高海拔的大龄树木还与2月降水量显著正相关。高中2个海拔的小龄树与低海拔的树木年龄相近,这3个响应结果比较发现,随海拔升高对前一年9月和当年4月降水量的正相关有显著性降低趋势,而2月份降水量有显著性升高趋势;5月平均温度的负相关有显著性降低趋势,而9月平均温度的负相关有显著性升高趋势;4、5月份最高温的负相关有降低趋势;2月最低温有正相关性升高趋势。
4 讨论与结论
4.1 海拔对岷江冷杉年轮-气候关系的影响
树木生长与春季均温和最高温的相关性规律比较明显,随海拔降低,负相关呈增加趋势(表3)。海拔梯度相当于一个天然增温平台,随着海拔降低气温升高,导致土壤水分的蒸散以及植物体的蒸腾作用加速,水分条件极有可能变成限制因素,在容易出现干旱胁迫的春季表现最为明显。春季是树木由休眠进入生长季的关键时期,在当地春季气温高、太阳辐射强和空气相对湿度低的气候条件下,树木失水较多,气孔部分关闭引起光合作用能力下降,在生理上起到生长抑制作用[28]。另外,春末夏初的高温往往会造成地面水分蒸发旺盛,使树木生长初期出现生理缺水现象,从而生长变缓[29]。
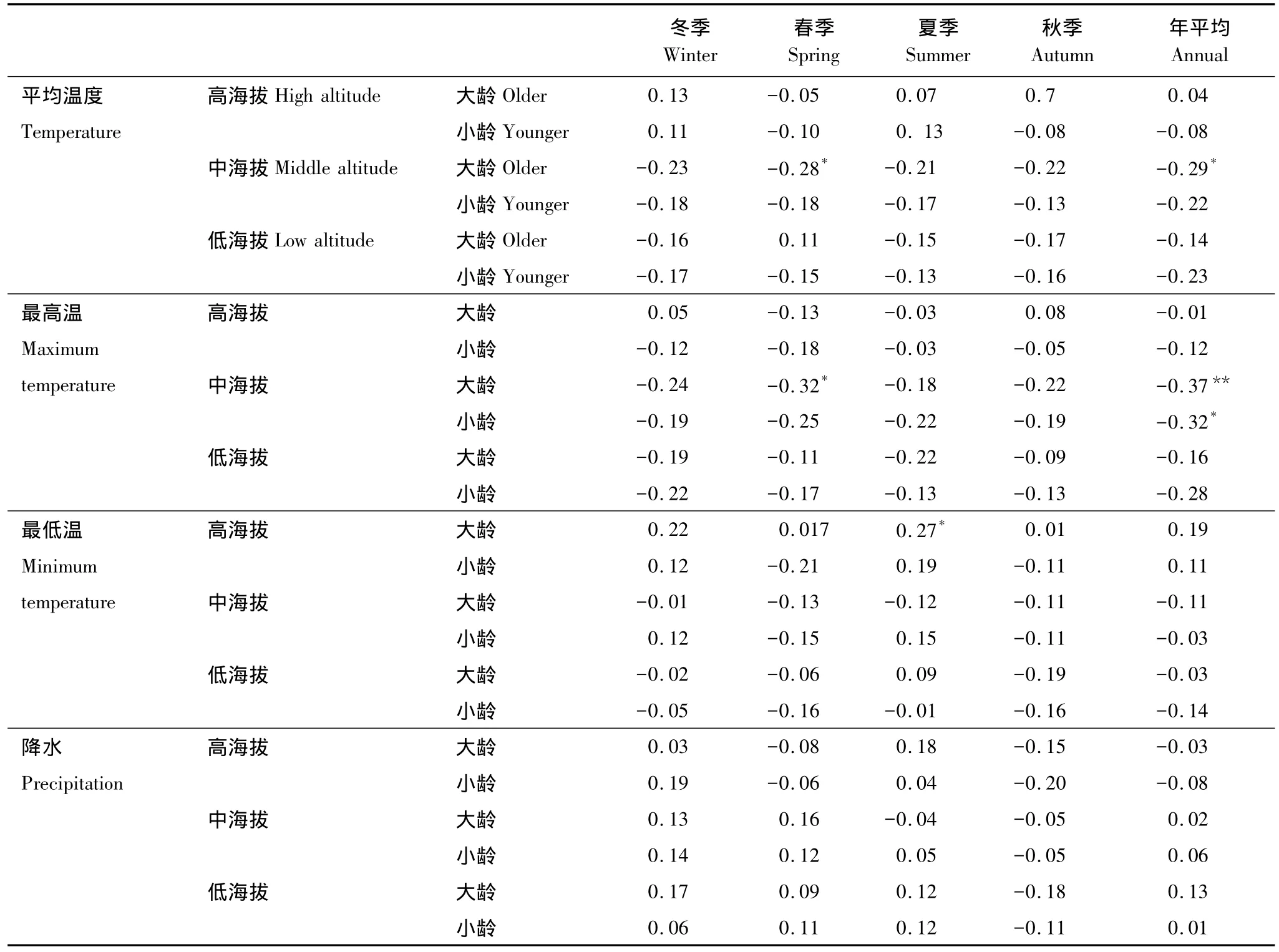
表4 不同海拔不同年龄段岷江冷杉与季节气候因子的相关关系Table 4 Correlation coefficients between younger or older chronologies of Abies faxoniana at different elevations and seasonal meteorological data
中低海拔岷江冷杉生长与5月均温和4、5月最高温负相关显著,表明春季高温抑制了树木生长。这与彭剑峰等[30]在阿尼玛卿山对青海云杉的研究结果相吻合,在国外也有类似结论[31]。春季均温、最高温与岷江冷杉生长显著负相关(表3),表明春季温度增高,同时春季降水偏少,使干旱程度加剧。PDSI在一定程度上了整合了降水量、大气温度和土壤水分蒸发等几种因素的综合影响[26],中低海拔岷江冷杉年表与4、5月份PDSI之间存在显著正相关(图3),低海拔还与3月份PDSI有显著正相关关系,同时伴随着或均温、或最低温、或最高温的显著负相关,这表明了中低海拔春季干旱胁迫的存在。米亚罗7、8月降水量处于一年中降水量的峰值,高海拔岷江冷杉年表与7月降水量显著负相关,同时,温度表现为一定程度的正相关,这是由于温度和降水之间的负相关关系所决定的。由于降水量的明显增加,导致温度降低,温度的降低会导致光合作用速率的降低,产生对树木生长所必须的有机物质量减少,进而抑制树木的生长[32]。可以认为,在研究区内中部海拔的7、8月份树木生长主要受由于高温引起的干旱影响。
李宗善等[33]对川西卧龙地区岷江冷杉的研究表明,晚冬至早春(1月至4月)温度对树木生长有着明显的促进作用,而春末5月份的高温对于树木生长有限制性影响。卧龙林区位于米亚罗林区的东南部,靠近华西雨屏区,年均降水量约710 mm,略高于米亚罗林区。这说明在青藏高原东缘,由东南至西北,随着降水量的减少,温度对生长的影响表现出了阈值效应,既当增温幅度在一定范围内时,对蒸发量影响不大,表现为促进生长;当增温幅度超过某一临界值时,蒸发量大于降水,从而抑制了生长[20]。升温和降水量减少相关联,则使得这种抑制作用表现得更为显著。
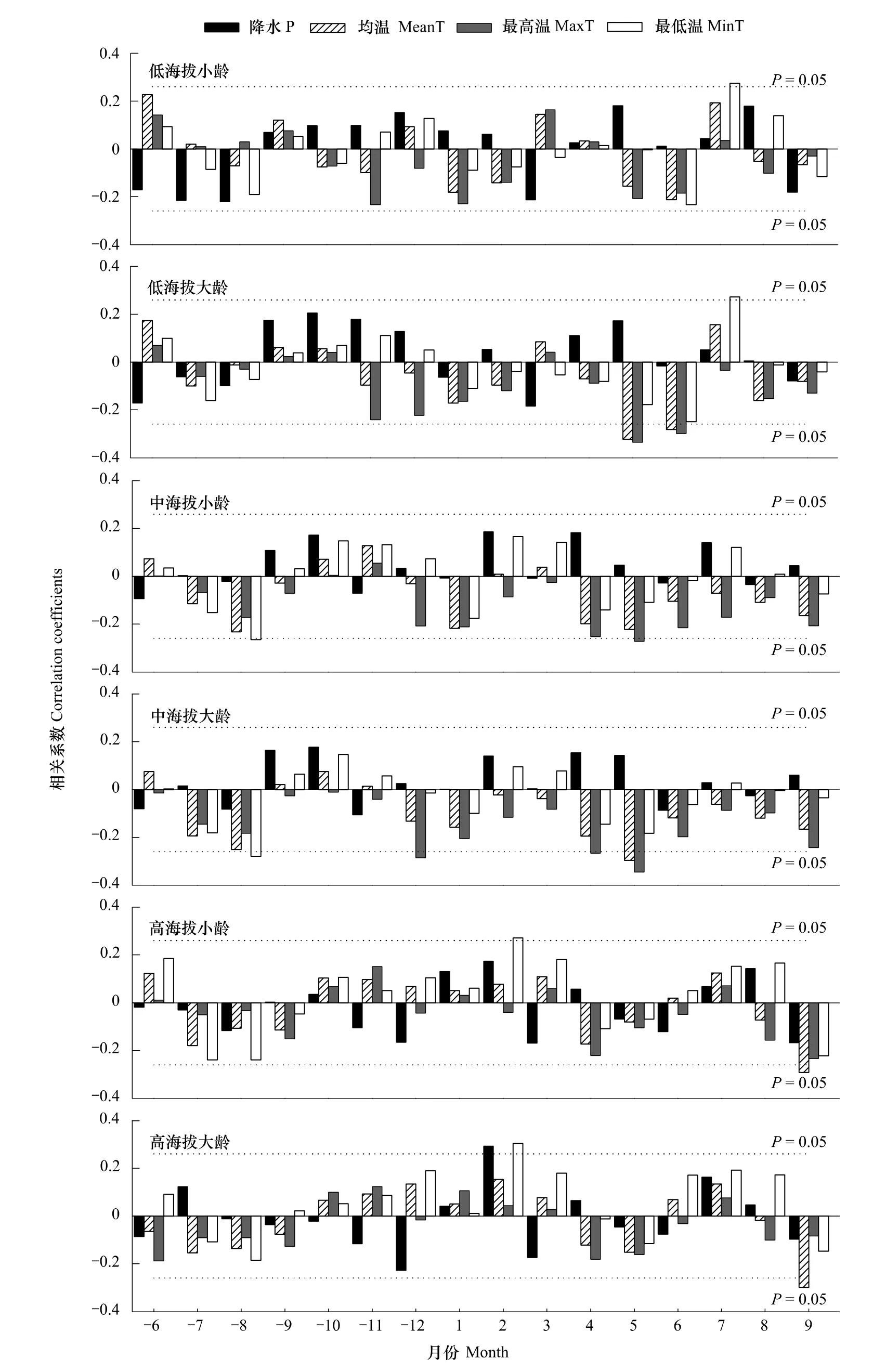
图4 不同海拔不同年龄段岷江冷杉年表与月气候因子的相关关系Fig.4 Correlation coefficients between monthly climatic data and older or younger chronologies of Abies faxoniana at different elevations
降水量对于岷江冷杉生长的影响,随海拔降低有从负相关向正相关变化的趋势。其中当年4月,高、中、低3个海拔的降水都与生长呈正相关,随海拔降低相关性递增,低海拔达到显著。这说明在一定的高度范围内,随海拔高度的上升降水逐步增加,越来越能满足岷江冷杉的生长需求,降水对树木生长的限制作用逐步减弱。因此,在没有其他限制因子出现的情况下,降水与树木生长的相关关系随海拔降低有从负相关向正相关变化的趋势。
在高海拔岷江冷杉生长主要受温度限制,与冬季最低温度呈显著正相关。这说明,树木在冬季停止生长,呼吸作用是树木主要的生理活动,温度越低,树木需要的保证正常生理代谢的能量越多;相反,温度越高,储存有机物的消耗越少,余下的有机物越多,更有利于后一年树木的生长[34];另一方面,冬季低温如果达到一定的低值,极有可能对树体产生损伤,进而抑制树木的生长[35]。而在中低海拔,生长与春季温度呈显著负相关,而与4月降水呈较高正相关,说明春季干旱对生长的影响更为显著。
4.2 树龄对岷江冷杉径向生长-气候关系的影响
总体上大龄树木的生长比同海拔小龄树木的生长对气候的响应更为敏感。这是由于大龄树木处于林冠层,受气候因子的影响更为强烈;而小龄树木更易受林内气候和竞争的影响,另外,幼龄树木本身生长旺盛,对外界环境胁迫的抵抗力也强,具体表现在生长方面,相对不易受外界干扰。王晓明等在长白山的研究表明,大龄红松对气候响应的敏感性更高,原因是生理和环境因素的共同作用[36]。Ryan和Yoder[37]曾指出,随着年龄增加,树体高度逐渐接近其最大值,树体内水分的运输成为一个主要限制因子,而水分胁迫能够导致气孔过早关闭,影响树体与外界的气体交换,进而限制树木生长。另外,树体内水分含量的变化,还可导致其他一系列复杂的生理活动的改变,比如有研究显示与树龄相关的水分胁迫甚至可以导致树木生理状况在遗传水平发生改变[38]。综合来看,大龄和小龄冷杉生理机制差异的复杂性影响了二者对气候的响应。除此之外,冷杉生长过程中局部环境的压力发挥着不可忽视的作用。大龄岷江冷杉在研究区域内与云杉及一些阔叶树种共同形成茂密的林冠层,而小龄冷杉往往位于林冠层之下,因此,小龄岷江冷杉的生长面临着更为严峻的竞争形势。有研究表明天然林内胸径为25—30 cm的树木个体面临的种间竞争强度远远大于胸径40—45 cm的个体[39]。小龄树木在与高大树体的竞争中处于劣势,且小龄树木之间为争夺有利的生长条件开展的竞争也是无可避免的,激烈的竞争削弱了小龄树木年轮生长对气候的响应能力,而大龄树木因为已经进入主林层,竞争趋于缓和,所以其径向生长表现出更高的气候敏感性。
[1] Wu X D.Tree-Ring and Climate.Beijing:China Meteorological Press,1990:125-125,149-149,135-135.
[2] Wu X D,Shao X M.A Preliminary analysis on response of tree-ring density to climate in the Qinling Mountains of China.Chinese Journal of Applied Ecology,1994,5(2):253-256.
[3] Wang X C,Song L P.Climate-tree growth relationships of Pinus sylvestris var.mongolica in the northern Daxing'an Mountains,China.Chinese Journal of Plant Ecology,2011,35(3):294-302.
[4] Fritts HC.Reconstruction large scale climate patterns from tree-ring data,Tuson,USA.Arizona:The Arizona University Press,1991.
[5] Shao X M,Fan J M.Past climate on west Sichuan plateau as reconstructed from ring-width of dragon spruce.Quaternary Sciences,1999,(1):82-89.
[6] Wu P,Wang L L,Shao X M.Reconstruction of summer temperature from maximum latewood density of Pinus densata in west china.Acta Geographica Sinica,2005,60,998-1006.
[7] Shao X M,Huang L,Liu H B,Liang E Y,Fang X Q,Wang L L.Reconstruction of precipitation variation from tree rings in recent 1000 years in Delingha,Qinghai.Science in China Series D-Earth Sciences,2005:48,939-949.
[8] Fan Z X,Bräuning A,Bao Y,Cao K F.Tree ring density based summer temperature reconstruction for the central Hengduan Mountains in southern China.Global and Planetary Change,2008,65:1-11.
[9] Liang E Y,Shao X M,Qin N S.Tree-ring based summer temperature reconstruction for the source region of the Yangtze River on the Tibetan Plateau.Global and Planetary Change,2008,61:313-320.
[10] Liu Y,An Z S,Linderholm H W,Chen D L,Song H M,Cai Q F,Sun J Y,Tian H.Annual temperatures during the last 2485 years in the mideastern Tibetan Plateau inferred from tree rings.Science in China Series D-Earth Sciences,2009,52:348-359.
[11] D'Arrigo R D,Jacoby G C.Secular trends in high northern latitude temperature reconstructions based on tree rings.Climate Change,1993,25:163-177.
[12] Lan T,Xia B and He S A.Tree ring analysis on relation of Pinus massoniana growth to climate factors.Chinese Journal of Applied Ecology,Oct.1994,5(4):422-424.
[13] Peng J F,Gou X H,Chen F H.Characteristics of ring-width chronologies of Picea crassifolia and their responses to climate at different elevations in the Anyemaqen Mountains.Acta Ecologica Sinica,2007,27(8):3268-3276.
[14] Gou X H,Chen F H,Yang M X.Analysis of the tree-ring width chronology of Qilian Mountains at different elevation.Acta Ecologica Sinica,2004,24(1):172-176.
[15] Bonan G B,Sirois L.Air temperature,tree growth,and the northern and southern range limits to Picea mariana.Journal of Vegetation Science,1992,3:495-506.
[16] Brooks J R,Flanagan L B,Ehleringer J R.Responses of boreal conifers to climate fluctuations:indications from tree-ring widths and carbon isotope analyses.Canadian Journal of Forestry Research,1998,28:524-533.
[17] Yu D P,Wang S Z,Tang L N.Relationship between tree-ring chronology of Larix olgensis in Changbai Mountains and the climate change.Chinese Journal of Applied Ecology,2005,16(1):14-20.
[18] Sun F,Zhong Z C.Relationship between tree-ring growth of Gordonia acuminate and climatic factors in Mt.Jinyun.Chinese Journal of Applied Ecology,1999,10(2):151-154.
[19] Guo Y Y,Liu H Y,Ren J,Zhan X F,Cao S P.Response of tree growth to vertical climate gradient in the middle section of the Tianshan Mountains.Quaternary Sciences,2007,27(3):322-331.
[20] Zhang W T,Jiang Y.Larix principis-rupprechtii's radial growth-climate relationship at different elevations in Luya Mountains.Journal of Beijing Normal University(Natural Science),2011,47(3):304-309.
[21] Duan J P,Wang L L,Xu Y.Response of tree-ring width to climate change at different elevations on the east slope of Gongga Mountains.2010,29(11):1940-1949.
[22] Zeng L B,Wang X P,Chang J F,L X,Wu Y L,Yin W L.Alpine timberline ecotone tree growth in relation to climatic variability for Picea crassifolia forests in the middle Qilian Mountains,northwestern China.Journal of Beijing Forestry University,2012,34(5):50-56.
[23] Fritts H G.Tree ring and climate.London:Academy Press,1976:1-567.
[24] Holmes R L.Computer-assisted quality control in tree-ring dating and measurement.Tree-Ring Bulletin,1983,43,69-78.
[25] Cook E R,Holmes R L.User's manual for program ARSTAN.In:Holmes R L,Adams R K,Fritts H C eds.Tree-Ring Chronologies of Western North America:California,Eastern Oregon and Northern Great Basin.Chronology Series VI.Laboratory of Tree-Ring Research,University of Arizona.1986,50-60.
[26] LIU G S,G A H,A S Q,L W W.Research progress in Palmer drought severity index and it's application.Journal of Natural Disaster,2004,13(4):21-27.
[27] Li X,et al.Age-dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau.Tree Physiology,2012,doi:10.1093/treephys/tps113.
[28] Li G Q,Bai F,Sang W G.Different responses of radial growth to climate warming in Pinus koraiensis and Picea jezoensis var.komarovii at their upper elevational limits in Changbai Mountain,China.Chinese Journal of Plant Ecology,2011,35(5):500-511.
[29] Cao Y Z.Plant physiology.Lanzhou:Lanzhou University Press.
[30] Peng J F,Gou X H,Chen F H.Characteristics of ring-width chronologies of Picea crassifolia and their responses to climate at different elevations in the Anyemaqen Mountains.Acta Ecologica Sinica,2007,27(8):3268-3276.
[31] Jacoby G C,D'Arrigo R D.Tree-ring width and density evidence of climatic and potential forest change in Alaska.Global Biogeochemical Cycles,1995,9,227-234.
[32] Liang E Y,Shao X M,Hu Y X.Dendro climatic evaluation of climate-growth relationships of Meyer spruce(Picea meyeri)on a sandy substrate in semi-arid grassland,North China.Trees,2001,15:230-235
[33] Li Z S,Liu G H,Fu B J.Evaluation of temporal stability in tree growth-climate response in Wolong National Natural Reserve,western Sichuan,China.Chinese Journal of Plant Ecology,2010,34(9):1045-1057.
[34] Cullen L E,Palmer J G,Duncan R P.Climate change and tree-ring relationships of Nothofagus menziesii tree-line forests.Canada Journal Forest Research,2001,31:1981-1991.
[35] Pan H L,Li M H,Cai X H,W J,Du Z,Liu X L.Responses of growth and ecophsiology of plants to altitude.Ecology and Environmental Sciences,2009,18(2):722-730.
[36] Wang X M,Zhao X H,Gao L S,Jiang Q B.Age-dependent growth responses of Pinus koraiensis to climate in the north slope of Changbai Mountain,North-Eastern China.Acta Ecologica Sinica,2011,31(21):6378-6387.
[37] Ryan M G,Yoder B J.Hydraulic limits to tree height and tree growth.Bio Science,1997,47:235-242.
[38] Day M E,Greenwood M S,Diaz-Sala C.Age and size related trends in woody plant shoot development:regulatory pathways and evidence for genetic control.Tree Physiology,2002,22(8):507-513.
[39] Liu H R,Li F R.Relationship between intra specific and inter specific competitions of natural Pinus Koraiensis forests.Bulletin of Botanical.Research,2010,30(4):479-484.
参考文献:
[1] 吴祥定.树木年轮与气候变化.1990,北京:气象出版社.
[2] 吴祥定,邵雪梅.中国秦岭地区树木年轮密度对气候响应的初步分析.应用气象学报,1994,5(2):253-256.
[3] 王晓春,宋来萍,张远东.大兴安岭北部樟子松树木生长与气候因子的关系.植物生态学报,2011,35(3):294-302.
[5] 邵雪梅,范金梅.树轮宽资料所指示的川西过去气候变化.第四纪研究,1999,19(1):81-89.
[6] 吴普,王丽丽,邵雪梅.采用高山松最大密度重建川西高原近百年夏季气温.地理学报,2005,60(6),998-1006.
[12] 兰涛,夏冰,贺善安.马尾松的生长与气候关系的年轮分析.应用生态学报,1994,5(4):422-424.
[14] 勾晓华,陈发虎,杨梅学,彭剑峰,强维亚,陈拓.祁连山中部地区树轮宽度年表特征随海拔高度的变化.生态学报,2004,24(1):172-176.
[17] 于大炮,王顺忠,唐立娜,代力民,王庆礼,王绍先.长白山北坡落叶松年轮年表及其与气候变化的关系.应用生态学报,2005,16(1):14-20.
[18] 孙凡,钟章成.缙云山四川大头茶树木年轮生长动态与气候因子关系的研究.应用生态学报,1999,10(2):151-154.
[19] 郭允允,刘鸿雁,任佶,战晓峰,曹善平.天山中段树木生长对气候垂直梯度的响应.第四纪研究,2007,27(3):322-331.
[20] 张文涛,江源,董满宇,杨艳刚,杨浩春.芦芽山不同海拔华北落叶松径向生长与气候因子关系的研究.北京师范大学学报(自然科学版),2011,47(3):304-309.
[21] 段建平,王丽丽,徐岩,孙毓,陈津.贡嘎山东坡不同海拔高度树轮宽度对气候变化的响应.地理研究,2010,29(11):1940-1949.
[28] 李广起,白帆,桑卫国等.长白山红松和鱼鳞云杉在分布上限的径向生长对气候变暖的不同响应.植物生态学报,2011,35(5):500-511.
[29] 曹仪植.植物生理学,1998,兰州:兰州大学出版社.
[30] 彭剑峰,勾晓华,陈发虎,刘普幸,张永,方克艳.阿尼玛卿山地不同海拔青海云杉(Picea crassifolia)树轮生长特性及其对气候的响应.生态学报,2007,27(8):3268-3276.
[32] 梁尔源,邵雪梅,胡玉熹,林金星.内蒙古草原沙地白扦年轮生长指数的变异内蒙古草原沙地白扦年轮生长指数的变异.植物生态学报,2001,25(2):190-194.
[33] 李宗善,刘国华,傅伯杰,张齐兵,胡婵娟,罗淑政.川西卧龙国家级自然保护区树木生长对气候响应的时间稳定性评估.植物生态学报,2010,34(9):1045-1057.
[35] 潘红丽,李迈和,蔡小虎等.海拔梯度上的植物生长与生理生态特性.生态环境学报,2009,18(2):722-730.
[36] 王晓明,赵秀海,高露双,姜庆彪.长白山北坡不同年龄红松年表及其对气候的响应.生态学报,2011,31(21):6378-6387.
[39] 刘红润,李凤日.红松天然林种内和种间竞争关系的研究.植物研究,2010,30(4):479-484.
