和田河流域胡杨、灰叶胡杨种群结构特征的比较研究
2013-11-12周正立梁继业
周正立 梁继业 于 军 李 利
(1 塔里木大学植物科学学院,新疆 阿拉尔843300)(2 新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室,新疆 阿拉尔843300)(3 中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011)
种群是群落结构和功能的最基本单位,也是联系群落与个体之间的纽带,在生态学各层次研究中始终处于核心地位。在森林种群研究中,年龄结构与空间分布格局作为其重要特征和属性,不仅可反映种群数量动态、发展趋势的变化[1],而且能反映出种内种间关系、环境适应机制、空间异质性等[2]。因此,研究森林优势种群的年龄结构和空间分布格局,对阐明森林生态系统的形成与维持、群落的稳定性与演替规律、种群的生态特征和更新具有极为重要意义[3-6]。
胡杨(Populus euphratica oliv)、灰叶胡杨(Populus pruinosa schrenk)隶属于杨柳科胡杨亚属,主要生长在中纬度的干旱荒漠地区,并以新疆南部塔里木盆地沿河流两岸分布最多,形成了干旱荒漠区独特的河岸林生态景观,在稳定区域生态环境方面发挥着重要作用。然而,由于人类不合理的开发利用活动,以胡杨、灰叶胡杨为建群种和优势种的荒漠河岸林大面积减少,林地质量急速退化,在区域生态环境中产生了许多连锁反应。1993年FAO 林木基因资源专家组例会上,胡杨被确认为世界急需优先保护的林木基因资源[7]。
目前,众多学者对胡杨、灰叶胡杨开展了深入研究,主要涉及解剖学特征[8,9]、生理生态学特性[10,11]、种群结构[12,13]、生殖生态[14,15]与苗木繁育[16,17]等方面。关于种群结构的研究虽然较多,亦主要集中于塔里木河[18]、黑河[19]、疏勒河[20,21]流域,而关于和田河流域的研究缺乏,尤其是对胡杨、灰叶胡杨种群结构特征的比较研究较少。鉴于此,本研究以塔里木河源头之一——和田河流域的荒漠河岸林为研究对象,比较其优势种群的结构特征差异,以为和田河流域荒漠河岸林培育及退化生态系统恢复提供理论依据与技术支撑。
1 研究区自然概况
研究区位于和田河流域的和田县、洛甫县、墨玉县与策勒县境内。该区位于塔里木盆地西南缘,南接昆仑山,北连塔克拉玛干大沙漠,属暖温带极端干旱荒漠气候,具有光热资源丰富、昼夜温差大、降水稀少、蒸发强烈、无霜期长等特点。年总辐射量为138.1~151.5 千卡/平方厘米,仅次于青藏高原;全年日照时数2 470~3 000 小时,年平均日照百分率在58%~60%之间,最高可达84%;年平均温度11.6℃,≥10℃积温4 200℃,无霜期170~201天;年降水量13.1~48.2毫米,年蒸发量2 450~3 137毫米,干燥度大于20;四季多风沙,每年浮尘天气220 天以上,其中浓浮尘(沙尘暴)天气在60 天左右。
该区荒漠河岸植被物种组成单一,乔木层以胡杨、灰叶胡杨为主,灌木层主要有柽柳、骆驼刺、铃铛刺、黑果枸杞,草本层主要有芦苇、胀果甘草、苦豆子、叉枝丫葱、花花柴、牛皮消、沙旋覆花、天门冬、河西苣、盐生草、羊角草、罗布麻等,物种数目变化在2~9 种之间。乔木层郁闭度在0.16~0.58 之间,灌木层盖度0.29~6.17% 之间、草本层盖度在7.39~49.76%之间,群落结构相对简单。
2 研究方法
2.1 样地设置及外业调查
对和田河流域荒漠河岸林进行踏查的基础上,在和田县、洛甫县、墨玉县与策勒县境内分别选择有代表性的地段,根据具体情况设立大小50 m ×50 m、100 m×50 m、150 m×50 m 或20 m ×30 m的胡杨、灰叶胡杨样地共14 块。其中,胡杨5 块、灰叶胡杨9 块,各样地林分状况见表1。在各样地内以5 m×5 m 为基本单元,用相邻格子法进行每木调查,记录H≥2 m的全部乔木树种的胸径、树高、冠幅等,并对其幼苗幼树逐一记录其种名、地径与高度。灌木层、草本层也同时进行高度、盖度、多度等详细调查。
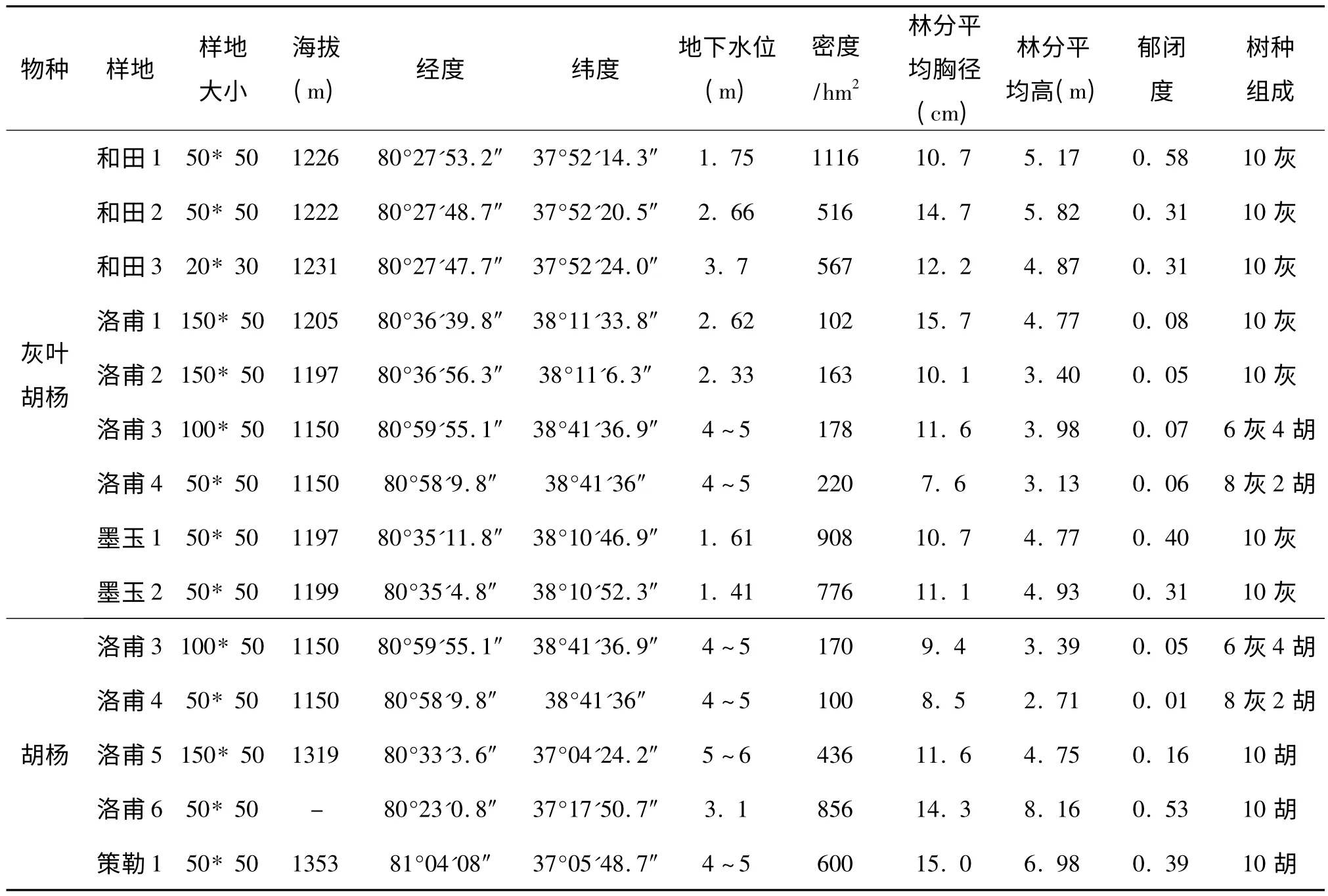
表1 胡杨、灰叶胡杨种群各标准地林分状况
2.2 种群大小级划分及静态生命表编制
在森林种群年龄结构研究中,确定所有个体的年龄往往较困难,使得以立木径级结构代替其年龄结构的大小级结构分析法得到广泛应用[4]。本文结合所调查林分的平均胸径,按4 cm 整化径阶划分立木大小级。其中,胸径小于2 cm的植株作为Ⅰ级,胸径≥2 cm的植株每隔4 cm 划为1 级。即胸径为2 cm≤D<6 cm的植株为Ⅱ级,6 cm≤D<10 cm的为Ⅲ级,依此类推。按此标准统计各径级的植株个体数,然后以大小级为横坐标,植株个体数为纵坐标,绘制胡杨、灰叶胡杨种群大小级结构图。
同时,把径级从小到大的顺序看作龄级顺序,Ⅰ径级对应第Ⅰ龄级,Ⅱ径级对应第Ⅱ龄级,依此类推。然后,以各径级的植株个体数作为各龄级的植株个体数,按江洪的方法对数据进行匀滑处理[22]并标准化,分别编制胡杨、灰叶胡杨种群静态生命表,并绘制存活曲线、死亡率和消失率曲线。
2.3 种群空间分布格局分析
2.3.1 不同发育阶段胡杨、灰叶胡杨种群分布格局分析
根据各胡杨、灰叶胡杨样地内的数据资料,以5 m×5 m 为标准对各径级个体在空间上的分布格局类型及聚集程度进行判定与分析。离散分布理论拟合采用方差/均值(扩散系数)的t 检验方法;聚集强度指标采用聚集度指数I、平均拥挤度指数M*、聚块性指数PAI、聚集指数Ca 和负二项式分布指数K[23]。
2.3.2 不同尺度胡杨、灰叶胡杨种群分布格局分析
对各胡杨、灰叶胡杨样地的调查数据,分别按5 m×5 m(1S)、5 m×10 m(2S)、10 m×10 m(4S)、10 m × 15 m(6S)、15 m × 15 m(9S)、15 m × 20 m(12S)、20 m×20 m(16S)、20 m ×25 m(20S)和25 m×25 m(25S)等尺度统计处理,并按2.3.1的方法对胡杨、灰叶胡杨种群的格局规模进行判定分析。
3 结果与分析
3.1 胡杨、灰叶胡杨种群大小级结构比较
由图1、2 可以看出,和田河流域胡杨、灰叶胡杨种群的年龄结构均呈纺锤形,并以Ⅳ径级的植株数量最多,分别占总株数的33.31%和39.56%。按刘普幸[20]的划分方法,可以看出无论是胡杨还是灰叶胡杨种群,均以中树(Ⅲ~Ⅵ)的植株数量最多,分别占总数的76.92%和81.16%,平均为373 株/hm2和410 株/hm2;而大树(Ⅶ~Ⅷ)稀少,占总数的1.98%和0.99%,平均为10 株/ hm2和5 株/hm2。此外,胡杨、灰叶胡杨种群的幼苗(Ⅰ级)、幼树(Ⅱ级)数量表现有所不同,在灰叶胡杨种群中幼树多于幼苗,幼苗仅占4.55%;而胡杨种群的林下更新好于灰叶胡杨,种群中幼苗数量多于幼树,占15.41%。但是,胡杨、灰叶胡杨种群总体上都表现为成年树多于幼树,表明种群有衰退可能。
3.2 胡杨、灰叶胡杨种群静态生命表


静态生命表是用同一时期收集到的植物样地内一个种群所有个体的年龄数据编制而成的生命表,可用于长生命期多年生植物(主要是乔木和灌木)的种群动态分析[24]。

由表2、图3 可以看出,在幼苗→幼树→中树→大树的整个生长阶段,灰叶胡杨种群的标准化存活数在Ⅰ~Ⅶ径级小于胡杨(意味着死亡率高于胡杨),在Ⅷ径级以后高于胡杨,这表明在生长发育阶段早期,灰叶胡杨种群较胡杨种群更易受到环境条件的影响。分别对胡杨、灰叶胡杨的种群存活曲线采用指数方程、直线方程和幂函数进行拟合(见表3),结果表明线性方程的拟合效果最好,相关系数分别为0.934 和0.982;其次以指数方程的拟合效果较好,相关系数分别为0.888 和0.952。根据Hett 和Loucks 估算存活状况的检验方法,指数方程式Nx=N0e-bx可描述I 型存活曲线,直线方程式Nx=N0+bx 可描述Ⅱ型,幂函数式Nx=N0x-b可描述Ⅲ型[24]。因此,胡杨、灰叶胡杨种群的存活曲线介于DeeveyⅠ和Ⅱ型之间,更近似于Deevey Ⅱ型。
从死亡率和消失率曲线来看(图4、5),胡杨、灰叶胡杨种群的死亡率和消失率曲线变化趋势相同,但两个种群之间表现有所不同。在灰叶胡杨种群中,其死亡率曲线呈双峰型,高峰值分别出现在第Ⅴ和第Ⅸ径级,死亡率最高值可能就是种群生活史中的脆弱阶段;而胡杨种群的死亡率随径级增加呈递增趋势。

表2 胡杨、灰叶胡杨种群静态生命表

表3 胡杨、灰叶胡杨种群存活曲线拟合方程
从期望寿命来看,灰叶胡杨种群的期望寿命呈“降-升-降”的趋势,峰值分别出现在第Ⅰ和第Ⅵ龄级;而胡杨种群的期望寿命呈单调递减趋势。Ⅰ龄级期望寿命较大可能由于幼苗摆脱草本和灌木的束缚后,个体所受到的竞争较小所致;Ⅵ龄级期望寿命较大表明这是种群生存质量最高的时期。另外,从同一龄级的期望寿命来看,胡杨种群在Ⅰ~Ⅴ龄级高于灰叶胡杨种群,而在Ⅵ龄级以后低于灰叶胡杨种群。期望寿命越高,说明种群的生存能力越强。

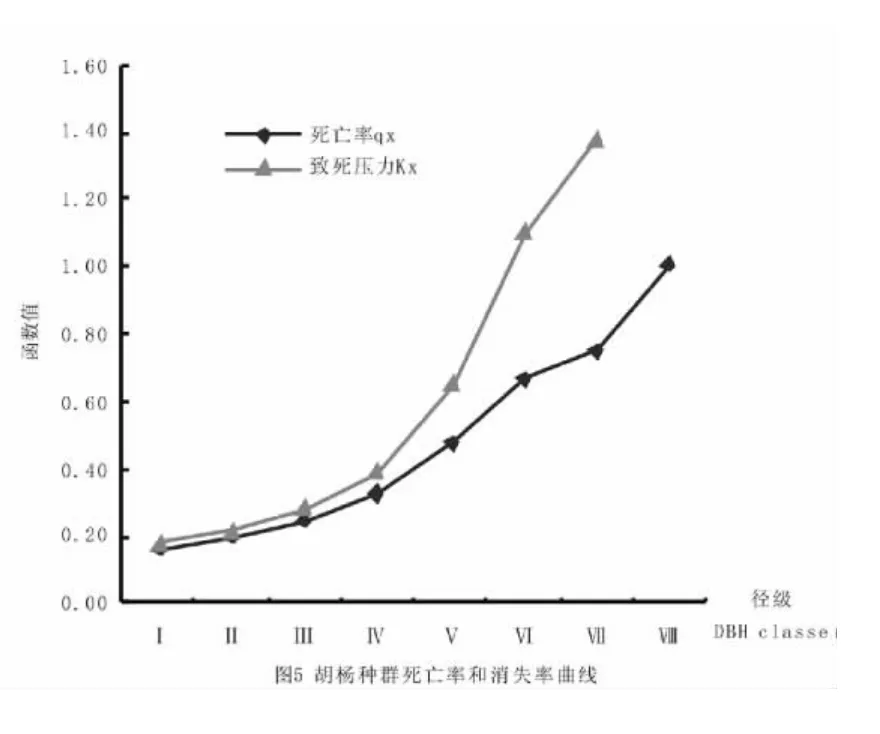

表4 不同发育阶段胡杨、灰叶胡杨种群空间分布格局结果
3.3 胡杨、灰叶胡杨种群空间分布格局的比较
3.3.1 不同发育阶段胡杨、灰叶胡杨种群空间分布格局
由表4 可以看出,在从幼苗(Ⅰ)→幼树(Ⅱ)→中树(Ⅲ~Ⅵ)→大树(Ⅶ以上)的发育过程中,胡杨、灰叶胡杨种群的空间分布格局在Ⅰ~Ⅵ龄级呈聚集分布,Ⅶ~Ⅷ龄级呈均匀分布。在Ⅰ~Ⅵ龄级从扩散系数(方差/均值)、聚集度指数、平均拥挤度来看,胡杨、灰叶胡杨种群的聚集强度总体表现为随年龄增加呈“先增后降”(单峰型)趋势,并分别在Ⅱ、Ⅲ龄级达到最大;其后,随树木生长发育对营养空间的需求增加,林木之间产生自疏现象,使得种群聚集强度逐渐下降,由聚集分布→均匀分布扩散。但从聚块性指数、聚集指数和负二项分布指数来看,胡杨、灰叶胡杨种群的聚集强度随年龄增加呈“双峰型”,两个峰值分别出现在Ⅱ和Ⅵ龄级。这说明六种指标的分析结果并不一定完全一致,这可能是由于扩散系数、聚集度指数、平均拥挤度指数的结果是以为计算依据,而聚块性指数、聚集指数、负二项分布指数是以为计算依据。
3.3.2 不同尺度胡杨、灰叶胡杨种群空间分布格局
由表5 可以看出,胡杨、灰叶胡杨种群在不同尺度上的分布型均属于聚集分布,并随尺度增加,由扩散系数、聚集度指数、平均拥挤度指数反映出的胡杨、灰叶胡杨种群聚集强度呈递增趋势,而负二项分布指数、聚块性指数、聚集指数反映出的聚集强度总体上呈递减趋势。从不同尺度上两个种群的聚集强度大小来看,在1S~6S 尺度上胡杨种群的聚集强度总体上大于灰叶胡杨种群,而在9S~25S 尺度上,胡杨种群的聚集强度小于灰叶胡杨种群。

表5 不同尺度胡杨、灰叶胡杨种群空间分布格局结果
4 结论与讨论
4.1 胡杨、灰叶胡杨种群年龄结构
种群年龄结构是种群的重要特征,反映了过去的更新机会和后来面临的死亡危险[25]。由于种群所处的生态环境不同在其年龄和个体大小上表现出某些差异,因而通过大小级结构探索种群年龄结构已成为研究种群动态的有效方法。和田河流域胡杨、灰叶胡杨种群的大小级结构呈纺锤形,以Ⅲ~Ⅴ径级的中树最多,这与瓜州绿洲[26]胡杨种群大小级的正态分布相似,与额济纳[19]、敦煌[21]绿洲的倒金字塔形不同。从林下更新苗(幼苗、幼树)数量来看,胡杨种群的更新好于灰叶胡杨。这可能与胡杨主要分布于远离河道的地方,其为了保持种族延续将较多的能量供应于生殖生长;而灰叶胡杨主要分布于靠近河道或地下水位较好的地方,其将更多的能量用于营养生长有关。
4.2 胡杨、灰叶胡杨种群存活曲线
存活曲线能够反映生物种群存活率或死亡率随年龄变化的情况,可根据其类型来判断森林树木种群是增长、下降还是稳定的动态特征[27]。和田河流域胡杨、灰叶胡杨种群的存活曲线介于DeeveyⅠ和Ⅱ型之间,更近似于Deevey Ⅱ型,按Leak[28]对存活曲线的划分属于稳定型种群,在整个生活史中种群有较为稳定的死亡率,与疏勒河[20]的DeeveyⅠ型不同。在生长发育过程中,灰叶胡杨种群死亡率曲线成双峰型,死亡高峰出现在中树和大树阶段的第Ⅴ和第Ⅸ径级,环境筛对种群有强烈的选择作用;而胡杨种群死亡率随径级增加呈递增趋势,生活史脆弱阶段主要出现在大树阶段。
4.3 胡杨、灰叶胡杨种群空间分布格局
树种在不同发育阶段会表现出不同的空间格局,这与森林群落的自然稀疏过程、干扰格局及环境变化有密切关系,同一年龄的个体在不同尺度下也有不同的分布格局,这主要与环境条件的空间变化相联系[29,30]。在由幼苗(Ⅰ)→幼树(Ⅱ)→中树(Ⅲ~Ⅵ)→大树(Ⅶ以上)的发育过程中,和田河流域胡杨、灰叶胡杨种群在Ⅰ~Ⅵ龄级呈聚集分布,Ⅶ~Ⅷ龄级呈均匀分布,并且聚集强度在Ⅱ、Ⅲ和Ⅵ龄级较大。Ⅱ、Ⅲ龄级聚集强度大,主要与胡杨、灰叶胡杨种群根蘖繁殖的幼苗和幼树聚集分布在母树周围有关;其后随着林木自疏现象的出现,导致种群聚集强度逐渐下降;而在Ⅵ龄级时聚集强度又有所增加,则可能与环境选择有关。此外,在不同尺度上胡杨、灰叶胡杨种群均表现为聚集分布,聚集强度在1S~6S 尺度上表现为胡杨种群总体上大于灰叶胡杨种群,而在9 S~25 S 尺度上表现为胡杨种群小于灰叶胡杨种群。这与塔里木河胡杨种群聚集强度与聚集规模均大于灰胡杨的结论有所不一致[18],其原因可能与水文水资源、气候、土壤和地形等异质性环境的差异有关。
[1]彭少麟.森林群落波动的探讨[J].应用生态学报,1993,4(2):120-125.
[2]张育新,马克明,祁建,等.北京东灵山海拔梯度上辽东栎种群结构和空间分布[J].生态学报,2009,29(6):2789-2796
[3]李博,杨持,林鹏.生态学[M].北京:高等教育出版社,2000.
[4]Harper J L.Population biology of plants[M].New York:Academic Press,1997.
[5]徐学红,于明坚,胡正华,等.浙江古田山自然保护区甜槠种群结构与动态[J].生态学报,2005,25(3):645- 653.
[6]杨永川,达良俊,陈波.天童米槠-木荷群落主要树种的结构及空间格局[J].生态学报,2006,26(9):2927- 2938.
[7]周正立,李志军,龚卫江,等.胡杨、灰叶胡杨开花生物学特性研究[J].武汉植物学研究,2005,23(2):163-168.
[8]李志军,焦培培,周正立,等.胡杨横走侧根及不定芽发生的形态解剖学研究[J].北京林业大学学报,2011,33(5):42-48
[9]杨赵平,刘琴,李志军.胡杨雌雄株叶片的比较解剖学研究[J].西北植物学报,2011,31(1):79-83.
[10]白雪,张淑静,郑彩霞,等.胡杨多态叶光合和水分生理的比较[J].北京林业大学学报,2011,33(6):47-52
[11]曹生奎,冯起,司建华,等.胡杨光合蒸腾与影响因子间关系的研究[J].干旱区资源与环境,2012,26(4):155-159
[12]张青青,徐海量,安红燕,等.塔里木河下游河水漫溢后胡杨幼苗分布格局初探[J].西北植物学报,2011,31(11):2298-2303
[13]崔德宝,于瑞德,于洋,等.新疆艾比湖自然保护区阿奇克苏河沿岸胡杨年龄结构和生长量状况研究[J].干旱区资源与环境,2012,26(1):194-200
[14]刘建平,周正立,李志军,等.胡杨、灰叶胡杨果实空间分布及其数量特性的研究[J].植物研究,2005,25(3):336-343
[15]曹德昌,李景文.胡杨种群生殖生态学研究进展[J].科学技术与工程,2009,9(5):1202-1209
[16]刘建平,周正立,李志军,等.胡杨、灰叶胡杨不同种源苗期生长动态研究[J].新疆环境保护,2004,26(zk):107-111
[17]安桂香,曾凡江,刘波,等.胡杨种子出苗对沙埋和供水条件的响应[J].中国沙漠,2011,31(2):436-431
[18]于军,王海珍,陈加利,等.塔里木河流域荒漠河岸林胡杨群落的空间格局研究[J].中国沙漠,2011,31(4):913-918
[19]郝兴明,赵万羽,陈忠升.额济纳胡杨种群年龄结构与树干空心分布特征[J].干旱区地理,2011,34(5):794-799
[20]刘普幸.疏勒河中下游绿洲胡杨种群结构与动态研究[J].自然资源学报,2011,26(3):429-439
[21]刘普幸,鹿晨昱,姚晓军,等.敦煌绿洲不同生境胡杨种群结构与空间格局研究[J].北京林业大学学报,2011,33(2):48-52
[22]江洪.云杉种群生态学研究[M].北京:中国林业出版社,1992:8-50.
[23]范繁荣,潘标志,马祥庆,等.白桂木的种群结构和空间分布格局研究[J].林业科学研究,2008,21(2):176-181
[24]Silvertown J W.Introduction to plant population ecology[M].London:Longman Press,1982.
[25]韩忠明,马书民,李晓璐,等.刺五加种群结构的研究[J].吉林农业大学学报,2009,31(2):170-173
[26]刘普幸,张杰.瓜州绿洲胡杨种群结构与动态研究[J].中国沙漠,2012,32(2):407-412
[27]于大炮,周莉,董百丽,等.长白山北坡岳桦种群结构及动态分析[J].生态学杂志,2004,23(5):30-34.
[28]Leak WB.An expression of diameter distribution for unbalanced uneven- aged stands and forests[J].For.Sci.,1964.10:19-50.
[29]宋于洋,李园园,张文辉.基于Ripley的K(r)函数和分形维数的梭梭种群空间格局[J].应用生态学报,2010,21(4):827-835.
[30]李立,陈建华,任海保,等.古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析[J].植物生态学报.2010.34(3):241-252
