野老鹳草在安徽夏熟作物田过去20年间的恶化规律及其影响因子
2013-10-24陈国奇何云核
陈国奇, 何云核,2, 强 胜
(1.南京农业大学杂草研究室,江苏南京210095; 2.浙江农林大学风景园林与建筑学院,浙江杭州 311300)
野老鹳草在安徽夏熟作物田过去20年间的恶化规律及其影响因子
陈国奇1, 何云核1,2, 强 胜1
(1.南京农业大学杂草研究室,江苏南京210095; 2.浙江农林大学风景园林与建筑学院,浙江杭州 311300)
调查了野老鹳草在安徽夏熟作物田间的发生情况,并与历史资料进行对比分析,目的是揭示其在田间发生、演替的规律及其环境影响因子。结果表明:在过去20年间,野老鹳草的发生频度由23.08%上升到91.84%,在样地中的平均优势度由0.024极显著上升到0.559(P<0.01),这种演替主要发生在年降水量增加的皖南地区和人口密度上升较快的淮北地区。水旱连作田的发生频度显著高于旱连作田,农田作物连作模式成为影响野老鹳草发生频度的关键因子。而影响野老鹳草发生优势度的主要环境因子则是最热月均温,低于28.7 ℃的水旱连作田中野老鹳草发生的优势度最高,而最热月均温高于28.7 ℃的旱连作田中该种发生的优势度最低。
入侵杂草;历史调查数据;近期调查数据;气候;耕作活动
外来杂草入侵严重危害了我国农作物生产和农田生物多样性,并且难以有效防除[1],了解影响农田外来杂草发生和扩散的关键环境因子是有效对其进行综合防控的前提和基础[2]。一年生杂草野老鹳草(GeraniumcarolinianumL.)原产北美,现已侵入我国华东、华中、华北和华南及西南10多个省(市),成为油菜和小麦田间的主要恶性杂草[3],并且该种的扩散潜力非常大[4]。
恶性入侵杂草往往具有快速适应能力[5],尤其是在人工干扰强烈、资源丰富的生境中[6]。因而,当前我国农业耕作模式的变革可能会促进恶性入侵杂草在农田中的扩散。例如,由于资源丰富的生境更容易遭受外来植物入侵,因此长期大量施用化肥很可能会促进外来杂草的侵入和蔓延[7];大量喷施除草剂会导致抗药性杂草种群的出现和快速蔓延,而许多恶性入侵杂草都具有较高的可塑性和快速进化的能力,因而更容易产生除草剂耐性或抗性种群[8-9];农业机械的应用为外来杂草种子传播提供了新的高效途径[10];此外,气候变化可能会进一步促进外来入侵杂草在农田中的扩散[11]。
通过对比历史数据和近期数据,可以很好地揭示外来杂草发生和扩散的规律。本研究在2005—2010年调查数据的基础上,与1987—1990年调查数据进行对比,研究了野老鹳草的分布和发生格局以及影响该种扩散的关键环境因子,以期为该种的防控提供有效的理论参考,同时以此作为一个典型例子为我国农田入侵杂草发生规律的研究积累数据资料。
1 材料与方法
1.1 数据收集
本研究数据包含2个部分——1987—1990年和2005—2010年的调查数据分别作为安徽地区夏熟旱作物田入侵杂草的历史数据和近期数据。历次调查均采用7级目测法[12],记录每块样田中野老鹳草发生的优势度等级,即根据杂草在田间的高度、盖度和多度确定其在田间的优势度(分为7级)。历史数据共调查了130个样地,包括83块油菜田和47块小麦田;近期数据共调查了147个样地,包括78块油菜田和69块小麦田(图1);每个样地设置10块样田,每块样田均选择667 m2左右的田块进行调查[12]。
本研究收集了所调查样地的11个环境因子信息(表1),包括调查样地田块中的作物、前茬作物、以及调查地所处区域内当年的最冷月均温、最热月均温、年均温、年降水量、人口密度、交通频度(单位土地面积交通运输周转量)、单位耕地面积农业机械总动力、单位耕作面积化肥和除草剂的施用量。除作物和前茬作物外的其他9个样地环境因子采用样地所在地级市当年的官方统计数据[13]。除草剂施用量参考了统计年鉴和调查期间对各地植保站的咨询数据。
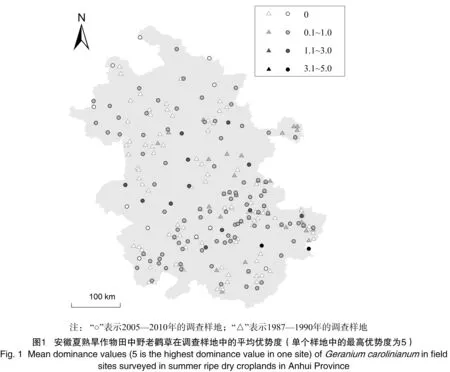
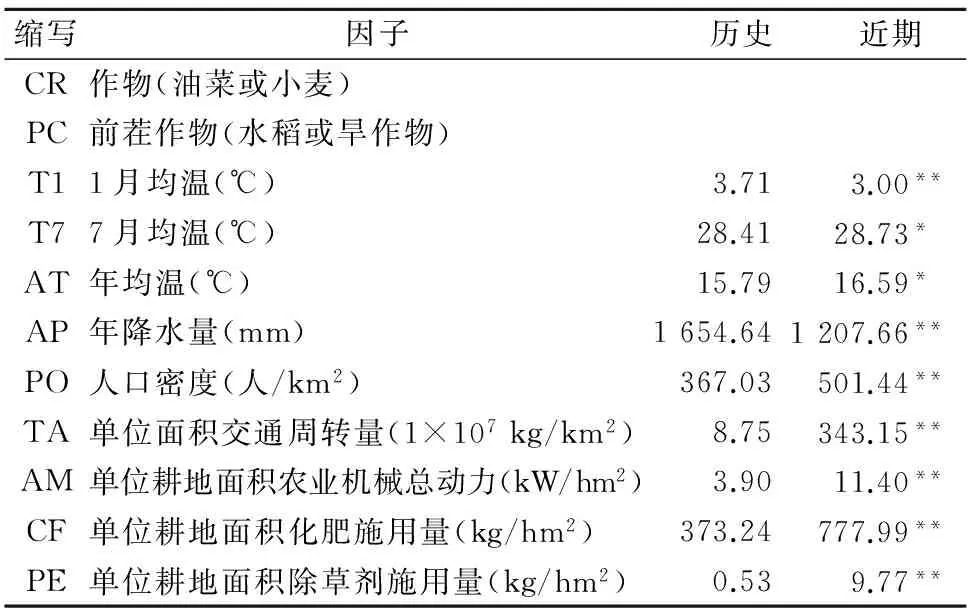
表1 调查样地11个环境因子及其历史和近期平均值比较Table 1 Mean values of 11 environmental variables analyzedin this study
注:历史调查样地环境因子和近期调查样地环境因子间的比较采用独立样本T检验进行。
1.2 数据统计分析
对野老鹳草的发生情况分别采用2种数据类型进行分析,即多度数据和0/1数据(有或无)分别形成样地-发生情况数据矩阵。本研究采用决策树模型 (decision tree model)[2,14]分别分析了影响当前外来入侵杂草发生和分布的环境因子。决策树模型又称为分类回归树模型(classification and regression tree model),它作为一种非参数的方法可以有效处理非线性关系[2,15],它将环境因子梯度作为分类节点,利用递归划分法将样方划分为尽可能同质的类别,实现各个类群内的总方差最小[14]。由于决策树模型是以环境梯度作为分类节点的,所以无需人为确定分类结果。决策树模型可以处理各种不同类型的环境因子数据(即包括数值型和分类型的数据),并将结果展示为树状结构图。由于该模型在运算过程中对环境因子进行逐一处理,因而可以有效避免环境因子间的多重共线性(multicollinearity)所导致的结果偏差[2]。因此决策树模型在生态学研究中被广泛应用,尤其是关于入侵种的分布,并且近年来逐渐被引入农田杂草的研究中。本研究的决策树模型采用R软件中“party”程序包的“ctree”函数进行运算[16]。“ctree”函数通过递归划分法将运算结果生成二岐决策树,并且在递归划分的每一步分别进行基于条件推理(conditional inference framework)的模拟检验(permutation test)。因而“ctree”函数能够避免环境因子选择上的偏差,进而得到最优的回归分类结果。此外,该函数通过自带的检验步骤自动选择统计上的最优分类结果,避免了人为选择。
为了定量地比较调查区域内野老鹳草发生情况的变化及其与环境因子间的关系,本研究采用地级市作为分析的基本单位,即以一个地级市内所有样地各个统计指标的平均值作为该市的各个指标,从而建立地级市-发生量和环境因子数据矩阵,共得到14个地级市的兼具历史调查和近期调查数据,这14个地级市分别是:安庆、蚌埠、亳州、巢湖、池州、滁州、阜阳、合肥、黄山、六安、马鞍山、芜湖、宿州和宣城。然后对这14个地级市的各个指标数据上的变化(温度和降水指标采用近期数据减去历史数据,其他指标采用近期数据除以历史数据),采用逐步回归模型(stepwise regression model)[17-18]来分析导致安徽夏熟旱作物田野老鹳草发生量加剧的关键环境因子。在最优回归模型的选择上,采用AIC检测标准(Akaike’s information criterion),即AIC值最小的为最优模型[19-20]。
2 结果与分析
在过去20年间,野老鹳草在调查样地中的频度由23.08%上升到91.84%(图1),样地平均优势度由0.024极显著上升到0.559(P<0.01),而在野老鹳草出现的样地中,其平均优势度由0.102极显著上升到0.608(P<0.01)。
2.1 回归决策树
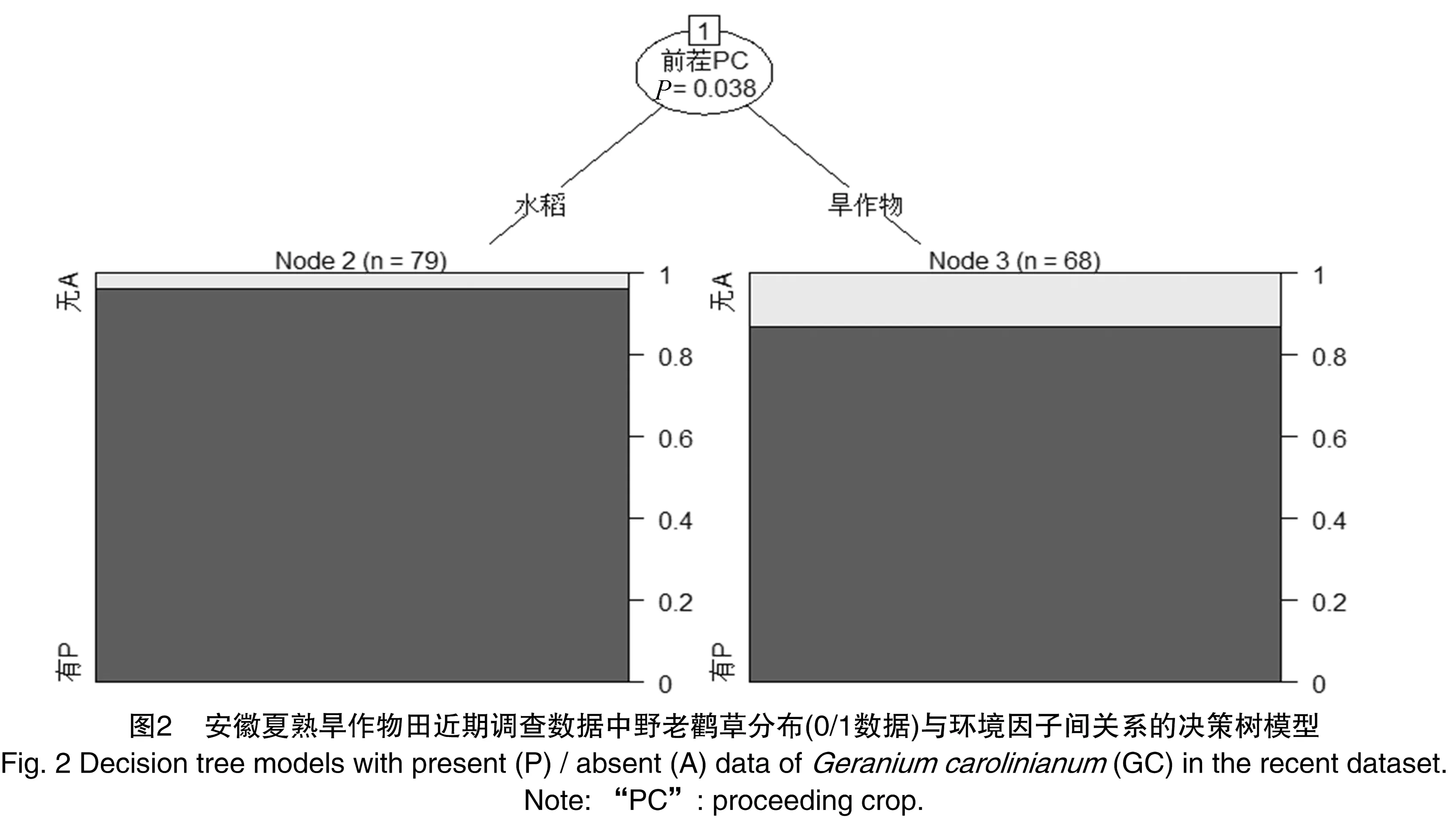
决定安徽夏熟旱作物田中野老鹳草发生频度回归决策树的因素是前茬作物(图2)。可见墅老鹳草在水旱连作田(前茬为水稻)中的发生频度显著高于旱连作田(前茬为秋熟旱作物)。
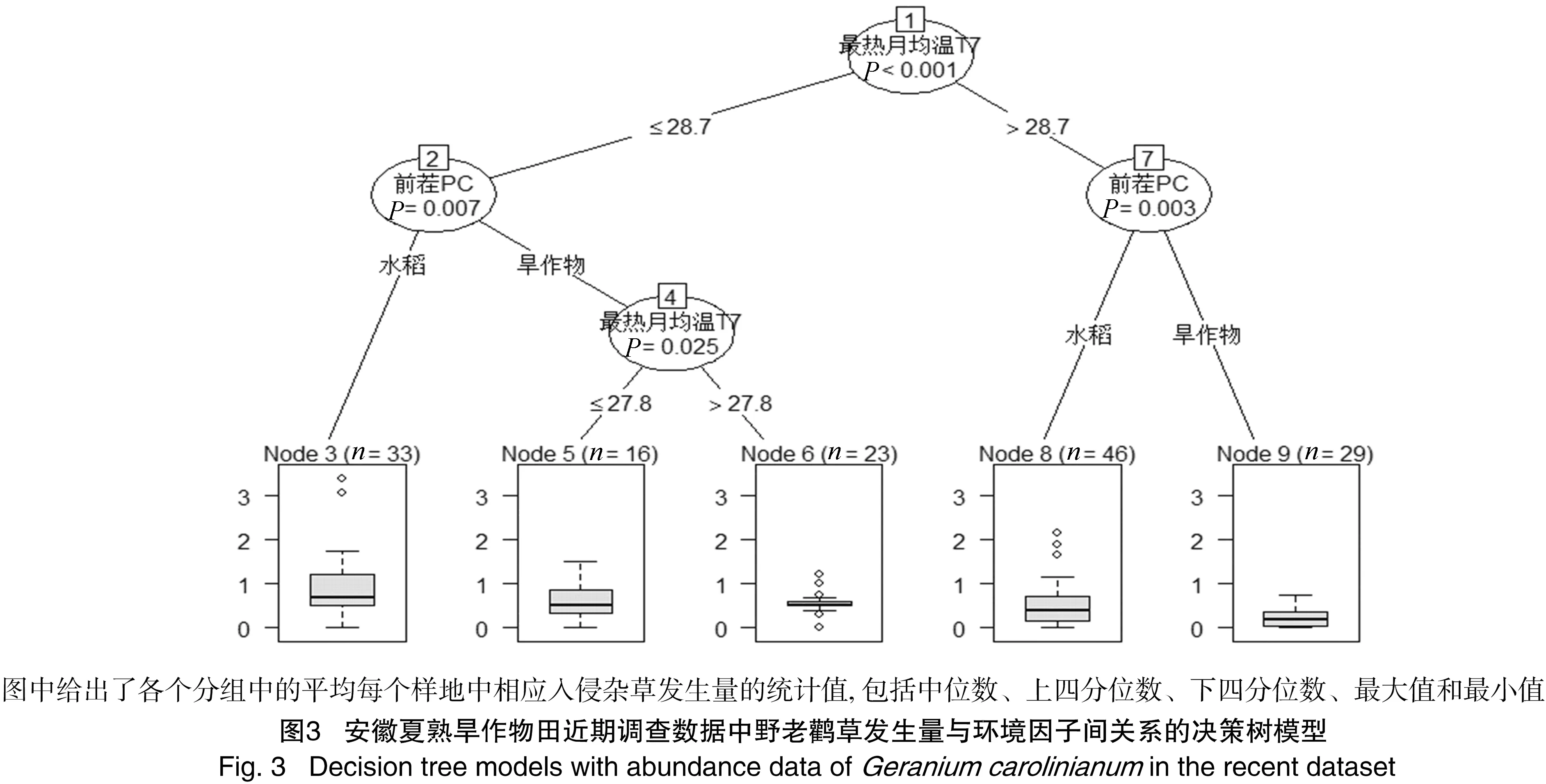
决定安徽夏熟旱作物田中野老鹳草发生的优势度分布的回归决策树第一分枝的因素是最热月(7月)均温(图3)。按最热月均温和前茬作物类型将调查样地分为5个类群,其中最热月均温低于 28.7 ℃ 的水旱连作田中野老鹳草发生的优势度最高,而最热月均温高于28.7 ℃的旱连作田中野老鹳草发生的优势度最低。
2.2 逐步回归
对野老鹳草发生量变化进行逐步回归分析得到的线性模型中包括6个环境因子(表2),即农业机械应用、化肥施用量、除草剂施用量、交通频度、年降水量和最热月均温的变化,年降水量和人口密度的变化与野老鹳草发生量的变化显著正相关 (P<0.05)。
3 讨论
3.1 温度和降水量的影响
本研究结果表明,最热月均温是影响野老鹳草田间优势度的最重要环境因子。在最热月均温较高地区,野老鹳草发生的优势度较低,然而最冷月温度和年均温对该入侵杂草的分布和扩散均无显著影响。降水量与野老鹳草在安徽夏熟旱作物田中的发生和分布现状无显著相关性,但是在过去20年间降水量增加较多的地方,野老鹳草的扩散也较快。气候条件是影响入侵植物分布的基本因素,因而原产地的气候条件是预测和评估入侵植物在入侵地潜在分布区和危害性的最重要因素[11,15,21]。野老鹳草是原产于北美洲的一年生入侵杂草,其在田间以种子进行繁衍和传播,花果期5月至6月[4]。有研究表明,野老鹳草种子中具有的同一层栅栏细胞构成的隔水层能有效促使种子物理休眠[22],因而野老鹳草种子在夏天成熟,且成熟后对土壤温度和湿度条件的适应力较强[22],最冷月温度对该种分布的影响较为有限。然而最热月均温反映了野老鹳草种子形

表2 安徽夏熟旱作物田中野老鹳草y优势度的变化量(近期调查数据减去历史调查数据)与环境因子变化量的逐步回归分析结果,以及各线性模型的AIC检测结果Table 2 Results of the stepwise regression models used to test the relationship between the changes in environmental factors andchanges in occurrence of Geranium carolinianum in summer ripe cropland in Anhui Province. Changes in Akaike’s informationcriteria between final and null models are also shown
注:温度和降水量指标的变化采用近期调查数据减去同一地级市的历史调查数据,其他指标均采用近期调查数据与历史调查数据的比值进行逐步回归,模型中排除的环境因子未列入表中。
成和成熟期间的温度,温度过高显然会抑制其生长和种子产量。降水量对野老鹳草分布的影响也较为有限,然而在较长的时间尺度上(例如本研究的20年左右),降水充沛地区野老鹳草的扩散较快。值得注意的是,安徽地区的年降水量在年际间起伏较大,全省绝大部分地区的暴雨量呈现上升趋势[23]。野老鹳草种子传播较小,在田间主要借水传播,因而暴雨极有可能促进该种子的扩散。
3.2 农田连作模式和栽培作物的影响
农田连作模式对野老鹳草的入侵和扩散具有显著影响。水旱连作田中野老鹳草发生频度和优势度较高,尽管同时在旱连作田中该种的发生频度也很高。栽培作物(油菜或小麦)对其分布和扩散无显著影响。野老鹳草都具有极强的适应能力和入侵性[3-4],在世界许多地方都被列为恶性杂草[24],对不同夏熟作物类型的农田都具有较高的适应性。因而,作为典型的杂草,野老鹳草在资源丰富、土壤湿润的生境下其更易建立种群并大量爆发[6]。水旱连作田土壤通常较为湿润,且水稻土通常比旱地更肥沃,此外水田浸水时间较长有利于野老鹳草种子在田间散布开来,从而减少同种个体间的竞争,此外种子充分浸水可以利于其破除物理休眠而萌发出苗[22],因而野老鹳草在水旱连作田中的危害比旱连作田重。
3.3 除草剂、化肥施用和农业机械应用的影响
除草剂和化肥施用量及农业机械化水平与野老鹳草的发生情况均无显著相关性。油菜田主要仅使用防除禾本科杂草的芳氧苯氧丙酸类除草剂,对野老鹳草无效。而麦田使用的苯磺隆、异丙隆等除草剂对野老鹳草效果较差,甚至长期使用这些除草剂,野老鹳草会对化学除草剂演化出较高的抗性[25- 26],并且其种子产量较高,出苗量极大,因此依赖除草剂控制野老鹳草的效果并不理想,尤其是从较长的时间尺度上看,大量施用除草剂甚至可能导致抗性杂草种群的迅速扩散。
化肥施用量的增加会促进外来杂草在农田中的入侵和扩散[7],尤其利于多年生入侵杂草的定植和扩散。而野老鹳草作为典型的一年生杂草,其生活史较短,在资源并不匮乏的农田生境下,化肥施用量加大会促进多种杂草的发生,进而可能挤压野老鹳草的生态位而抵消资源丰富的红利。
农业机械与外来杂草在农田中的入侵和扩散关系复杂,而本研究结果没有发现农业机械总动力对野老鹳草的发生和分布有显著影响。一方面,农田杂草防控势必需要应用农业机械;另一方面,农业机械的应用也可能为外来杂草在田间的传播提供新的有效途径[10]。
3.4 人口密度和交通
在过去20年间,人口密度的增加与野老鹳草发生量的增加呈显著正相关,也即人口密度较高地区野老鹳草发生量增幅较大。大量研究结果表明,人口密度与外来杂草的扩散显著相关[27-28],尤其是人类活动极大促进了外来杂草种子的传播[29-30]。野老鹳草种子较小、产量较高且传播能力有限,因而此类入侵杂草的种子往往依赖人类活动的传播,如灌溉、收割机运输等能够进行远距离传播[10,31]。野老鹳草种子无法有效借助风力传播,交通频度对该种的分布和扩散无显著影响。
4 结论
通过对野外调查数据的比较发现,过去20年间,野老鹳草在所调查农田中扩散极为迅速,并成为期间最严重的恶性杂草之一。并且在调查区域内,野老鹳草表现出明显的喜湿性,具体表现为其在水旱连作田间发生危害较重,并且在降水量增加的地区,其发生量上升较快。随着气候变化和农业耕作模式的变革,野老鹳草在我国农田中的危害极有可能进一步加剧。
[1]强 胜,陈国奇,李保平,等. 中国农业生态系统外来种入侵及其管理现状[J]. 生物多样性,2010,18(6):647-659.
[2]Pinke G,Karácsony P,Czúcz B,et al. Environmental and land-use variables determining the abundance of Ambrosia artemisiifolia in arable fields in Hungary[J]. Preslia,2011,83:219-235.
[3]李扬汉. 中国杂草志[M]. 北京:中国农业出版社,1998.
[4]徐海根,强 胜. 中国外来入侵生物[M]. 北京:科学出版社,2011.
[5]Clements D R,Ditommaso A,Jordan N,et al. Adaptability of plants invading North American cropland[J]. Agriculture Ecosystems & Environment,2004,104(3):379-398.
[6]Seastedt T. Plant ecology:resourceful invaders[J]. Nature,2007,446(7139):985-986.
[7]Kovacs-Hostyanszki A,Batary P,Baldi A,et al. Interaction of local and landscape features in the conservation of Hungarian arable weed diversity[J]. Applied Vegetation Science,2011,14(1):40-48.
[8]Powles S B. Evolved glyphosate-resistant weeds around the world:lessons to be learnt[J]. Pest Management Science,2008,64(4):360-365.
[9]Gaines T A,Zhang W,Wang D,et al. Gene amplification confers glyphosate resistance in Amaranthus palmeri[J]. Proceedings of the National Academy of Sciences of the United States of America,2010,107(3):1029-1034.
[10]Shirtliffe S J,Entz M H. Chaff collection reduces seed dispersal of wild oat(Avena fatua)by a combine harvester[J]. Weed Science,2005,53(4):465-470.
[11]Clements D R,Ditommaso A. Climate change and weed adaptation:can evolution of invasive plants Lead to greater range expansion than forecasted?[J]. Weed Research,2011,51(3):227-240.
[12]Analysis Q M. Description,and ecological interpretation of weed vegetation in the summer crop fields of Anhui province,China[J]. Journal of Integrative Plant Biology,2005,47(10):1193-1210.
[13]安徽省统计局. 安徽统计年鉴[M]. 北京:中国统计出版社,出版年度缺失:1987-2011.
[14]Loh W Y. Classification and regression trees[J]. Wiley Interdisciplinary Reviews:Data Mining and Knowledge Discovery,2011,1(1):14-23.
[15]Freckleton R P,Stephens P A. Predictive models of weed population dynamics[J]. Weed Research,2009,49(3):225-232.
[16]Hothorn T,Hornik K,Zeileis A. Unbiased recursive partitioning:a conditional inference framework[J]. Journal of Computational and Graphical Statistics,2006,15(3):651-674.
[17]Whittingham M J,Stephens P A,Bradbury R B,et al. Why do we still use stepwise modelling in ecology and behaviour?[J]. Journal of Animal Ecology,2006,75(5):1182-1189.
[18]Mundry R,Nunn C L. Stepwise model fitting and statistical inference:turning noise into signal pollution[J]. The American Naturalist,2009,173(1):119-123.
[19]José-María L,Armengot L,Blanco-Moreno J M,et al. Effects of agricultural intensification on plant diversity in Mediterranean dryland cereal fields[J]. Journal of Applied Ecology,2010,47(4):832-840.
[20]Burnham K P,Anderson D R. Model selection and multimodel inference:a practical information-theoretic approach[M]. New York:Springer,2002.
[21]Graziani A,Steinmaus S J. Hydrothermal and thermal time models for the invasive grass,Arundo donax[J]. Aquatic Botany,2009,90(1):78-84.
[22]Gama-Arachchige NS,Baskin JM,Geneve RL,et al. Identification and characterization of the water gap in physically dormant seeds of Geraniaceae,with special reference to Geranium carolinianum[J]. Annals of Botany,2010,105(6):977-990.
[23]谢五三,田 红. 近50年安徽省暴雨气候特征[J]. 气象科技,2011,39(2):160-164.
[24]Holm L G,Pancho J V,Herberger J P. A geographical Atlas of world weeds[M]. New York:John Wiley and Sons,1979.
[25]娄群峰,黄建中,张敦阳,等. 南京地区油菜田杂草群东特点及分布规律的研究[J]. 江西农业大学学报,1999,21(3):370-375.
[26]胡友忠. 使阔得春季防除小麦田野老鹳等阔叶杂草试验[J]. 安徽农学通报,2010,16(9):143-143,179.
[27]Chown S L,Van Rensburg B J,Gaston K J,et al. Energy,species richness,and human population size:conservation implications at a National scale[J]. Ecological Applications,2003,13(5):1233-1241.
[28]Pysek P,Jarosík V,Hulme PE,et al. Disentangling the role of environmental and human pressures on biological invasions across Europe[J]. Proceedings of the National Academy of Sciences of the United States of America,2010,107(27):12157-12162.
[29]Strubbe D,Matthysen E. Establishment success of invasive ring-necked and monk parakeets in Europe[J]. Journal of Biogeography,2009,36(12):2264-2278.
[30]陈国奇,李明丽,郭水良,等. 植物性状与苏浙沪地区草本被子植物入侵性的关系[J]. 生态学报,2009,29(9):5145-5151.
[31]Blanco-moreno J,Chamorro L,Masalles R,et al. Spatial distribution of Lolium rigidum seedlings following seed dispersal by combine harvesters[J]. Weed Research,2004,44(5):375-387.
IncreasingSeriousnessPatternofGeraniumcarolinianuminSummerCropFieldsinAnhuiProvinceDuringthePast20YearsandItsAffectingFactors
CHEN Guo-qi1, HE Yun-he1,2, QIANG Sheng1
(1.Weed Research Laboratory,Nanjing Agricultural University,Nanjing,210095,China;2.School of Landscape Architecture,Zhejiang Agricultural and Forestry University,Hangzhou 311300,China)
In order to characterize the pattern of occurrence and succession and the environmental factors,the current occurrence ofGeraniumcarolinianumwas investigated in summer crop fields in Anhui Province and compared to that recorded in historical data. Over the past 20 years,the frequency ofG.carolinianumincreased from 23% to 92% and its mean dominance value increased significantly from 0.024 to 0.559 (P<0.01) among the surveyed sites. These changes mainly occurred in the southern Anhui Province where annual precipitation increased and in the Huaibei area where population density also increased rapidly. Frequency in the land-water continuous cropping fields was significantly higher than that in the land continuous cropping field,and crop cultivation became the key factor affecting the occurrence ofG.carolinianum. The mean temperature of the hottest month was the main environmental factor affecting its dominance with dominance increasingand decreasing below and above a discriminating temperature of 28.7 ℃,respectively.
invasive weed;historic dataset;recent dataset;climate;farming practice
S451;Q948
A
1003-935X(2013)02-0013-06-
陈国奇,何云核,强 胜. 野老鹳草在安徽夏熟作物田过去20年间的恶化规律及其影响因子[J]. 杂草科学,2013,31(2):13-18.
2013-04-27
国家“973”计划(编号:2009CB119200);国家科技支撑计划(编号:2012BAD19B06);江苏省普通高校研究生科研创新计划(编号:CXZZ11.0647)。
陈国奇(1982—),男,浙江龙游人,博士研究生,研究方向为杂草科学。Tel:(025)84395117;E-mail:chenguoqi_21@163.com。
强 胜。Tel:(025)84395117;E-mail:wrl@njau.edu.cn。
