近200年来湖北涨渡湖对江湖联通变化的环境响应*1
2013-09-25张清慧董旭辉陈诗越羊向东
张清慧,董旭辉,姚 敏,陈诗越,羊向东
(1:聊城大学环境与规划学院,聊城252059)
(2:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京210008)
长江中下游平原湖区是我国乃至世界上罕见的典型浅水湖群,湖泊总面积约14073 km2,多数湖泊平均水深只有2 m左右[1].该区湖泊地处亚热带季风气候区,降水充沛,是洪水易发地.历史上,长江中下游地区大多数湖泊与长江联系密切、自然相通,保持着较高的水体交换能力.江湖水交换周期短、频率高,两者之间的能量流(水量、水位)、物质流(泥沙、污染物)、生物流和价值流不停交换,构成江湖生态系统[2-3].虽然过去长期的农业活动使集水区域的营养物质等汇入湖泊,但由于存在自然联通,江湖之间不断地进行着水体交换,湖泊水体也一直维持较好的状况.然而,自1950s以来,随着人口的迅速增长和现代化农业的发展,长期的人类活动干扰(如兴修水利、森林砍伐和湖泊围垦等)日益加剧,导致该区域湖泊渐渐失去了与长江的自然联通.如今,该区与江湖连接的湖泊只剩下3个,即洞庭湖、鄱阳湖和石臼湖.
江湖联通状况对湖泊生态系统有着重要的影响.江湖联通时,江湖之间自然相通,江湖水位齐平,水交换周期短且强度大,水体长期保持着较好的状态,湖泊生态系统平衡得以维持.江湖隔绝时,湖泊面积变小甚至干涸,换水周期变长,水质明显恶化(如富营养化),生态系统结构也随之发生明显转变,引起了湖泊水文、生态环境的根本改变,导致了生态功能和生物多样性的衰退,并已对湖泊生态系统构成了威胁[4-5].因此,江湖联通是控制湖泊水量平衡,决定湖泊生态系统结构与生态状况的一个重要因素,目前已经引起科学家的广泛关注,纷纷从长期历史文献记录、生态水文观测记录或操纵生态系统实验等角度来阐述水文联通状况与水生生态系统间的因果关系[6-7].
在缺乏长期历史记录的情况下,基于湖泊沉积物研究的古湖沼学提供了一种恢复湖泊历史过程的有效手段[8].例如,沉积物的粒度组成能指示水动力强度、泥沙来源[9-10];沉积物中氮、磷、有机质等营养物质浓度高低能反映湖泊生产力状况[11];沉积硅藻因其对环境变化敏感,能较好地反映水位、营养水平等水体状况的变化[12-13].因此,基于沉积物多种理化 生物性质的分析,能有效地提供历史时期湖泊生态系统变化的过程.在长江洪泛平原湖泊,通过巢湖和太白湖210Pb年代序列和多指标的分析(硅藻、粒度、地球化学指标等)重建了过去200年生态环境变化序列,以此揭示了不同演化阶段环境指标组合关系及生态系统对变化的水文条件的生态响应特点[6,14].如今,愈来愈多的古湖沼分析认为水文变化特征是该区湖泊生态系统变化的最重要因素之一[6].
本文以涨渡湖为研究对象,在精确定年的基础上,对涨渡湖沉积岩芯进行高分辨率的多指标分析(硅藻、元素地球化学和粒度),探讨过去200年来湖泊系统对江湖连接状态改变的响应过程,为长江中下游湖泊的环境治理,特别是湖泊水文、江湖间水利工程的实施提供科学依据.
1 研究区概况
涨渡湖位于湖北省武汉市新洲区境内(图1a),距长江约1 km,面积35.2 km2,平均深度1.2 m,最大水深2.3 m.涨渡湖属于北亚热带季风气候,年平均气温16.3℃,降雨量1150 mm,蒸发量1525.4 mm.湖区地势起伏不大,海拔多在16~21 m[15].涨渡湖主要入湖河流有南边长江、西边的倒水和东边的举水等,出流于东南隅经人工渠道排入长江.历史上,涨渡湖直通长江,江湖水位齐平,而且与周围的七湖、陶湖相通,与洪水期连成一片[16].然而,自1950s以来,由于在湖区进行大规模的围垦开发和堤坝建设,致使涨渡湖与长江失去了自然的联系,1970s围垦和水利工程建设达到高峰期,1980s基本结束,基本形成当前形状,仅湿地就被围垦了约50 km2[17].2003年,世界自然基金会(WWF)将涨渡湖区域列入长江中下游流域湿地保护示范项目.2005年,实施了以“灌江纳苗”为主要内容的季节性江湖联通[18].
2 研究方法
2.1 样品采集
2011年4月,利用重力采样器,在涨渡湖湖心最深处(30°39'N,114°42'E;图1b)采集长45 cm的柱状沉积岩芯.沉积岩芯按0.5 cm间隔现场分样,样品密封带回实验室后<4℃保存以备分析测试.

图1 涨渡湖地理位置(a)和沉积岩芯位置(b)Fig.1 Location of Lake Zhangdu(a)and its core sediment(b)
2.2 实验室分析
年代测定:采用美国EG&GOrtec公司生产的高纯锗井型探测器(HPGe GwL-120-15)在中国科学院南京地理与湖泊研究所进行210Pb和137Cs测定.采用Constant Rate of Supply(CRS)模式结合1963年峰值时标的复合模式方法[19]来获得涨渡湖年代框架.
沉积物粒度测定:用10%的双氧水去除有机质,然后利用马尔文激光光学自动粒度分析仪测定.
沉积物C、N、P测定:总有机碳(TOC)浓度用重铬酸钾容量法(外加热法)测定;总氮(TN)浓度用重铬酸钾 硫酸消化凯氏法测定;总磷(TP)浓度用高氯酸 硫酸溶钼锑抗比色法测定[20-21].
沉积硅藻鉴定:取约0.5 g沉积物,先用10%盐酸去钙质胶结物,再用双氧水去有机质,离心清洗制片.硅藻种属鉴定参照Krammer和Lange-Bertalot的分类系统[22],依据硅藻保存情况,每个样品的硅藻统计数控制在300粒以上,属种丰度按百分比表示.

图2 涨渡湖沉积岩芯210Pb和137Cs活度变化曲线Fig.2 Variation curves of210Pb and137Cs activities in the core sediment of Lake Zhangdu
2.3 数值分析
降维对应分析(DCA)是一种间接梯度分析方法,用来评估生物群落变化的主要趋势,通常,这种趋势用DCA排序第一轴、第二轴得分来描述[23].对于硅藻数据,选择至少在2个样品中出现且至少在1个样品中含量大于1%的属种,对其含量进行平方根转换.数值分析在Canoco 4.5 软件中进行[24].
3 结果
3.1 沉积年代的确定
涨渡湖沉积钻孔中在20 cm左右出现一137Cs峰值,对应于1963年的全球核试验.由于210Pb活度存在一定的波动,并且近百年来流域人类活动强度发生改变,特别是筑坝后会导致沉积速率发生显著变化,因此采用CRS模式的复合模式,即用137Cs时标对CRS计算结果进行校正的模式对钻孔沉积物进行年代计算[19].沉积岩芯中210Pb和137Cs活度随深度的变化曲线见图2,整个沉积岩芯代表过去近200年的沉积.通过 CRS模式计算的沉积岩芯的平均沉积速率为0.37 g/(cm2·a).1930年以前,沉积速率由下层至上层逐渐增加,在1930年附近出现峰值,随后降低,但在沉积岩芯表层(近20年)又呈增加趋势.
3.2 化石硅藻组合
涨渡湖钻孔沉积物中硅藻含量表明,沉积柱中以浮游类型为主,还有一些附生和底栖类型(图3).根据硅藻丰度变化可划分为3个组合带,硅藻组合特征自下而上叙述如下:
ZD1带(45~24 cm;1954年前):以浮游种Cyclotella bodanica占优势,最高含量达78%,Aulacoseira granulata为次优势种,占总含量的10% ~35%,总的特征是浮游硅藻呈现上升趋势,还有少量附生和底栖种,如Gyrosigma acuminatum、Navicula spp.、Fragilaria construens var.venter、Fragilaria brevistriata 等也经常出现,但含量极低.

图3 涨渡湖沉积柱硅藻分布Fig.3 Fossil diatom assemblages in Lake Zhangdu
ZD2带(24~5 cm;1954--2005年):本带总的特征是C.bodanica含量迅速降低,A.granulata属种在本带大量出现,附生底栖属种含量增多,一些富营养指示属种在本带出现,且呈明显增多趋势,可进一步分为2个亚带:
ZD2-1带(24~16 cm;1954 -1980年):C.bodanica含量迅速减少,A.granulata急剧增加,最高含量达58%,成为优势种.附生底栖属种 G.acuminatum、Navicula spp.、F.construens var.venter、F.brevistriata 等出现.
ZD2-2带(16~5 cm;1980--2005年):A.granulata属种含量降低,附生底栖种含量增加,一些富营养指示属种(如 C.meneghinena、A.alpigena、Nitzschia palea、Surirella minuta)出现,且含量较高.
ZD3带(5~0 cm;2005 -2011年):硅藻组合底栖种Navicula spp.增加,富营养属种A.alpigena呈减少趋势.附生属种G.acuminatum、N.palea仍然保持较高含量.
3.3 数值分析
硅藻数据的 DCA 分析显示,四个排序轴的特征值分别为 0.425、0.097、0.074、0.034,第一、二排序轴分别捕获了最大硅藻数据的变率(分别为27.9%、6.3%),代表了硅藻群落的主要变化趋势.除样点17外,其他所有样点的第一、二排序轴的排序图也将3个组合带清晰地区分开来(图4),分别对应于图3中的分带.相应地,根据硅藻的生活习性特征将其群落变化进行划分,可以看出:在ZD1带浮游硅藻呈增加趋势,而在ZD2带又开始迅速下降,非浮游类型增加(图5).
3.4 地球物理化学指标
在ZD1带总有机碳含量较低(<10%),但1950s以后呈增加的趋势;沉积物中总磷和总氮的浓度变化和TOC类似,在ZD1带含量不高,1950s后逐渐增加;粒度指标有明显变化,中值粒径在ZD1带很大(>20 μm),在1950年左右急剧下降到低值(<10 μm),之后保持稳定(约9 μm)(图 5).
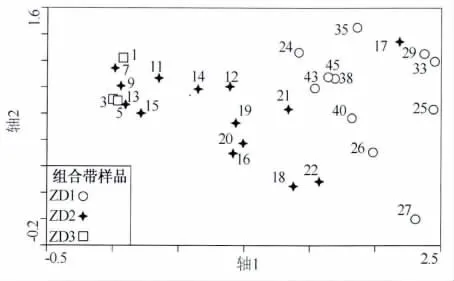
图4 沉积柱硅藻数据的DCA排序分析结果Fig.4 DCA ordination results of the diatom data of the sediment column

图5 沉积柱中浮游硅藻和非浮游硅藻含量、DCA第一和第二排序轴得分和沉积岩芯粒度、有机碳含量、总磷及总氮浓度的变化趋势Fig.5 Percentages of plankton and non-plankton diatoms of the sediment column,the first two DCA ordination axisscores,variation of median grain size,content of TOC,concentrations of TP and TN
4 讨论
涨渡湖沉积物忠实地记录了过去200余年来该湖的环境演变过程.基于沉积物的多指标分析包括硅藻、粒度、沉积物氮磷、有机碳等,呈现出基本一致的阶段性变化(图5),反映了各种生态系统各要素对环境变化过程响应的一致性.配合高精度210Pb、137Cs测年,重大环境变化事件得以重建.据历史文献记载,涨渡湖区建坝始于1954年[17].这一事件在湖泊沉积物中有着清楚的记录,比如沉积物粒径由1950s前的粗砂(>20 μm)急剧下降到稳定低值(<10 μm),其它各个沉积物理化、生物指标均出现相似的跳跃性变化.沉积记录和历史记录的相互印证,保证了利用沉积物记录来反演湖泊环境演化历史的可行性[25].
江湖联通期(1954年以前)涨渡湖与长江自然联通,两者保持着较高的水体交换能力,硅藻组合以浮游硅藻C.bodanica为优势种,它是一种低营养属种[26-27],偏好于高水位、扰动的水体状态.因此,此阶段出现的大量C.bodanica指示了营养水平低、水位高、水动力强的水体状态.另外,沉积物粒度变化可以指示浅水湖泊的水动力变化的强度,提供水文变化信息[28].1954年之前中值粒径持续高值(>20 μm),以砂质沉积物为主.考虑到1950s初期涨渡湖仍有较大面积(>200 km2),湖心处钻孔中的粗颗粒不可能是由流域物质搬运而来(大的颗粒会直接沉积在湖岸带),极有可能是由江水携带入湖,因此江河性来源的粗颗粒沉积物反映了较强的江湖水交换过程.同时,由于水交换强度大,流域内输入的营养物质无法蓄积,有机质和营养盐浓度都较低,相应地,水体营养水平也较低.因此,在江湖联通期,涨渡湖处于低营养、水位相对较深和水动力较大的水体状态.
江湖阻隔后(1954年以后),涨渡湖营养水平明显升高,生态系统出现一定程度的退化.筑坝后,江湖水体交换基本消失,湖泊换水周期变长,流域入湖营养盐得以蓄积,导致该湖营养水平逐渐升高并出现富营养化特征.硅藻组合以 A.granulata为优势种,喜好富营养环境的属种[29-30],如 C.meneghinena、A.alpigena、N.palea、S.minuta含量开始快速升高,指示了湖泊营养水平的快速上升.相应地,沉积营养要素TOC、TP、TN等浓度亦开始显著升高(图5).同时,湖泊生态系统出现一些退化信号.有文献记载,随着大坝的建设及垦殖活动的兴起,涨渡湖淤积严重;鱼类多样性骤减,由1951年的82种下降到2003年的46种[17];随着营养水平的进一步上升,水生植被的物种数也出现退化,由筑坝初期的多种水草群落演化到棱角为主的单调群落[31].
A.granulata是江湖阻隔后初期含量较高的一个特征属种.该属种主要分布在中-中富营养水体中[29,32];在长江洪泛平原地区,适宜在70 μg/L左右总磷浓度下生长[30];同时它硅化程度高、有较高沉降率且偏好于扰动强烈的水体以保证其悬浮于水柱中.例如,Owen等在上升流强烈的美国Malawi湖中发现Aulacoseria属种含量丰富[33],Pilskaln等还在该湖中发现Aulacoseria适合在干燥风强的季节和较高营养、高扰动的水体状态[34].最近还有研究表明A.granulata属种对水动力强度和风的强度较敏感[35].涨渡湖是长江中下游地区一个典型的浅水湖泊,容易受到风浪作用的影响,水动力扰动大,再加上筑坝使得湖泊换水周期变长、水体透明度降低、营养蓄积,适宜A.granulata的生存.不仅如此,在调查的49个长江中下游浅水湖泊的表层沉积物中,Aulacoseria属也是大多数湖泊中含量较高的属种.因此,在1954年以后出现的大量A.granulata可能反映出浅水湖泊所特有的这种易受风浪影响、混浊和营养蓄积的水文条件.江湖水体交换减少,蓄水周期增加,从而导致细颗粒无法冲刷到长江而集聚在湖泊中.在过水性浅水湖泊中细颗粒增多往往代表水动力条件的减弱[36].也有研究表明水的流速与细颗粒呈负相关[10].随着湖坝对涨渡湖与长江水的拦截,水体交换减弱,中值粒径急剧下降,细颗粒增多,表明筑坝后水动力条件减弱.因此,在江湖阻隔期,水动力条件减弱,营养蓄积并出现富营养化.
C.bodanica是长江中下游地区湖泊江湖联通的一个潜在的指示性属种.如前文所述,1954年前江湖联通时期该种含量持续较高,反映了低营养、高水位、过水性的水体状态.该属种在涨渡湖筑坝后亦有少数年份出现较高含量,如17 cm样品(对应于1980s初期)中含量达22%,在DCA排序分析中被归入江湖联通期的样品组合之中.该异常样点可能指示了当年较大规模的洪水[37]或人为开闸导致江湖短暂联通,在合适的环境条件下C.bodanica得以大量生长,否则,在1980s后该湖营养较高、水位低的背景下,C.bodanica根本无法繁殖.
1980年以后,A.granulata含量持续下降,附生、底栖硅藻种增多.附生硅藻增多通常与水生植物大量发育有关[6].1970s湖泊围垦和水利工程建设达到高峰期、人类活动的加强输入更多的营养物质及围垦所导致的湖泊淤积变浅,这些因素都导致水生植物的大量生长.同时,水生植物生长发育可以吸收部分营养盐,对悬浮物质具有吸附和净化等功能,能使总磷保持低值.因此,在1980年后,虽然营养蓄积速率加快,但是由于水生植物的大量生存,附生、底栖硅藻属种相应增多,该湖营养水平有所上升,但水体仍然没有恶化.底栖种的增多是浅水湖泊的特征.长江洪泛平原地区是一个典型的浅水湖群区,但涨渡湖与长江洪泛平原地区的其它浅水湖泊不同,涨渡湖水很浅,且水深趋于一致.如龙感湖平均水深3.78 m,最大水深4.58 m;太白湖平均水深3.2 m,最大水深 3.9 m;巢湖平均水深 2.7 m,最大水深 5.1 m[1].而涨渡湖目前常年平均水深1.3~1.5 m,最大水深也只有2.3 m,因此比较适合底栖种生存.
涨渡湖季节性通江后(2005年后),湖泊水质状况有所好转.据文献报道,涨渡湖2005年实施“灌江纳苗”的季节性江湖联通后,银鱼等9种曾经消失的野生鱼类重返涨渡湖,随后的监测也表明湖泊水质改善[38-39].相应地,硅藻群落组合亦发生明显变化,富营养属种A.alpigena含量显著降低,底栖种Navicula spp.含量明显上升,表明水体透明度变高,更多的光线抵达湖底,有利于底栖硅藻属种的生长[40].附生种含量无显著变化,表明该时期水生植被未发生较大变化.其他指标如TOC含量以及沉积物TP、TN浓度亦无显著变化,表明流域内仍然有较高的营养输入.这些古湖沼学记录揭示了短期的江湖联通尽管对水质有一定改善,但湖泊的彻底治理可能要依赖于长期、更为有效的江湖互通.
涨渡湖过去近200年的历史记录证实了“江湖联通”是缓解长江中下游地区湖泊环境压力的有效手段.事实上,针对该区湖泊普遍面临的富营养化、污染问题,早有学者提出引入长江之水来改善湖泊水质,并且自2002年开始在国家水利部的统一部署下正式启动了“引江济太”调水工程[41].然而,由于太湖水质影响因素众多,尽管太湖水质近年有所好转,但各界专家对这一工程的效果褒贬不一,仍缺乏可信的证据.涨渡湖过去近200年的历史记录清楚地揭示了该湖筑坝后水面迅速萎缩、蓄水周期增加、营养富集、生态环境逐步恶化,实现江湖联通后水质出现一定好转.因此江湖联通不仅有利于江湖水体交换,而且对水资源保护、水生生物保护以及湖泊生态保护发挥着重要作用.通过江湖的联通,水体的交换强度增加、蓄水周期缩短、自净能力增强,从而水体的水质状况在一定程度上得到改善.同时,江湖联通可恢复江湖之间的生态通廊,促进江湖之间的水生生物交流,实现生态系统的健康稳定[7].
5 结论
1)结合放射性定年,涨渡湖沉积柱高分辨率的多指标分析(硅藻、地球化学指标和粒度)揭示了过去近200年来该湖生态系统对江湖联通关系改变的响应过程.
2)江湖联通状况的改变,通过湖泊的换水周期、营养状态和光照条件等因素的改变影响着湖泊生态系统:江湖联通时,湖泊系统长期保持着健康的状态;江湖隔绝时,蓄水周期增加,营养易富集,生态环境也易随之恶化.
3)涨渡湖过去近200年的历史记录表明,江湖联通有利于江湖水体交换,对水资源保护、水生生物保护以及湖泊生态保护发挥着重要作用,是缓解湖泊生态环境压力的有效手段.
致谢:中国科学院南京地理与湖泊研究所姚书春博士、陶玉强博士和桂志凡博士协助野外采样;夏威岚副研究员提供放射性测年结果,在此一并感谢.
[1]王苏民,窦鸿身.中国湖泊志.北京:科学出版社,1998.
[2]陈 进,黄 薇.长江流域水资源配置的思考.水利发展研究,2005,5(12):14-17.
[3]李长安.从地学规律和可持续发展看长江分蓄洪区建设.长江流域资源与环境,2005,14(1):6-11.
[4]熊绍钧.长江中游涨渡湖的水平衡分析与研究.水利水电快报,2006,27(24):67-71.
[5]Yin H,Li C.Human impact on floods and flood disasters on the Yangtze River.Geomorphology,2001,41(2/3):105-109.
[6]Liu Q,Yang X,Anderson NJ et al.Diatom ecological response to altered hydrological forcing of a shallow lake on the Yangtze floodplain,SE China.Ecohydrology,2011,5(3):316-325.
[7]雷阿林,李志军,贾海燕等.重建江湖动态联系 修复水网生态环境.人民长江,2004,35(9):1-4.
[8]王苏民,张振克.中国湖泊沉积与环境演变研究的新进展.科学通报,1999,44(6):579-587.
[9]Okuda S,Imberger J,Kumagai M.Physical processes in a large lake:Lake Biwa,Japan.Washington,DC:American Geophysical Union,1995.
[10]Molinaroli E,Guerzoni S,De Falco G et al.Relationships between hydrodynamic parameters and grain size in two contrasting transitional environments:The Lagoons of Venice and Cabras,Italy.Sedimentary Geology,2009,219(1):196-207.
[11]羊向东,王苏民,沈 吉等.近0.3 ka来龙感湖流域人类活动的湖泊环境响应.中国科学:D辑:地球科学,2001,31(12):1031-1038.
[12]Sokal MA,Hall RI,Wolfe BB.Relationships between hydrological and limnological conditions in lakes of the Slave River Delta(NWT,Canada)and quantification of their roles on sedimentary diatom assemblages.Journal of Paleolimnology,2008,39(4):533-550.
[13]Wiklund J,Bozinovski N,Hall R et al.Epiphytic diatoms as flood indicators.Journal of Paleolimnology,2010,44(1):25-42.
[14]Chen X,Yang XD,Dong XH et al.Nutrient dynamics linked to hydrological condition and anthropogenic nutrient loading in Chaohu Lake(southeast China).Hydrobiologia,2011,661:223-234.
[15]李黔湘,于秀波,李家永.湖北省涨渡湖流域湿地变化及其动因分析.长江流域资源与环境,2005,14(5):600-604.
[16]吴 寒.基于RS和GIS的涨渡湖天然湿地近20年演化研究.安徽农业科学,2008,36(14):6050-6052.
[17]王利民,胡慧建,王 丁.江湖阻隔对涨渡湖区鱼类资源的生态影响.长江流域资源与环境,2005,14(3):287-292.
[18]朱 江,王利民,雷 刚.重建江湖联系保护涨渡湖湿地.人民长江,2005,36(11):60-62.
[19]刘恩峰,薛 滨,羊向东.基于210Pb与137Cs分布的近代沉积物定年方法——以巢湖、太白湖为例.海洋地质与第四纪地质,2009,29(6):89-93.
[20]中国科学院南京土壤研究所.土壤理化分析.上海:上海科学技术出版社,1978:62-67.
[21]薛 滨,姚书春,王苏民等.长江中下游不同类型湖泊沉积物营养盐蓄积变化过程及其原因分析.第四纪研究,2007,27(1):122-127.
[22]Krammer K,Lange-Bertalot H.Baeillariophyceae(l-4 Teil).In:Ettl H,Gerloff J,Heynig H et al eds.Susswasserflora von Mitteleuropa.Stuttgart and Jena:Gustav Fischer Verlag,1986 1991,2(1/2/3/4).
[23]Ter Braak CJF,Van Tongeren O.Data analysis in community and landscape ecology.Cambridge:Cambridge University Press,1995.
[24]Ter Braak CJF,Smilauer P.CANOCO Reference Manual and CanoDraw for Windows User's Guide:Software for Canonical Community Ordination(version 4.5).Ithaca,N.Y.Microcomputer Power,2002.
[25]Smol J.Pollution of lakes and rivers:a paleoenvironmental perspective.Oxford:Wiley-Blackwell,2008.
[26]Bradbury P,Cumming B,Laird K.A 1500-year record of climatic and environmental change in Elk Lake,Minnesota III:measures of past primary productivity.Journal of Paleolimnology,2002,27(3):321-340.
[27]Stoermer EF,Emmert G,Julius ML et al.Paleolimnologic evidence of rapid recent change in Lake Erie's trophic status.Canadian Journal of Fisheries and Aquatic Sciences,1996,53(6):1451-1458.
[28]Ashley GM.Interpretation of polymodal sediments.The Journal of Geology,1978,86(4):411-421.
[29]Van Dam H,Mertens A,Sinkeldam J.A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands.Aquatic Ecology,1994,28(1):117-133.
[30]Yang XD,Anderson NJ,Dong XH et al.Surface sediment diatom assemblages and epilimnetic total phosphorus in large,shallow lakes of the Yangtze floodplain:their relationships and implications for assessing long-term eutrophication.Freshwater Biology,2008,53(7):1273-1290.
[31]李立银,倪朝辉,李云峰等.涨渡湖湿地保护与渔业生产优化模式探讨.长江流域资源与环境,2006,15(3):366-371.
[32]Gómez N,Riera JL,Sabater S.Ecology and morphological variability of Aulacoseira granulata(Bacillariophyceae)in Spanish reservoirs.Journal of Plankton Research,1995,17(1):1-16.
[33]Owen R,Crossley R.Spatial and temporal distribution of diatoms in sediments of Lake Malawi,Central Africa,and ecological implications.Journal of Paleolimnology,1992,7(1):55-71.
[34]Pilskaln CH,Johnson TC.Seasonal signals in Lake Malawi sediments.Limnology and Oceanography,1991,36(3):544-557.
[35]Stager JC,Hecky RE,Grzesik D et al.Diatom evidence for the timing and causes of eutrophication in Lake Victoria,East Africa.Hydrobiologia,2009,636(1):463-478.
[36]殷志强,秦小光,吴金水等.湖泊沉积物粒度多组分特征及其成因机制研究.第四纪研究,2008,28(2):345-353.
[37]赵 艳.关于长江中游洪灾问题的思考.地学前缘,2000,7(B08):87-93.
[38]严 黎,陈 立,吴门伍等.浅析灌江纳苗对涨渡湖湿地水质及泥沙的影响.水资源与水工程学报,2006,17(5):38-41.
[39]陈 勇.在全国率先建立江湖连通机制长江水首入涨渡湖.湖北日报,2006 -07 -08(001).
[40]Bennion H.Surface-sediment diatom assemblages in shallow,artificial,enriched ponds,and implications for reconstructing trophic status.Diatom Research,1995,10(1):1-19.
[41]夏 军.引江济太工程与太湖流域水资源可持续利用刍议.中国水利,2004,(2):32-35.
