不同品种葡萄1年生枝的导管分子形态
2013-10-10齐永顺张京政谢兆森
王 浩,齐永顺,张京政,谢兆森
(河北科技师范学院园艺科技学院,河北 秦皇岛,066600)
植物的组织结构与功能是密切相关的,在葡萄抗寒性研究中发现,不同抗寒性葡萄枝条结构差异明显,而组织结构失水是影响葡萄抗寒性的重要因素。地上部分水分的保持对于葡萄安全越冬十分重要,冬季葡萄地上部分水分强烈散失容易使茎因失水发生冻害。植物茎蒸腾所散失的水分直接或间接地来自茎木质部导管[1]。导管分子是植物木质部输导水分和矿质营养的基本结构单位,其直径的大小对木质部水分输导效率具有重要影响[2]。分别对秦皇岛地区抗寒葡萄(不需埋土防寒越冬)和非抗寒葡萄(需埋土防寒越冬)1年生枝条导管分子进行观察,比较导管结构和类型差异,以期为葡萄枝条导管分子与葡萄抗寒性关系研究提供参考。
1 材料与方法
1.1 试验材料
试验材料由河北科技师范学院园艺实验站提供,2011年11月15日分别取长势、粗度一致的贝达、龙紫宝、北玫、马瑟兰、梅鹿辄、赤霞珠6个葡萄品种(系)的1年生枝进行试验。
1.2 试验方法
田间试验于2012年在河北科技师范学院园艺试验站进行,室内试验在园艺实验室进行。
1.2.1 葡萄品种抗寒性鉴定 6种试验材料分别在0,-15,-25,-35℃低温条件下处理12 h。然后将材料进行沙藏处理[3],第2年春季进行扦插试验,采用随机区组排列,每个处理扦插30株,重复3次,统计成活率。
1.2.2 导管观察 将1年生枝条第4节间和第5节间木质部切削成长1 cm,粗0.5 mm的小条,充分混匀;采用郭学民等[4]解离制片方法制片,在显微镜下观察并照相。随机选取100个导管分子,测量其长度、直径、两端斜度,并统计两端尾数。采用李正理[5]石蜡切片法对1年生枝进行横切,观察横截面导管分布。
以上数据应用DPS v3.01专业版数据处理系统进行统计分析。
2 结果与分析
2.1 葡萄抗寒性分析
扦插试验结果表明,6个葡萄品种枝条在0℃时,生根率和萌芽率均在80%以上未受低温伤害(表1);在-15℃下,龙紫宝和贝达生根率仍达到88.9%和78.8%,显著高于其他品种,而赤霞珠、梅鹿辄、马瑟兰等3个品种大幅下降到21.1%~25.6%,已经严重受冻;在-25℃时,只有龙紫宝、贝达、北玫3个品种生根;在-35℃下6个品种都没有生根。萌芽率的调查结果与生根率相似,但在0℃以下,萌芽率明显小于生根率。在-25℃时只有龙紫宝和贝达萌芽,并且龙紫宝的萌芽率显著低于贝达,分别为2.2%和9.8%。所以6个葡萄品种抗寒性由强到弱为贝达、龙紫宝、北玫、马瑟兰、梅鹿辄、赤霞珠。
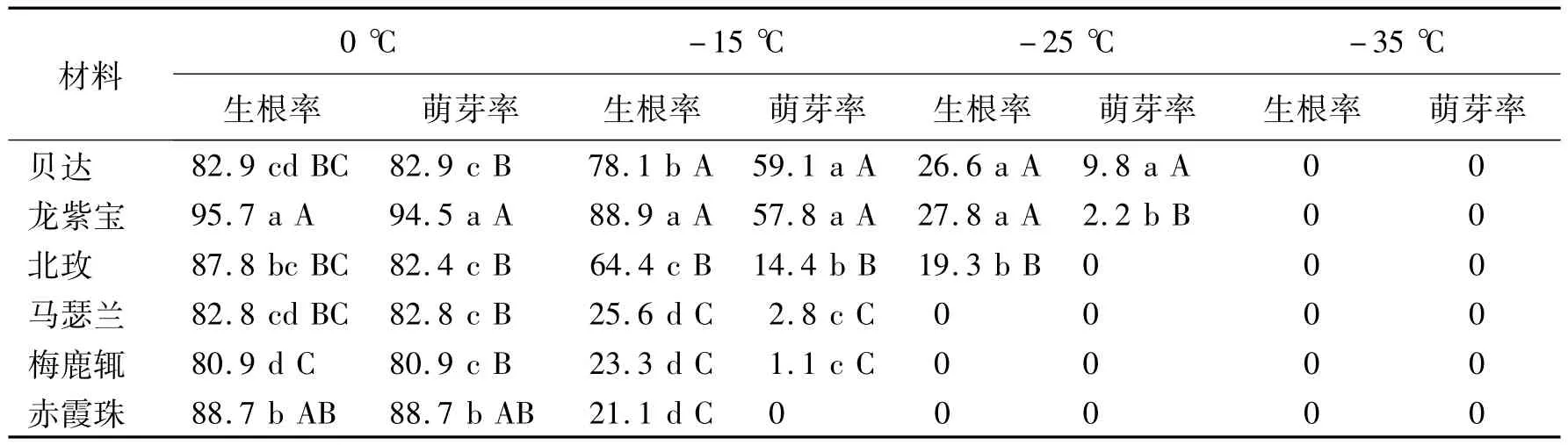
表1 低温处理后葡萄生根率和萌芽率 %
2.2 导管类型
葡萄枝条中含有多种类型导管,导管类型除了在长度和直径上有较大差异之外,根据导管两端端臂斜度,可以分为两端倾斜、一端倾斜一端水平、两端水平(斜度小于15°视为水平)3种类型;根据导管两端具尾情况,可以分为两端无尾、一端具尾一端无尾、两端具尾3种类型。离析材料中,发现多种导管类型,(1)两端有尾、两端倾斜(图1);(2)一端有尾、两端水平(图2A,图2B,图2E);(3)一端有尾、一端倾斜(图2C,图2F,图2G);(4)一端有尾、两端倾斜(图2D,图2H,图2I,图2J,图2K,图2L);(5)两端无尾、两端水平(图3A,图3B);(6)两端无尾、一端倾斜(图3D,图3E);(7)两端无尾、两端倾斜(图3C,图3F,图3G,图3H,图3I,图3J);所有导管中大部分为网纹导管(图4A,图4B,图4C,图4D),一小部分导管为螺纹导管(图4E,图4F,图4G,图4H);大部分导管两端为单穿孔板(图5B),一小部分导管为一端双穿孔板(图5A,图5C,图5D)。每一种导管在抗寒性不同的葡萄中所占比例也不相同。

图1 两端有尾导管类型
2.3 导管的长度和直径大小
分别测量6个葡萄品种枝条100个导管的长度、直径,求平均值,进行显著性分析,可以看出贝达、北玫、龙紫宝(抗寒)平均长度显著小于马瑟兰、梅鹿辄、赤霞珠(非抗寒)导管平均长度,其中龙紫宝导管平均长度最小。贝达、北玫、龙紫宝(抗寒)的导管平均直径显著大于马瑟兰、梅鹿辄、赤霞珠(非抗寒)导管平均直径,其中龙紫宝平均直径最大。
6个葡萄品种枝条中导管分子平均长度和直径差异显著(表2),龙紫宝和贝达的导管平均长度显著小于其它品种,北玫居中,马瑟兰、梅鹿辄、赤霞珠导管长度显著大于龙紫宝。在导管直径的比较中,龙紫宝导管平均直径最大,赤霞珠最小。
2.4 导管长度和直径分布
6个葡萄品种枝条导管长度在301~450 μm范围所占的比例最高,其中抗寒品种达到54.1%~65.2%,比非抗寒品种高12.8%~17.2%;在451~600 μm 和601~750 μm 范围内抗寒品种枝条导管长度都显著高于非抗寒品种,在451 μm以上时,抗寒品种所占比例为30.5%~36.3%,比非抗寒品种低16.3%~16.7%。抗寒品种中短导管所占比例显著高于非抗寒品种(表3)。
在不同抗寒性的葡萄中,不同直径导管所占比例差别较大(表3),6个品种导管直径在51~100 μm范围所占比例最高,抗寒品种为44.5%~50.7%,非抗寒品种为65.3%~72.7%,抗寒品种导管比例比非抗寒品种低20.8%~22.0%;导管直径在101~150 μm 范围内,抗寒品种导管比例为31.3%~36.9%,非抗寒品种导管比例为15.7%~25.1%,抗寒品种导管比例比非抗寒品种高11.8%~15.6%,在150 μm以上抗寒品种导管比例为9.7%~11.7%,非抗寒品种导管比例在2.9%以下。抗寒性强的品种中直径大的导管所占比例大。
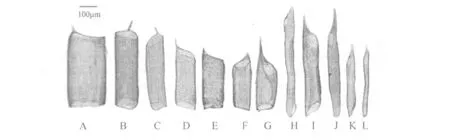
图2 一端有尾导管类型

图3 两端无尾导管类型
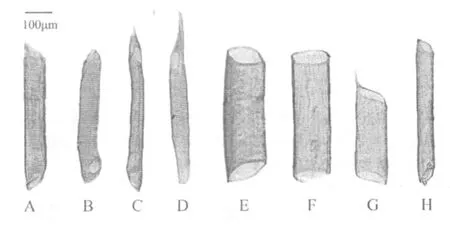
图4 网纹导管和螺纹导管类型

图5 穿孔板数量不同的导管类型

表2 导管分子的长度和直径 μm
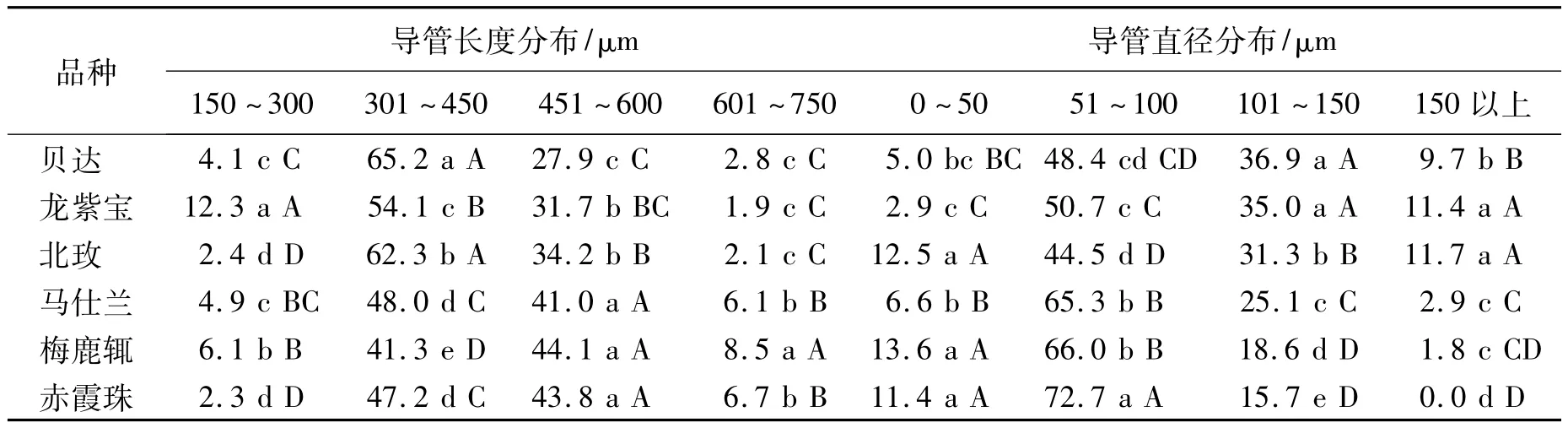
表3 导管分子的长度和直径分布比例 %
2.5 导管两端倾斜度
6个葡萄品种枝条导管两端斜度在两端水平导管类型中贝达、马瑟兰、赤霞珠导管所占比例高于其它3个品种(图6);在一端倾斜导管类型中,抗寒品种导管比例高于非抗寒品种;在两端倾斜导管类型中抗寒品种导管斜度明显小于非抗寒品种,在15~30°范围内抗寒品种导管所占比例为39%~53%,比非抗寒品种高19%~24%;在30°以上范围内抗寒品种导管所占比例为29%~36%,比非抗寒品种低23%~24% 。

图6 导管两端斜度分布情况
2.6 导管具尾数量
抗寒葡萄与非抗寒葡萄导管具尾情况差别较大,在两端具尾导管类型中,抗寒品种导管所占比例为11%~14%,非抗寒品种导管所占比例为24%~36%(图7);抗寒品种比非抗寒品种低13%~22%;在两端无尾导管类型中,抗寒品种导管所占比例为60%~70%,非抗寒品种导管所占比例为31%~43%,抗寒品种比非抗寒品种高27%~29%。抗寒品种无尾或单尾导管所占比例大。
2.7 导管的密度和分布规律
通过石蜡切片观察6种葡萄枝条横截面显微结构(图8),计算6种葡萄品种导管密度(表4)。结果表明,3个抗寒品种导管密度均显著小于3个非抗寒品种,龙紫宝和贝达导管密度最小。通过观察,横截面上的导管直径由内向外递增,抗寒品种导管并没有分布到木质部边缘,且韧皮部较厚,有3~4层质密的细胞将导管包围在木质部内;而非抗寒葡萄品种导管几乎分布到木质部边缘,韧皮部较薄,只有1~2层质密细胞。导管的这种分布规律可以提高抗寒性强的品种枝条保水能力。

图7 导管具尾情况

图8 葡萄茎的横截面(50×)
3 讨 论
3.1 导管类型多样性的形成
植物的结构和功能是密切相关的,葡萄在不同环境中经过长期进化,为适应环境变化[6],导管进化和分化程度不同,从进化角度看,导管长度短的比导管长的进化,导管直径粗的比细的进化,导管两端斜度小的比斜度大的进化,无尾比有尾进化[7]。在不同温度、水分等条件下,导管结构分化也会有所差异[8]。葡萄中含有多种类型导管,是对环境长期适应的结果。而在不同环境条件下,不同导管类型所占比例不同,可见不同类型导管在功能上是不同的。

表4 葡萄枝条中导管密度
3.2 不同抗寒性葡萄枝条导管结构差异
3.2.1 导管的长度与直径 导管的主要功能是输导水分和无机盐,从试验结果分析,抗寒品种与不抗寒品种所含导管结构比例上差异较大,从导管长度看,抗寒品种显著小于非抗寒品种,在长度分布比例上抗寒品种短导管所占比例均高于非抗寒品种;从导管直径看,无论是平均直径还是比例分布,抗寒品种导管直径均大于非抗寒品种。葡萄能否安全越冬的一个重要制约因素是水分的供应,抗寒品种的粗导管特性可以提高输水效率,在冬季和早春保证植株对水分的需要,这与肖啸、郭学民等[9]的研究结果一致。试验中还发现大多数粗导管长度都较短,这可能是因为导管的加粗生长和加长生长互相制约,两者呈负相关关系,所以抗寒性强的品种具有粗导管性和短导管性。
3.2.2 导管的两端斜度和具尾情况 导管是由又细又长的管胞进化形成的[10],管胞分子两端是封闭的,两端尖锐呈尾状,水分在管胞间的运输是通过渗透进行的,导管分子两端溶解形成穿孔板,如果分化导管分子的细胞两端溶解不彻底,导管分子形成后两端穿孔板斜面角度就大并且具尾,两端穿孔板[11,12]斜面角度大并且具尾的导管穿孔板直径大多小于导管直径,对水分的运输有阻碍作用。研究发现两端斜度大和具尾的导管长度大,直径较小,而导管两端水平、无尾的导管长度短、直径较大,这和导管的进化程度一致。在抗寒品种中两端斜度小、具尾数量少的粗导管所占比例大,有利于水分的运输。
3.3 导管的密度和分布与抗寒性关系
导管直径大、导管分布范围小是导致抗寒性葡萄导管密度小于非抗寒性葡萄的原因,葡萄导管直径大促进水分运输的同时,必然容易失水导致抽条。试验发现抗寒品种导管并没有分布到木质部外缘,且韧皮部较厚,韧皮部内侧有3~4层质密的细胞;而非抗寒品种导管几乎分布到木质部边缘,韧皮部较薄。抗寒品种的这种结构特性能够有效防止水分的散失。这种结构可能是抗寒品种葡萄可以不埋土防寒越冬的关键因素。
4 结 论
在葡萄枝条中有多种类型导管,每种类型导管在抗寒性不同的葡萄枝条中所占比例差异显著,抗寒葡萄枝条中导管平均长度短,直径大,导管密度小,并且长度短、直径大、端壁斜度小、无尾的导管类型所占比例大。这种导管结构有利于提高水分运输效率,保证葡萄越冬期间对水分的需要。葡萄枝条中导管分子的形态差异可以作为评价葡萄抗寒性的一项指标。
[1]刘晓燕,李吉跃,翟洪波,等.从树木水力结构特征探讨植物耐旱性[J].北京林业大学学报,2003,25(3):48-54.
[2]陈树思.人心果次生木质部导管分子的观察研究[J].园艺学报,2007,34(1):7-1.
[3]谢丽芬.葡萄砧木抗寒性鉴定及抗寒机理的研究[D].内蒙古:内蒙古农业大学,2007.
[4]郭学民,肖啸,徐兴友,等.21世纪桃树次生木质部导管分子特征的观察[J].果树学报,2008,25(1):22-26.
[5]李正理.植物组织制片学[M].北京:北京大学出版社,1996.
[6]肖静,杨洪强.植物导管分子分化和形成的生理生化机制[J].植物生理学通讯,2004(2):141-145.
[7]胡蓉.花椒茎次生木质部导管分子研究[J].内江师范学院学报,2012,27(6):45-47.
[8]郑玲,张海艳,李盈盈.蕨类植物管状分子的研究进展[J].生物学通报,2012,47(12):1-3.
[9]肖啸,郭学民,刘建珍,等.桃3种砧木次生木质部导管分子性状比较[J].果树学报,2012,29(2):171-176.
[10]青争.植物的导管和筛管是怎样形成的?[J].生物学通报,1987,32(3):10.
[11]陈永哲,李正理.毛茛科植物导管穿孔板结构的比较观察[J].植物学报,1990,32(4):245-251.
[12]李红芳,田先华,任毅.维管植物导管及其穿孔板的研究进展[J].西北植物学报,2005,25(2):419-424.
