九连山保护区米槠群落主要种群空间分布格局研究
2013-11-18徐国良汤正华
徐国良, 汤正华, 廖 娟
(1.九连山自然保护区管理局, 江西 龙南 341700; 2.龙南县九连山林场, 江西 龙南 341700; 3.龙南县龙南镇第二小学, 江西 龙南 341700)
九连山保护区米槠群落主要种群空间分布格局研究
徐国良1, 汤正华2, 廖 娟3
(1.九连山自然保护区管理局, 江西 龙南 341700; 2.龙南县九连山林场, 江西 龙南 341700; 3.龙南县龙南镇第二小学, 江西 龙南 341700)
采用方差/均值的t检验法以及负二项式分布指数K、聚块性指数IP、丛生指标IMorisitar指数对九连山保护区米槠群落的主要乔木种群的分布格局进行了研究。结果表明:该群落重要值排在前17位的种群中,8种为聚集分布,9种为随机分布;该群落中,竞争力较强的种群主要表现为聚集分布,竞争力较弱的种群主要表现为随机分布;种群的不同径级分布格局是,随着径级的增加,先表现为聚集程度的增加,当种群的径级增长到一定程度时又表现为聚集程度的减小;优势灌木的分布格局是,除狗骨柴与山血丹外,树参、草珊瑚、黄楠、鼠刺等4种优势灌木均表现为聚集分布;各优势种群的格局规模都表现为多种格局规模分布。
九连山自然保护区; 米槠群落; 主要种群; 分布格局
米槠(Castanopsiscarlesii) 林是我国东部亚热带湿润地区常绿阔叶林中重要群系之一[1]。相比壳斗科的其它种,米槠作为九连山自然保护区常绿阔叶林的主要建群种更为普遍。目前对米槠群落的研究多集中在群落组成、结构特征、植物区系成分等方面。我们对九连山米槠群落优势种群的空间分布特征进行了研究,以期为进一步研究该地区的常绿阔叶林生态系统提供参考。
1 研究区自然地理概况
九连山国家级自然保护区地处南岭山地东段,位于24°29′— 24°38′N,114°22′—114°31′E。境内海拔自 300m 至主峰黄牛石 1 430 m,地势南高北低,属中亚热带常绿阔叶林南缘,为中亚热带与南亚热带过渡地带;其典型的地带性植被为常绿阔叶林,现保存有较大面积的原生性常绿阔叶林。九连山气候具有大陆性、海洋性,又受山地影响,其性质属中亚热带湿润山地气候,年平均气温 16.7℃,1 月平均气温 7.5℃,7月平均气温24.2℃,极端最低气温-7.4℃,极端最高气温37.0℃, 年均降水量2 070.4mm,年均相对湿度85%;基岩多为花岗岩﹑变质岩,成土母质主要有砂岩和砂页岩,山坡及沟谷中为残积物和坡积物所形成的黄棕壤及山地黄壤,pH值4.5~6.5[2]。
2 研究方法
2.1野外调查
对亚热带常绿阔叶林进行空间分布格局研究的取样以10m×10m为合适的样方大小,且样地面积应在1600m2上[3]。选取一处有代表性的米槠群落,设置17个10m×10m的调查样方。对样方内胸径(DBH)>2.5cm 的乔灌木作每木调查,测定个体总数、胸径、树高、枝下高、冠幅等。在样方的右上角设置 2m×2m的小样方,对小样方内胸径(DBH)<2.5cm的乔木幼苗和灌木统计种类、株数或丛数,测定幼苗高度,估计草本的多度等。样地设置地海拔560~580m,坡度10~40°,位于西南和东南两坡的夹沟之间。
2.2优势种群年龄结构及群落垂直结构的划分
将优势种群根据胸径大小划分为四级[4]:Ⅰ级幼苗,胸径<2.5cm;Ⅱ级幼树,胸径2.5~7.5cm ;Ⅲ级立木,胸径7.5~22.5cm ;Ⅳ级大树,胸径>22.5cm 。对胸径>2.5cm 的优势乔木种群进行垂直结构分层:乔木Ⅰ层,树高15m以上;乔木Ⅱ层,树高5~15m;乔木Ⅲ层,树高5m 以下[5]。
2.3分布格局测定
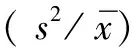
3 结果与分析
3.1主要乔木种群的垂直分层与重要值
对胸径>2.5cm 的乔木种群进行垂直结构分层与重要值计算,从表1中可以看出,米槠的重要值和相对显著度居本群落之首,说明米槠在本群落中的大树最多,在上层乔木中其数量远远多于其它种。可见,米槠是本群落的建群种。木荷在第Ⅱ乔木层中的数量最多,相对多度和频度最大,重要值位居第二,在群落中发挥着重要的作用。丝线吊芙蓉个体虽小,但其在下层乔木层中数量最多,对本群落的影响也较大。因此,本群落的准确命名应为米槠+木荷+丝线吊芙蓉群落。

表1 九连山米槠群落主要乔木种群的垂直分层及重要值Tab 1 TheverticaldelaminationandimportantvalueofmainarborpopulationsofCastanopsiscarlesiicommunityinJiulianshanNatureReserve树种[8]在各乔木层的数量ⅢⅡⅠ相对多度(%)频度(%)相对频度(%)相对显著度(%)重要值(%)重要值序(%)米槠(Castanopsiscarlesii)34583018 2094 128 5161 5488 251木荷(SchimasuperbaGardn)6578223 35100 009 0416 6549 042丝线吊芙蓉(Rhododendronwestlandii)6845018 2088 247 986 0532 233罗浮柿(Diospyrosmorrisiana)63306 2876 476 913 8717 064密花树(Rapaneameriifolia)231105 4852 944 790 8811 155鸭公树(Neolitseachuii)71403 3858 825 321 269 966南岭栲(Castanopsisfordii)111103 5452 944 791 409 737少叶黄杞(Engelhardtiafenzelii)12602 9052 944 790 588 278虎皮楠(Daphniphyllumoldhami)3901 9352 944 790 927 649细枝柃(Euryaloquaiana)6401 6147 064 260 276 1410黄樟(Cinnamomumporrectum)0600 9723 532 131 454 5511冬桃(Elaeocarpusduclouxii)0600 9723 532 130 793 8912杨桐(Adinandramillettii)3300 9729 412 660 243 8713鼠刺(Iteachinensis)3100 6423 532 130 092 8614多穗柯(Lithocarpuspolystarhyus)3100 6417 651 600 342 5815桃叶石楠(Photiniaprunifolia)0300 4817 651 600 292 3716华南桂(Cinnamomumjaponicam)0300 4817 651 600 232 3117 注:本群落另有29种乔木的重要值共计38%未列出
3.2主要乔木种群空间分布格局
对重要值排在前17位的种群进行分析,结果表明:方差/均值法和Morisita指标测得的结果相同,即有8个种是聚集分布,而丛生指数、聚块性指数、负二项式指数则反映了这些乔木种群聚集程度的差异。表2表明,本群落重要值排在前7位的优势种群除罗浮柿外,均为极显著的聚集分布,其中聚集程度最强的是米槠。重要值排在第7位以后的除黄樟外都是随机分布。由此可见,在本群落中,竞争力较强的种群主要表现为聚集分布,而竞争力较弱的种群为争取更大的空间主要表现为随机分布。
种群的聚集分布主要是由于生境的异质性或其本身的生物学特性引起的[9],比如米槠南岭栲是靠壳斗包着的坚果繁殖,果实较重,其传播距离较近而分布在母株周围。同样木荷与丝线吊芙蓉是塑果,塑果开裂后种子也大多聚集在母株周围。密花树、黄樟多聚集在调查样地的西南坡,坡度较陡,土层相对较薄,上层米槠、木荷等大乔木相对较少的地方,这可能与它们对光照的需求有关。少叶黄杞的果实为翅果,因其种子传播距离更远而呈随机分布。结合表3中各种群不同立木级的分布格局可知,还有一种可能,就是种群幼苗呈随机分布,随着林木的年龄增长与径级增大,对资源的需求量也增加,密度制约效应与种间竞争使其数量随着径级的增加而减少,最后呈集中分布。

表2 九连山自然保护区米槠林主要乔木种群的分布格局Tab 2 ThedistributionpatternofmainarborpopulationsofCastanopsiscarlesiicommunityinJiulianshanNatureRe⁃serve树种方差/均值value值t检验格局丛生指数平均拥挤度聚块性指数Morisita指数value值F检验格局负二项式指数米槠13 33834 897 C∗12 33818 6912 9422 84513 339 C∗0 515木荷4 63210 273 C∗3 63212 1611 4261 4044 636 C∗2 348丝线吊芙蓉4 0798 709 C∗3 0799 7261 4631 4404 079 C∗2 158罗浮柿1 6761 912C0 6762 9701 2951 2841 675C3 390密花树2 6254 596 C∗1 6253 6251 8124 5962 625 C∗1 231鸭公树2 2803 620 C∗1 2802 5152 0362 0242 279 C∗0 965南岭栲2 9715 575 C∗1 9713 2652 5232 5022 971 C∗0 656少叶黄杞1 3541 001R0 3541 4131 3341 3331 354R2 989虎皮楠1 0210 059R0 0210 7271 021 0301 021R33 200细枝柃1 0750 212R0 0750 6631 1281 1331 075R7 864黄樟1 7502 120C0 7501 1033 1253 4001 750C0 472冬桃1 3961 120R0 3960 7492 1222 2671 396R0 893杨桐1 0420 119R0 0420 3951 1191 1331 041R8 333鼠刺0 813-0 528R-0 1870 0480 2050 0000 813R-1 257多穗柯1 3450 975R0 3450 5802 4682 8331 344R0 681桃叶石楠0 875-0 353R-0 1250 0510 2900 0000 875R-1 408华南桂0 875-0 353R-0 1250 0510 2900 0000 875R-1 408 注:“C”为Clump,表示聚集分布;“R”为Random,表示随机分布;带“∗”者为极显著的聚集分布。下同。
3.3不同发育阶段优势种群的分布格局
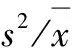
木荷在本群落中是衰退型种群,根据表1,其胸径>2.5cm的株数在本群落中最多。但随着群落郁闭度的增加,木荷幼苗的数量及分布频度已经变得较少了,在激烈的种内与种间竞争状态下,现有的Ⅱ级和Ⅲ级立木必然会不断疏散从而使整个种群的聚集程度不断变低,最终可能表现为随机分布。虽然丝线吊芙蓉的3个立木级均表现为极显著的聚集分布,但其幼苗的数量以及分布的频度也在逐渐的减少,幼苗只聚集在少数几个适合其生长的样方里,因此,随着演替的进行,其种群的聚集程度亦会慢慢降低。根据以上分析可以预测:在本群落的演替过程中,在竞争中占优势的种群可以较长期的保持聚集;在竞争中占劣势的种群因为其更新能力差,缺少幼苗补充,而大树为争取更大的生存空间,其种群聚集程度将逐渐变小。从表3还可以看出,在本群落中表现为增长型的种群还有鸭公树,表现为衰退型的还有密花树、罗浮柿等。
3.4灌木的分布格局
表4是6种优势灌木的分布格局。由表4可知,除狗骨柴与山血丹外,树参、草珊瑚、黄楠、鼠刺等4种优势灌木均表现为聚集分布。这是因为山血丹与狗骨柴为耐荫灌木[10],所以不必聚集在林窗下而呈随机分布。灌木的聚集分布也与其树形较矮,种子传播距离相对较近有关。表4中所列出的优势灌木相对优势乔木出现的频度较小也可以说明这一点。

表3 九连山自然保护区米槠林优势乔木种群各立木级的分布格局Tab 3 ThedistributionpatternofdominantarborpopulationswithdifferentsizeclassofCastanopsiscarlesiicommunityinJiulianshanNatureReserve树种立木级株数频度(%)方差/均值聚块性负二项式指数value值t检验格局指数Ⅰ级157100 01 0900 254R1 01095 718米槠Ⅱ级5270 61 3470 980R1 1130 567Ⅲ级3147 17 69118 900 C∗4 6680 273Ⅳ级2964 72 6214 585 C∗1 9501 052Ⅰ级1658 81 1250 353R1 1337 504Ⅱ级10294 14 2509 180 C∗1 5421 846木荷Ⅲ级4076 52 2813 618 C∗1 5441 836Ⅳ级317 70 875-0 353R0 290-1 408丝线Ⅰ级1229 42 4384 062 C∗3 0370 491吊芙蓉Ⅱ级7288 22 7905 056 C∗1 4232 366Ⅲ级4170 62 6464 649 C∗1 6821 465树名立木级株数频度(%)方差/均值聚块性负二项式指数value值t检验格局指数Ⅰ级1341 22 2133 426 C∗2 5880 632密花树Ⅱ级3152 92 4834 189 C∗1 8131 230Ⅲ级311 81 5851 652R4 3241 408Ⅰ级3388 21 4474 088 C∗1 2304 340鸭公树Ⅱ级1341 22 3733 883 C∗2 2490 555Ⅲ级829 41 3561 007R1 7561 314Ⅰ级1223 52 9695 562 C∗3 7890 358南岭栲Ⅱ级1647 12 0552 985 C∗2 3640 733Ⅲ级523 51 1770 500R1 6021 654Ⅰ级735 30 927-0 206R0 8235 657罗浮柿Ⅱ级1947 12 1113 138 C∗1 9941 006Ⅲ级1958 81 6641 876C1 5941 685 注:除米槠和木荷外,其它几种优势种群的Ⅳ级立木株数均为“0”。
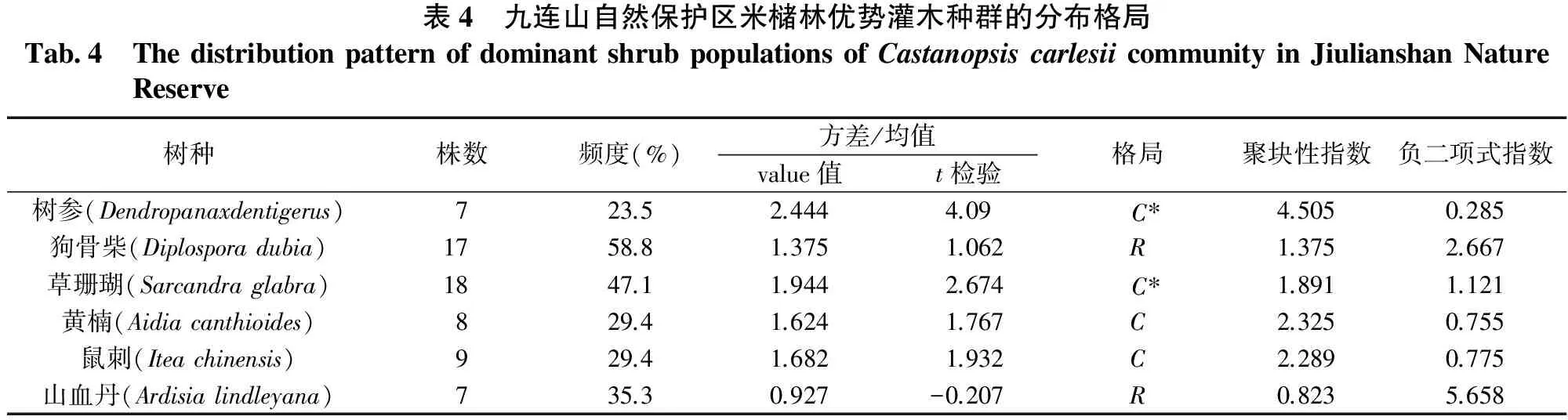
表4 九连山自然保护区米槠林优势灌木种群的分布格局Tab 4 ThedistributionpatternofdominantshrubpopulationsofCastanopsiscarlesiicommunityinJiulianshanNatureReserve树种株数频度(%)方差/均值格局聚块性指数负二项式指数value值t检验树参(Dendropanaxdentigerus)723.52.4444.09 C∗4.5050.285狗骨柴(Diplosporadubia)1758.81.3751.062R1.3752.667草珊瑚(Sarcandraglabra)1847.11.9442.674 C∗1.8911.121黄楠(Aidiacanthioides)829.41.6241.767C2.3250.755鼠刺(Iteachinensis)929.41.6821.932C2.2890.775山血丹(Ardisialindleyana)735.30.927-0.207R0.8235.658
3.5优势种群的格局规模
格局规模是指种的1个斑块和另1个斑块间隙之和的平均长度,等于1个斑块的中心到另1个相邻斑块中心的距离[11]。采用双项轨迹方差法[12]对种群的格局规模进行分析,结果如图1所示。各种群都表现为多种格局规模,这可能是各种群互相竞争的结果或者是受生境异质性影响的结果。图1(b)中的 “均方—区组”曲线出现了2个峰值,表明木荷在200m2和400m2范围内有明显的斑块,而米槠、丝线吊芙蓉、虎皮楠最小的斑块出现在100m2左右。由于受样地大小的限制,种群的大规模斑块未能在图上表现出来。
4 结论与讨论
(1) 九连山保护区米槠群落中,竞争力较强的种群主要表现为聚集分布,竞争力较弱的种群主要表现为随机分布。这与柴勇﹑李玉媛等研究的菜阳河自然保护区山地雨林主要乔木种群分布格局大体一致[5]。该群落中,种群的不同径级分布格局是随着径级的增加,先表现为聚集程度的增加,当种群的径级增长到一定程度时又表现为聚集程度的减小;群落优势灌木的分布格局是除狗骨柴与山血丹外,树参、草珊瑚、黄楠、鼠刺等4种优势灌木均表现为聚集分布;群落中各优势种群的格局规模都表现为多种格局规模分布。
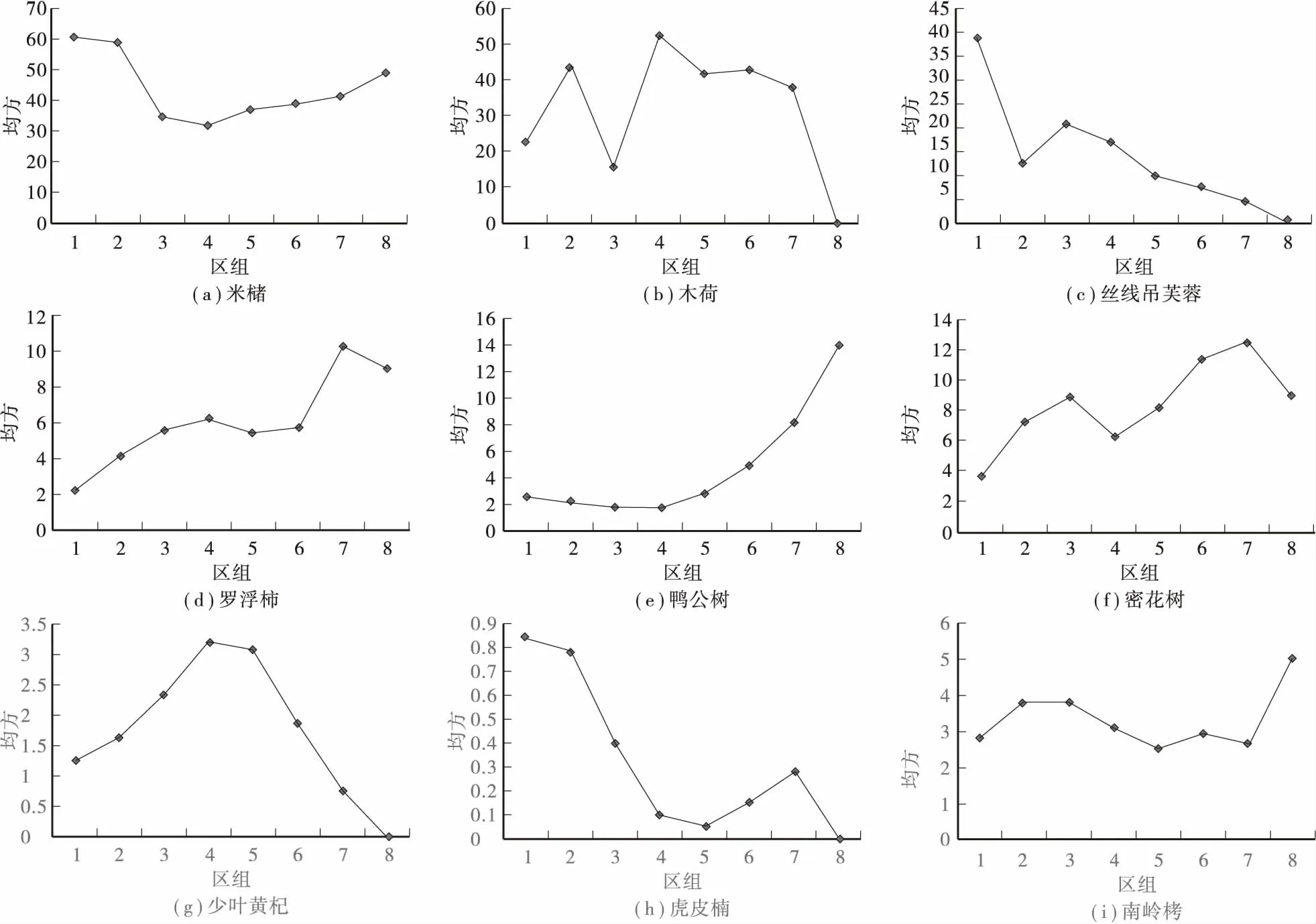
图1 优势乔木种群的“均方—区组”曲线Fig.1 The meansquare-block curve of dominant arbor populations
(2) 九连山保护区米槠群落正朝着正向顶级群落的方向发展。这是建立自然保护区后保护力度加强的结果。密花树、丝线吊芙蓉在本群落中能成为优势种,说明本群落曾受到过严重的干扰。
(3) 丛生指数、聚块性指数、负二项指数是常用的3种衡量聚集程度的指标。以该3种指标分析九连山保护区米槠群落的结果大致相同,但又略有差异。聚块性指数与负二项指数的计算公式分别为IP=1+I/m和K=m/I,式中m为某个种在样地中的株数的平均值,I为丛生指数;从其函数关系中知道聚块性指数IP与负二项指数K更加注重m的影响,也就是说2个种在I值相等时,如果某个种的m值相对较小,根据聚块性指数与负二项指数就会得出该种聚集程度相对较高的结论。
(4) 从格局规模分析得知:九连山保护区米槠群落中米槠、丝线吊芙蓉、虎皮楠等3个种群的均方峰值都出现在区组1。为了更清楚的显示这3个种群的斑块,有缩小样方大小的必要;鸭公树﹑南岭栲的第1个均方峰值都出现在区组8以后,因此,为了更清楚的显示这2个种群的斑块,有增大样地面积的必要。
[1] 宋晓英,牛树奎, 彭彪,等.福建三明不同更新方式的米槠林物种多样性研究[J].亚热带植物科学,2006,35(2):15-20.
[2] 肖忠优,刘信中.江西九连山自然保护区科学考察与森林生态系统研究[M].北京:中国林业出版社,2002:35-40.
[3] 陆阳.南亚热带森林种群分布格局取样技术研究[J].植物生态学与地植物学学报,1986,32(10):272-281.
[4] 朱祥福,林宝珠,周华.九连山碟斗青冈群落结构及小气候特征分析[J].江西林业科技,2006,34(5):4-9.
[5] 柴勇,李玉媛,方波,等.莱阳河自然保护区山地雨林主要乔木种群分布格局研究[J].西部林业科学,2005,34(9)40-44.
[6] Greig-Smith,P.Quantitative plant ecology.3rd ed[M].London,England Butterworths,1983.54-104.
[7] Sinclair DF.Ontests of spatial random ness using meall nearest neighbourdistan ce[J].Ecology,1985(66)84-85.
[8] 侯宽昭.中国种子植物科属词典(修订版) [M] .北京:科学出版社,1982:1-527.
[9] 孙儒泳,李庆芬,牛翠娟.基础生态学[M].北京:高等教育出版社,2004:152.
[10] 中国科学院中国植物志编辑委员会.中国植物志[M].北京: 科学出版社,1979:72.
[11] 张金屯.数量生态学[M].北京:科学出版社,2003:264.
[12] 张金屯,孟东平.芦芽山油松辽东栎优势树种空间分布格局研究[J].西北植物学报,2006,27(8):83-85.
(文字编校:唐效蓉)
Themainpopulations’spatialdistributionpatternofCastanopsiscarlesiicommunityinJiulianshanNatureReserve
XU Guoliang1, TANG Zhenghua2, LIAO Juan3
(1.Jiulianshan Nature Reserve Administrative Bureau, Longnan 341700, China; 2.Jiulianshan Forestry Station in Longnan County, Longnan 341700, China;3.Longnan Second Elementary School in Longnan County, Longnan 341700, China)

Jiulianshan Nature Reserve;Castanopsiscarlesiicommunity; main populations; distribution pattern
2012-12-10
2013-02-21
国家环保总局项目(20061A0013)。
徐国良(1982-),男,江西省龙南县人,本科,助理工程师,主要从事森林保护与森林生态研究工作。
Q 948.155
A
1003-5710(2013)02-0015-05
10. 3969/j. issn. 1003-5710. 2013. 02. 004
