温度、盐度和密度交互作用对波纹巴非蛤幼贝生长的影响
2013-10-13高如承赖素兰
陈 志, 高如承, 胡 青, 李 鹏, 赖素兰
(福建师范大学 生命科学学院, 福建 福州 350108)
波纹巴非蛤(Paphia undulata)是一种肉质细嫩、口感鲜美、营养丰富的海产双壳贝类。隶属于软体动物门(Mollusca), 瓣鳃纲(Lamellibranchia), 帘蛤目(Veneroida), 帘蛤科(Veneridae), 巴非蛤属(Paphia),俗称“花蚶”、“油蛤”。它是一种暖水性浅海底栖贝类, 大都生活于大潮时水深0.5~3.5 m的潮下带浅海底部软泥中, 最深可分布到10 m水深。它具有高蛋白质、低脂肪的特点[1], 除了富含有多种氨基酸以外,还含有氨基多糖和牛磺酸, 易被肠道消化吸收, 食疗药用价值较高, 具有抗肿瘤、降低血糖和改善糖尿病症状等作用[2]。目前对波纹巴非蛤的研究涉及众多领域, 包括形态学[3-4]、繁育生物学[5-6]、生理学[7]、分子免疫学[8-9]、生态学[10]、食品开发与应用[1]、养殖技术与应用[11-12]等。
响应面法(Response Surface Methodolody, 简称:RSM)是先进行合理的试验方案设计, 后通过实验论证得到相关的实验数据, 采用多元二次回归方程拟合因素与响应值之间的函数关系, 通过对回归方程各参数的分析来寻求最优参数并解决多变量问题的一种统计方法[13-14]。应用软件系统对试验得到的数据进行统计分析, 观察响应面[15], 利用软件提供的三维立体图形和二维等高线图, 最优化试验结果。响应面法具有次数少、周期短, 所得回归方程精度高等优点, 可以研究几种因素间的交互作用[16]。本文利用响应面法, 探讨温度、盐度和密度对波纹巴非蛤幼贝生长的联合作用, 构建波纹巴非蛤幼贝生长模型并优化其生长条件, 以期对波纹巴非蛤大规模人工养殖提供科学理论指导。
1 材料与方法
1.1 实验材料
实验用波纹巴非蛤幼贝均来自福建宏峰泰海洋生物开发有限公司育苗基地, 于2012年6月人工育苗成功, 选取壳长相近的幼贝用于实验, 随机抽取100粒幼贝测量壳长, 平均壳长为4.94 mm±0.07 mm。模型验证实验选取同批次幼贝, 幼贝规格与响应面实验开始时基本相同, 随机抽取30粒幼贝测量壳长,平均壳长为5.04 mm±0.06 mm。
1.2 实验设计
1.2.1 响应面法分析实验
在前期单因素实验基础上, 选取温度、盐度和密度3个影响因子的实验组, 根据Box-Benhnken的中心实验设计原理进行3因素3水平(3因素为温度(A),盐度(B), 密度(C); 3水平为-1, 0, 1)的二次多项回归组合实验, 探索波纹巴非蛤幼贝生长的最优条件。实验在500 mL烧杯中进行, 烧杯底部铺设5 mm厚的底泥。不同温度梯度由恒温水槽控制, 通过添加淡水至天然海水(盐度为32~33)中配置不同盐度梯度。实验进行 20 d, 每天早晚投喂塔胞藻(Pyramidomonassp.),各实验组投喂饵料相同, 并在投喂前将藻液盐度调至与实验组相同。响应面影响水平、因素编码及实验设计见表1和表2。
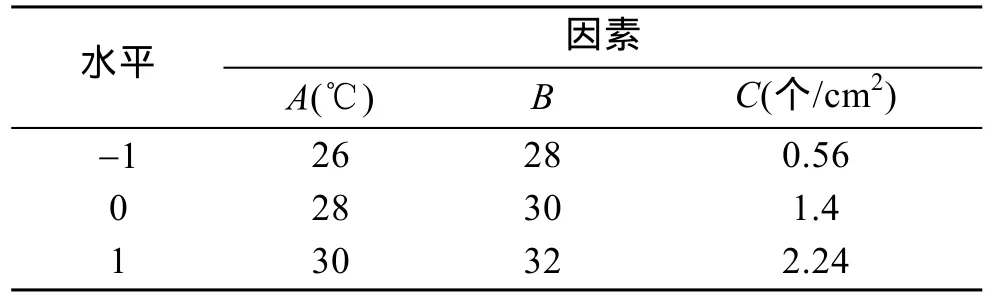
表1 实验水平和因素设计Tab. 1 Factors and levels of the experiment
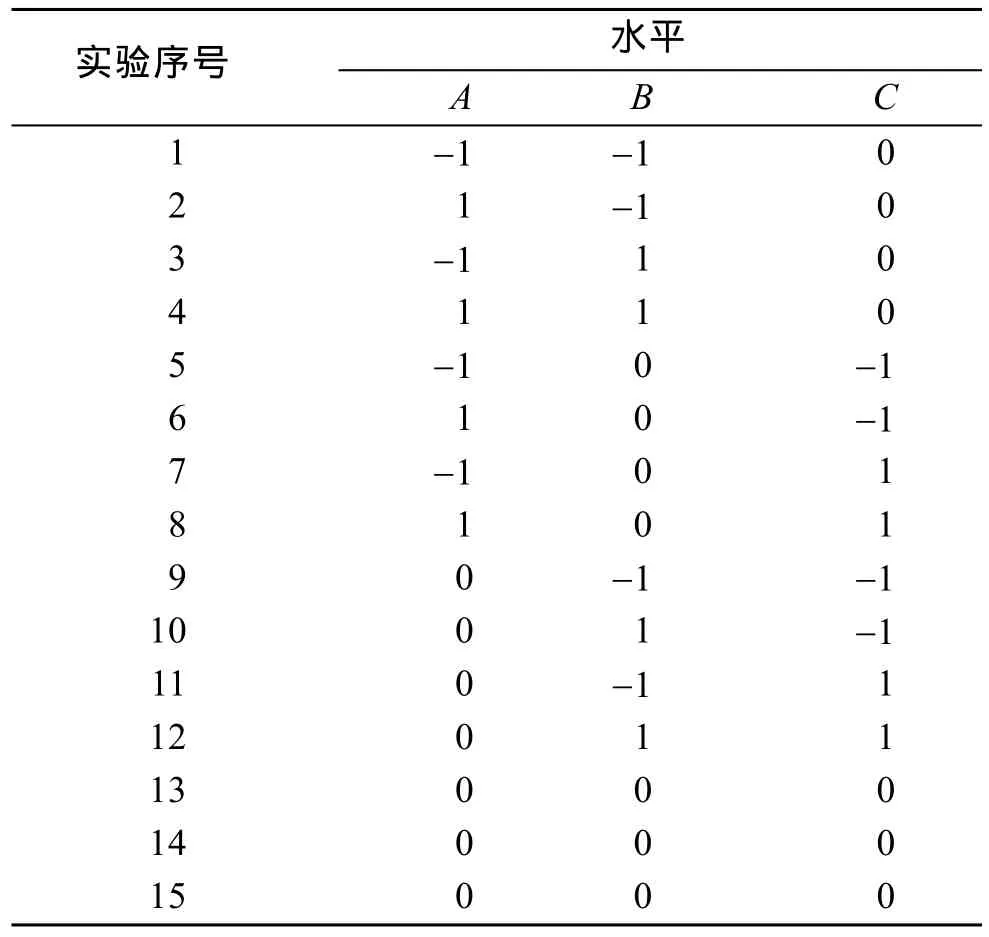
表2 响应面3因素3水平实验设计Tab. 2 The experiment design of 3 factors and 3 levels of RSM
1.2.2 模型验证实验
实验条件按照响应面优化得出的最适生态环境条件进行, 实验设置3个平行, 实验时间和响应面实验相同, 为期 20 d, 实验结束后统计各平行组的平均壳长和死亡个体数。
1.3 计算公式与数据分析
生长率:Rg=(L-L0)/L0
存活率:Rs=(D-D0)/D0
公式中L0,L分别表示实验开始和结束时幼贝的壳长(单位: mm),D0,D分别表示实验开始和结束时幼贝个数。
实验数据用软件SPSS17.0进行平均值、标准差及单因素方差ANOVA分析, 独立样本均数进行T检验, 其中P< 0.05认为有显著差异,P< 0.01认为差异极显著。采用SAS 9.2软件对响应面的实验数据进行二次多项回归拟合、方差分析、显著性检测和响应面法分析。
2 结果
2.1 回归模型和回归方程的建立
综合单因素影响的实验结果, 以幼贝生长率为响应值。采用SAS 9.2软件计算多元回归拟合, 建立二元回归方程。实验结果如表3所示。
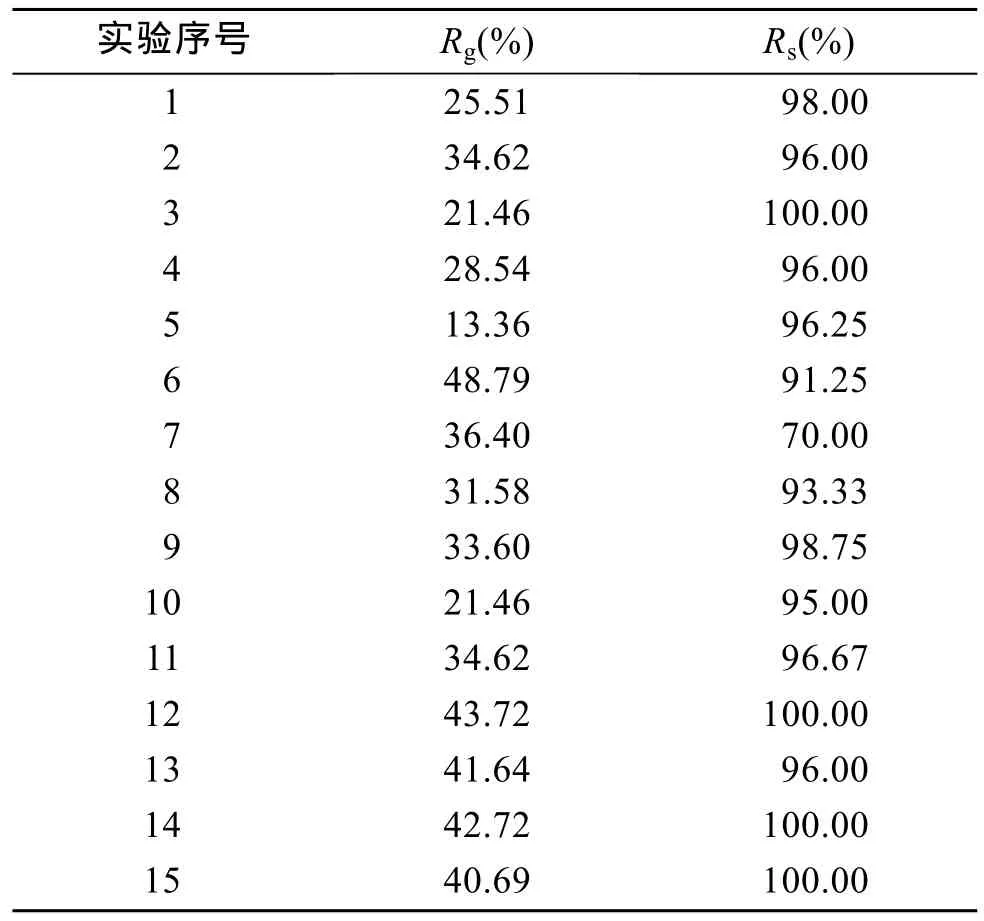
表3 响应面分析方案与实验结果Tab. 3 Experimental result and scheme of RSM
通过 SAS软件的响应面回归过程进行数据分析, 建立二次响应面回归模型, 求出最优相应因子水平。拟合得到的生长率(Y)对A,B,C的回归方程为:

2.2 回归方程系数的方差分析和显著性检验
利用SAS 9.2统计软件对Box-Behnken实验设计中的各组合实验数据进行方差分析和多元回归分析(表 4)。
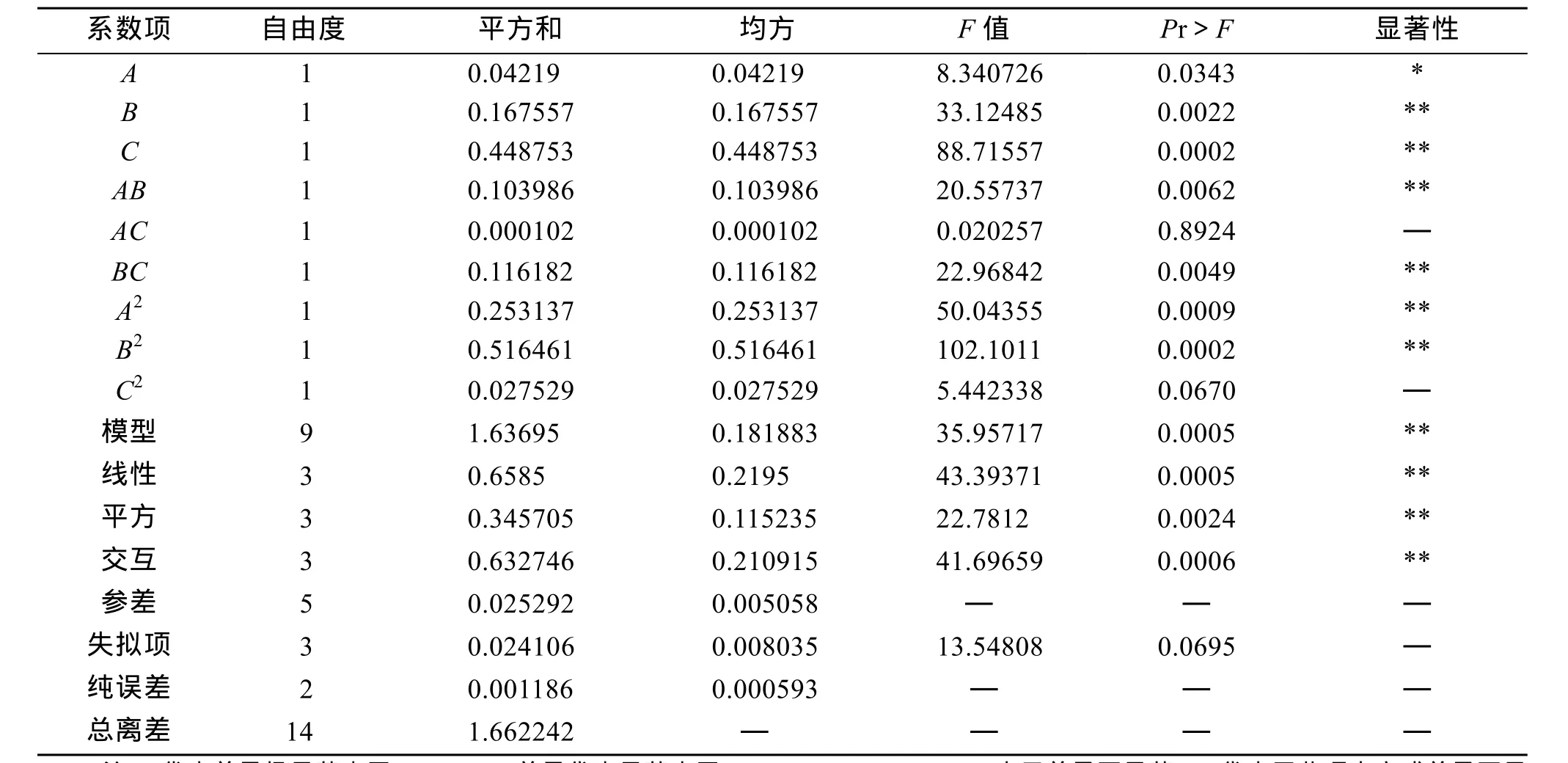
表4 二次多项式模型方差分析和多元回归分析Tab. 4 Variance analysis of quadratic equation and significance test of regression
由表4可以看出, 本实验总模型回归显著性检测P值为0.0005, 远小于0.01, 失拟项显著性检测P值为0.0695, 说明模型失拟不显著, 回归极显著, 拟合方程可靠, 线性回归(P=0.0005<0.01)、二次回归(P=0.0024<0.01)和交互(P=0.0006<0.01)都极显著。说明模型是可行的。模型的校正决定系数为R2adj=0.9574, 说明该模型可以解释95.74%影响值的变化, 只有4.26%不可以用于该模型。决定系数R2=0.9848, 说明该模型拟合程度良好, 实验误差很小, 可以用来分析和预测波纹巴非蛤幼贝生长率。从回归方程各项系数的显著性检验可知, 一次项A显著,B,C极显著; 交互项AB,BC极显著,AC不显著; 二次项A2,B2极显著,C2不显著。综合以上数据分析结果, 可以看出3种生态因子对幼贝生长率影响的大小顺序是:B>A>C。
2.3 波纹巴非蛤幼贝生长率响应面分析
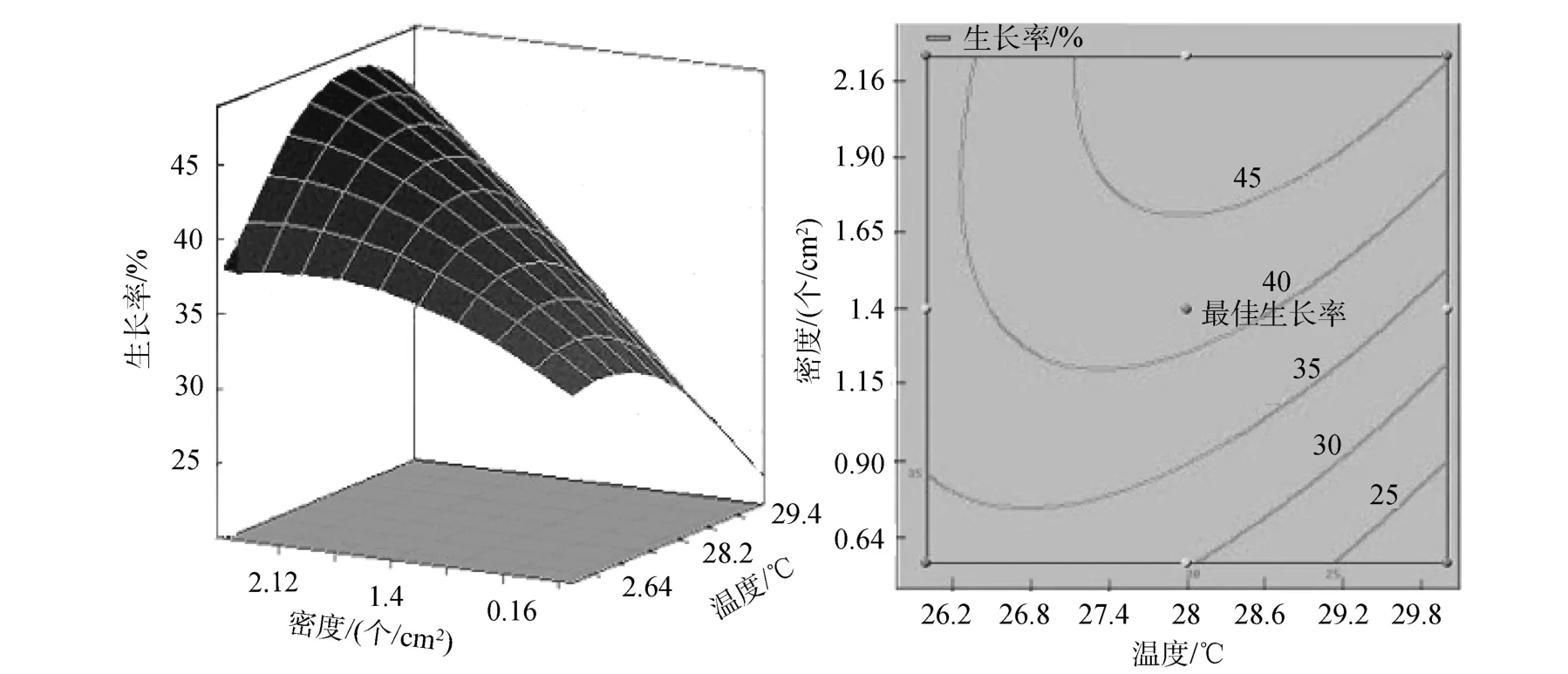
图1 密度和温度交互作用对波纹巴非蛤幼贝生长率响应面图和等高线图Fig. 1 Response surface plot and contour plot showing the interactive effect of density and temperature on Paphia undulata growth rate
三维响应面和二维等高线图可以用来评价不同因子之间的相互作用对响应值的影响。三维响应面和二维等高线图能够直观地表现出波纹巴非蛤幼贝生长率随温度、密度、盐度的不同而产生的变化趋势。温度和密度不存在显著交互作用(图1)。从温度和盐度交互作用的响应面图可以看出, 响应面分析图呈现山丘形曲面, 从等高线图看出, 等高线呈椭圆形, 温度和盐度存在显著交互作用, 生长率有极大值(图 2)。盐度和密度存在一定的交互作用(图3), 但响应面没有呈现出明显山丘形。从各图中最佳生长率相对于等高线位置的变化可以看出,盐度对幼贝生长率影响最大, 温度次之, 密度影响最小。
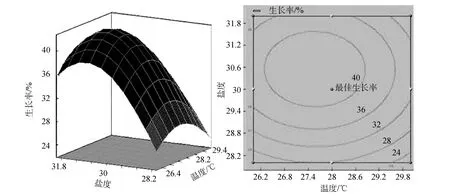
图2 盐度和温度交互作用对波纹巴非蛤幼贝生长率响应面图和等高线图Fig. 2 Response surface plot and contour plot showing the interactive effect of salinity and temperature on Paphia undulata growth rate
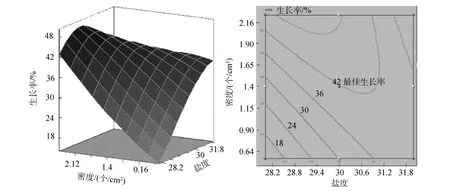
图3 密度和盐度交互作用对波纹巴非蛤幼贝生长率响应面图和等高线图Fig. 3 Response surface plot and contour plot showing the interactive effect of density and silinity on Paphia undulata growth rate
通过对SAS9.2软件模拟出的三维响应面图和等高线分析可以得出,ABC对应的编码值分别为A=-0.534,B= 0.696547,C= -0.6048, 根据回归方程中的系数计算出的实际值温度为 26.93℃、盐度为31.39、密度为 1.12个/cm2, 理论最佳生长率为41.47%。
2.4 模型验证实验结果
对模型预测的结果进行验证实验, 验证实验结果如表5所示。
将模型验证实验得出的结果与响应面预测结果进行对比(表 6)。从表中可以看出, 实验得出结果和预测结果基本一致, 生长率仅相差0.83%, 所以响应面法较为可靠。
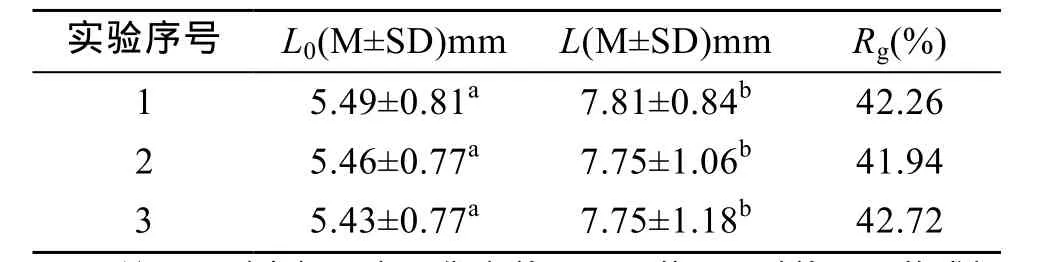
表5 实验结果验证Tab. 5 Demonstration of test result
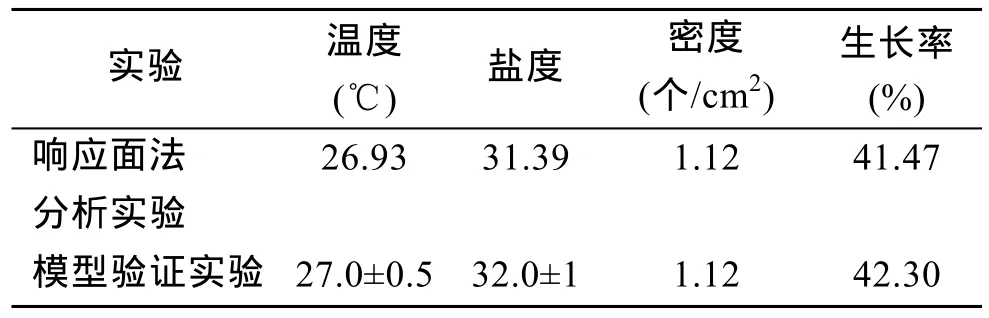
表6 验证实验结果与响应面预测结果的对比Tab. 6 Comparison of the best condition and the predicted condition
3 讨论
温度和盐度是影响海洋无脊椎动物胚胎发育和生长的重要因子, 有关温度和盐度对海洋双壳贝类的影响常见于单因子实验分析, 温度和盐度对栉江珧(Atrina pectinata)[17]、织锦巴非蛤(Paphia textzle)[18]、毛蚶(Scapharca subcrenata)[19]等的单一因素影响均有报道, 但针对温度和盐度两者对海洋双壳贝类联合作用的研究则相对较少, 目前有针对华贵栉孔扇贝(Mimachlamys nobilis)[20]的报道, 密度因子对贝类生长的影响主要通过个体之间直接对空间和食物的竞争和对环境的影响反作用于自身的生长。养殖密度对贝类生长的影响前人已做过大量的研究,例如, 青蛤(Cyclina sinensis)稚贝不同放养密度的试验结果表明[21], 青蛤稚贝的生长受到了放养密度的制约, 低密度养殖时其瞬时生长率最高, 随着放养密度的增加, 瞬时生长率逐渐下降, 死亡率上升。
虽然在单因素实验中, 温度、盐度和密度均对波纹巴非蛤幼贝生长有显著影响, 但它们之间的交互作用对幼贝生长的影响有所不同。密度与相关生态因子联合作用对贝类影响的报道也相对较少。对毛蚶的研究表明, 在适宜的范围内, 盐度和温度对毛蚶受精卵孵化和幼虫生长交互作用不显著, 但将其中一个条件设为极限时, 交互作用显著[22]。王雅倩等[23]对中国蛤蜊(Mactra chinensis)的研究显示, 个体质量和温度交互对中国蛤蜊的耗氧率和排氨率的影响显著(P<0.05)。桑士田等[24]研究了温度、盐度及pH交互作用对菲律宾蛤仔(Ruditapes philippinarum)稚贝生长的影响, 结果表明, 温度、盐度及pH三者间对蛤仔稚贝生长无显著交互作用。
本文对波纹巴非蛤的研究发现, 温度和盐度、盐度和密度对幼贝生长影响极显著, 而温度和密度影响不显著, 说明盐度对幼贝的生长是至关重要的。波纹巴非蛤是一种暖水性、在浅海生活的贝类, 对高盐的耐受能力要强于对低盐的耐受能力。李文波[25]研究了温度对波纹巴非蛤消化酶的影响, 认为波纹巴非蛤肠淀粉酶、蛋白酶、纤维素酶的活力最适温度分别为40, 55, 30℃, 波纹巴非蛤成贝耐受的极限温度范围一般为 5~31℃, 最适温度为 20~28℃[11]。响应面预测实验表明, 温度为26.93℃时幼贝生长率最高,通过模型验证实验证实此温度下幼贝生长率达到了42.30%, 与以上研究结果基本吻合。运用响应面法进行分析, 拟合得到的生长率的二次响应面回归方程,软件模拟分析后, 波纹巴非蛤幼贝最适生长环境组合是温度为26.93℃、盐度为31.39、密度为1.12 个/cm2,理论最佳生长率为 41.47%, 经实验验证后, 得到的生长率为 42.30%, 与理论值相近, 证实了响应面模型能较好地优化波纹巴非蛤幼贝的生长条件。
4 结论
本文采用响应面法研究了温度、盐度和密度三者交互作用对波纹巴非蛤幼贝生长的影响, 取得了以下结论: (1)盐度因子对波纹巴非蛤幼贝生长至关重要, 适宜的盐度和温度对幼贝生长有极大地促进作用, 在实际养殖过程中要注意二者联合效应。(2)响应面预测结果和实验验证结果十分接近, 响应面法不仅减少多因素交互实验次数, 同时可靠地反应出各研究因子对幼贝生长的影响程度, 准确地预测了实验结果, 此方法在明确了影响因子的前提下,对分析多因子影响实验效果较好。
[1]崔艳, 江莉, 夏昆, 等. 低温条件对波纹巴非蛤营养成分的影响研究[J]. 食品工业科技, 2009, 12:227-229.
[2]张能, 方海立. 牛磺酸对链佐酶素糖尿病大鼠胰岛细胞的保护[J]. 中国糖尿病杂志, 1998, 6(1): 30-33.
[3]张水波. 东山湾云霄区波纹巴非蛤生物学特性的研究[J]. 齐鲁渔业, 2008, 25(3): 15-16.
[4]赵志江, 李复雪. 波纹巴非蛤(Paphia undulata)精子发生的超微结构[J]. 台湾海峡, 1992, 11(3):238-243.
[5]吴洪流. 波纹巴非蛤雌性生殖腺的组织学研究[J].海南大学学报: 自然科学版, 2000, 18(3):258-265.
[6]吴洪流, 王红勇, 王珺. 波纹巴非蛤性腺发育分期的研究[J]. 海南大学学报: 自然科学版, 2002, 20(1):41-47.
[7]陈忻, 梁海鹰, 刘爱文, 等. 以波纹巴非蛤为原料制备海洋生物活性肽[J]. 食品科学, 2008, 29(11):35-37.
[8]黄希忠, 闵志勇. 波纹巴非蛤幼蛤凝集素活性初步研究[J]. 科技资讯, 2010, 7(a): 3.
[9]林静, 吴鸾玉, 饶小珍, 等. 波纹巴非蛤(Paphia undulata)体液和肌肉凝集素的初步研究[J]. 生物技术, 2007, 17(1): 26-29.
[10]刘建勇, 吴继兴, 孙成波. 我国东南沿海 5 个波纹巴非蛤(Paphia undulata)地理种群形态差异的分析[J].海洋与湖沼, 2010, 41(1): 114-120.
[11]左江鹏, 黄胆忠, 黄雪芬, 等. 波纹巴非蛤的生物学及养殖技术[J]. 水产科技, 2009, 35(4): 16-18.
[12]吴陈州. 波纹巴非蛤海区增养殖技术[J]. 中国水产,2005, 2: 60-62.
[13]Thompson D D. Reponse surface experimentation[J].Journal of Food Processing and Preservation, 1982,6(3): 155-188.
[14]王显. 高效铅吸附菌株的筛选及其应用的研究[D].福州: 福建师范大学, 2012.
[15]王永菲, 王成国. 响应面法的理论与应用[J]. 中央民族大学学报(自然科学版), 2005, 14(3): 236-240.
[16]汪仁官. 试验设计及分析[M]. 北京: 中国统计出版社, 1998.
[17]李金碧, 龚世园, 喻达辉. 温度和盐度对栉江珧耗氧率和排氨率的影响[J]. 安徽农业科学, 2009,37(5):2016- 2018 .
[18]栗志民, 刘志刚, 韩伟贤. 织锦巴非蛤稚贝盐度适应性研究[J]. 海洋科学, 2011, 35(10): 96-102.
[19]施祥元, 尤仲杰, 沈伟良, 等. 盐度对毛蚶稚贝生长和存活的影响[J]. 水产科学, 2007, 26(10): 554-556.
[20]刘志刚, 刘建勇, 杨博. 温度与盐度对华贵栉孔扇贝幼贝存活与生长的互作效应研究[J]. 海洋科学, 2011,35(10): 75-80.
[21]李长松, 房斌, 王慧, 等. 青蛤稚贝放养密度与底质中硫化物相关性研究[J]. 水产学报, 2006, 30(6): 796-800.
[22]沈伟良, 尤仲杰, 施祥元. 温度与盐度对毛蚶受精卵孵化及幼虫生长的影响[J]. 海洋科学, 2009, 10(4): 5-8.
[23]王雅倩, 赵文, 程芳晋, 等. 体重和温度对中国蛤蜊耗氧率和排氨率的影响[J]. 大连水产学院学报, 2009,24(6): 544-550.
[24]桑士田, 闫喜武, 杨鹏, 等. 菲律宾蛤仔稚贝最适生长环境条件的响应面法分析[J]. 水产学报, 2012,36(9): 1410-1418.
[25]李文波. 波纹巴非蛤(Paphia undulata)消化酶及蛋白质组学研究[D]. 福州: 福建师范大学, 2012.
