伊洛河流域外来草本植物分布格局
2013-09-07郭屹立丁圣彦卢训令苑晓雯郭静静李乾玺
郭屹立,丁圣彦 ,苏 思,卢训令,苑晓雯,郭静静,李乾玺
(1.河南大学环境与规划学院,开封 475004;2.河南大学生命科学学院,开封 475004)
外来种指由于人类有意或无意的活动被带到其自然演化区域以外的物种[1]。在人类频繁的经济活动推动下,许多外来种在新的分布区建立种群,并给当地生态系统、人类健康及农林业生产等造成了巨大影响,进而成为入侵种[2-3]。生物入侵不仅可以降低生物多样性,同时还威胁着全球的生态环境和经济发展[4-5]。
构建一个国家或地区的涵盖基本生物学和生态学信息的外来物种数据库对于研究生物入侵至关重要[6-7]。这不仅有助于人们认识生物入侵的分布格局,研究影响生物入侵的重要因素[8],同时也是制定外来入侵生物管理政策、建立环境风险评估系统的早期工作的重要一步[9]。全球范围内约有395种入侵植物获得了详细研究,占入侵物种数目的44%[10]。目前研究者的目光主要聚焦在入侵种上,然而,形成入侵并造成危害的入侵种仅占外来种的很小一部分[11]。因此构建一个比较健全的外来种数据库是当前急需要填补的一个空白[12]。
伊洛河流域地处半湿润、半湿润偏旱地区,是黄土边缘地带和山地平原交接地带,具有典型的生态过渡带特征[13]。该区地处黄河流域的中下游,是我国原始农业发展最早地区之一,是中华民族的发祥地之一,有着悠久的历史和灿烂的文明。而物种的入侵规模与区域经济、人口密度、交通流量等呈正相关[14-15]。随着当地农业、经济、旅游业的发展和人口的膨胀,其与外界的交流也日益频繁,一些外来种也逐渐扩散,甚至一些恶性入侵种(如空心莲子草)开始肆虐扩张,危害着当地农业等生态系统[13,16]。
目前,国内学者就伊洛河流域的水文[17]、地貌[18]、景观格局动态变化[19-20]、流域内草本植物物种多样性[13,21]和功能群划分[16]等方面做了大量的研究工作。良好的水热条件再加上人为干扰及洪水等引起的自然干扰,使河流湿地成为可入侵性较强的生态系统。然而,有关该流域内外来种的组成、引入方式、分布格局及对当地生态环境的影响情况尚未见报道。基于前人研究及《河南植物志》等信息的收集整理,以伊洛河流域洛河段为研究区域,沿流域自上到下分别选取3种具有代表性的生境类型:河滩地、农田、林地。运用群落生态学中的样方法对流域内典型生境类型中的外来草本植物进行调查,探讨流域内外来草本植物种类组成、重要特征和引入方式等,及其在不同生境类型中的分布差异和流域纵向梯度上的分布格局,进而构建一个比较健全的该流域内外来物种数据库。以期为流域内外来种的入侵风险评价和扩散监测奠定基础,并为制定合理的生态系统保护策略、控制外来种危害和防止其进一步蔓延扩散提供依据。
1 研究区域概况
伊洛河是黄河中游南岸的一条重要水系(图1),它位于109°43'—113°10'E,33°39'—34°54'N之间。年均气温7.8—13.9℃,最低气温-21.6—-11.8℃,最高气温37—40.8℃。年均降水量710—930 mm。
本研究以流域内洛河干流及伊河和洛河合流后的伊洛河为研究区域。洛河源出于陕西省洛南县洛源乡木岔沟,向东流入河南境内,经卢氏县、洛宁县、宜阳县、洛阳市,在偃师市杨村附近与伊河合流后称伊洛河,在巩义市洛口北汇入黄河,全长453 km。长水以上为上游,洛河与伊河合流后为下游。该流域地处北亚热带向暖温带过渡带,景观类型上从山地经丘陵向平原延伸。自然植被以暖温带落叶阔叶林为主,间有亚热带区系的植物成分,如黄连木(Pistacia chinensis)、黄栌(Cotinus coggygria)、山胡椒(Lindera glauca)、牛鼻栓(Fortunearia sinensis)、美丽胡枝子(Lespedeza formosa)等[20]。
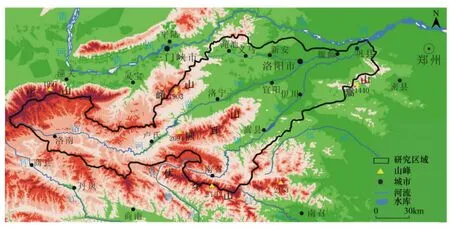
图1 伊洛河流域地形图Fig.1 A topographic map of the Yiluo River basin
2 研究方法
2.1 样地设置
在2008年前期实地踏查的基础上,于2009年8—9月,在整个伊洛河流域洛河段内沿主河道,参考海拔高度并结合行政区选取典型样地。在上游山地的洛南县和卢氏县,自海拔1227 m至528 m的河道上,选择具有代表性的地段,每隔海拔50 m确定一个样地;中游丘陵地区的洛宁县和宜阳县,自海拔375 m至188 m的河道上,每隔海拔20 m确定一个样地;下游平原地区的偃师市和巩义市(海拔118 m至101 m),结合行政区域,沿主河道每间隔约10 km设置一个样地。共计39个样地(表1)。每个样地内选取3种生境类型,分别为河滩地、农田和林地,其中河滩地距河道较近,受河流影响最大,农田与林地次之。农田为夏秋季节最常见的玉米地,选定作物生长期保持在果穗形成但尚未成熟期;林地选取人工杨树林,树龄及种植密度具有代表性林地;河滩地选取河流沿岸不受挖沙等强烈人为干扰的地段。每种生境分别设置一个20 m×20 m的样方,在样方的四角及中央分别设置1个1 m×1 m的草本样方进行调查观测。最终共设置小样方530个,河滩地和农田均为195个,林地140个。调查样地内的外来物种种类、出现个体数,盖度,株高等(文中海拔、经纬度均以河滩地为准)。
2.2 外来草本植物调查及选取
在2008年沿线踏查的基础上于2009年8—9月,调查伊洛河流域内的外来草本物种,并采集标本带回鉴定。参考《中国植物志》、《中国杂草志》、《河南植物志》、《中国外来入侵种》等书籍,结合中国数字植物标本馆(http://www.cvh.org.cn/cms/)鉴定流域内的外来草本植物种类。选取鉴定出的27种外来草本植物(表2)为研究对象。
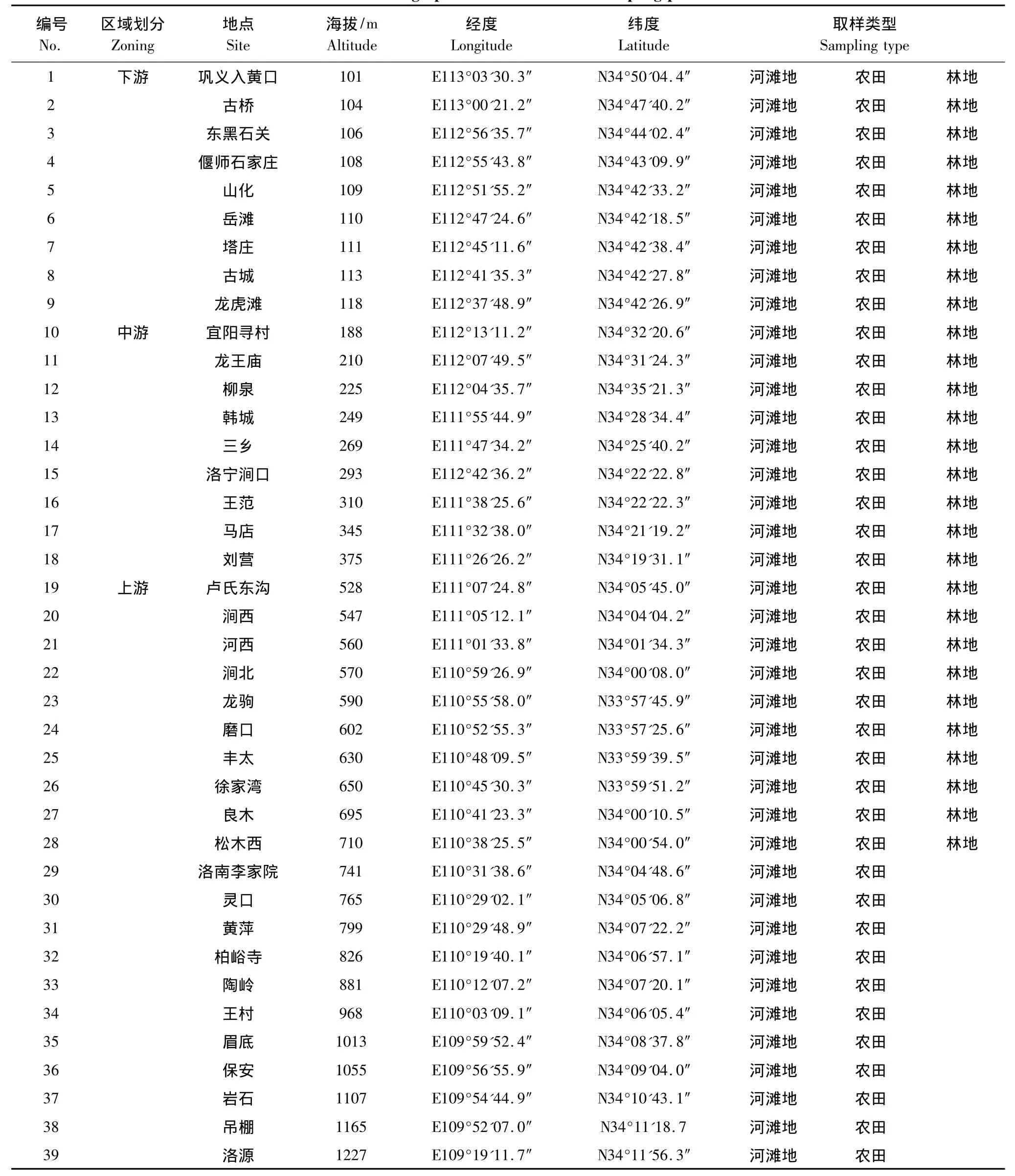
表1 样地概况Table1 Geographical conditions of the sampling plots
2.3 数据分析
采用重要值作为多样性指数的计算依据,其计算公式如下:

生境间所有外来物种重要值的和不具有方差齐次性,故使用非参数检验。流域横向梯度(不同生境间)总体差异性用Kruskal-Wallis秩和检验,两两间差异性用Wilcoxon秩检验。纵向梯度(沿流域自下而上)分布的39个样地代表不同的环境因子(如海拔、水热条件、人类扰动等)形成的环境梯度的综合。文中编号大小仅表示不同环境因子的综合,代表流域纵向梯度下的环境梯度。比如上游地区海拔差异明显,且人为干扰较小。而下游地区海拔差异很小,但人为干扰强烈。本文依据外来种重要值的大小在流域尺度内横向和纵向梯度的比较,以及不同外来种在不同生境间的分布差异,进而确定流域内外来草本植物的分布格局。
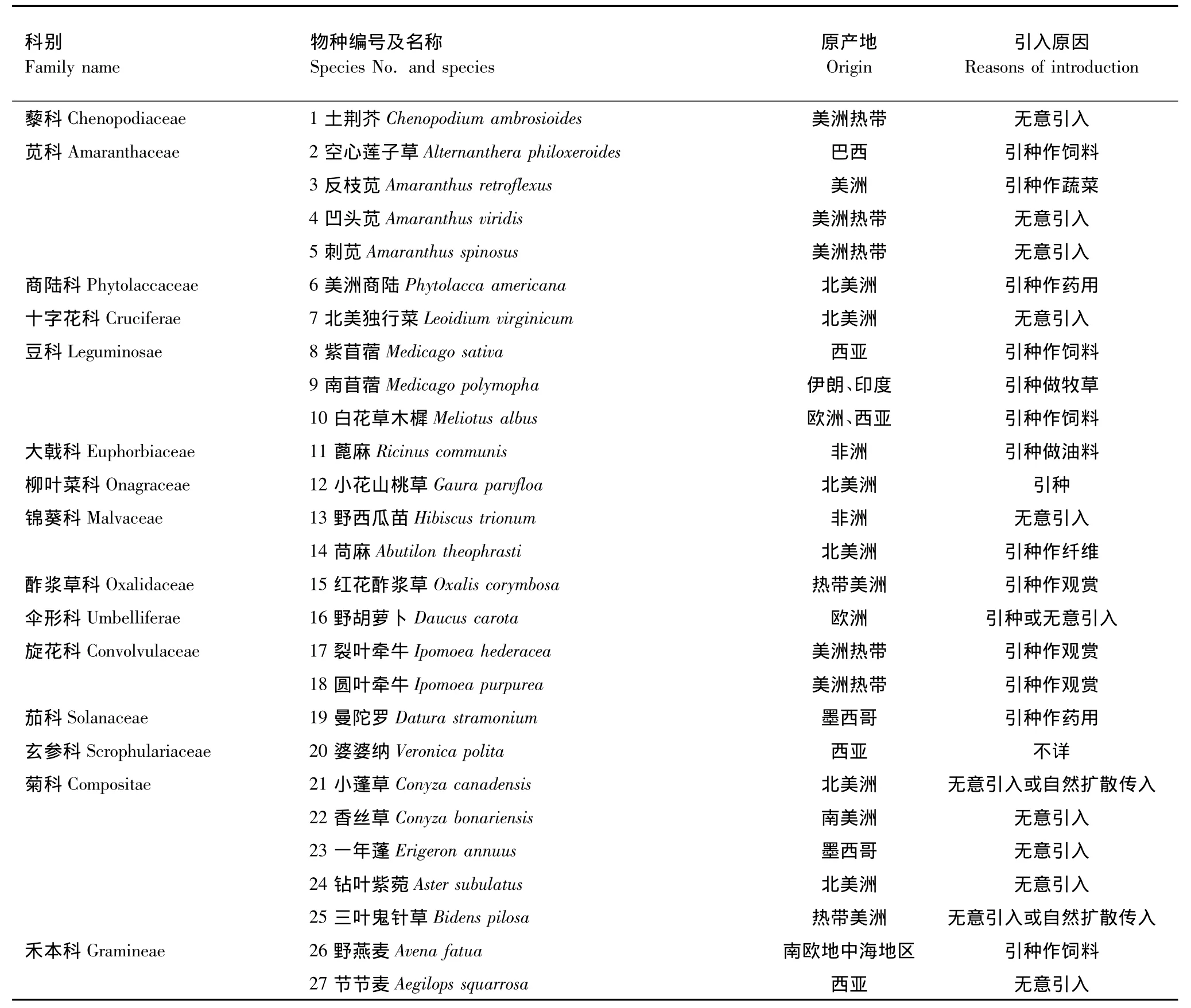
表2 伊洛河流域外来草本植物名录Table2 List of alien invasive plants in the watershed of the Yiluo River
3 结果
3.1 流域内外来种概况
通过2008、2009年夏秋季对伊洛河流域实地调查和对相关文献的查阅,初步确定伊洛河流域有外来草本植物15科27种。以菊科、苋科和豆科分布最多,分别有5种、4种和3种。其中18种来自美洲,占所有物种的67%,其余外来草本植物来自于欧洲、西亚等地。引入方式以有意引种和无意引入为主,自然扩散进入只有两种(表2)。
3.2 伊洛河外来草本植物横向分布格局
运用所有外来种在样方中的重要值作为指标,生境间外来种差异不显著(Kruskal-Wallis test,chi-squared=2.2449,d f=2,P=0.3255),且各生境间差异不显著(Wilcoxon test,P>0.05)(图2)。各生境重要值的平均值可以看出,河滩地外来种分布最多,且样方间变化最大;农田分布最少。
不同外来种横向分布(不同生境间)存在明显差异(表3)。如苘麻主要分布于农田中,依赖人畜活动进行传播、扩散;小花山桃草、钻叶紫菀和空心莲子草则主要分布于河滩地生境类型,依赖水路进行传播;红花酢浆草、圆叶牵牛、牵牛和紫苜蓿主要分布于林地。由于选取样地皆为人工杨树林地,且绝大部分沿河道两旁和公路旁分布,认为此类外来种是依靠道路进行扩散;相思草、小蓬草和一年蓬等较均匀地分布在河滩地与林地中。空心莲子草仅分布于下游平原地区,在河滩地大面积分布,但在部分农田和林地均发现有一定分布。
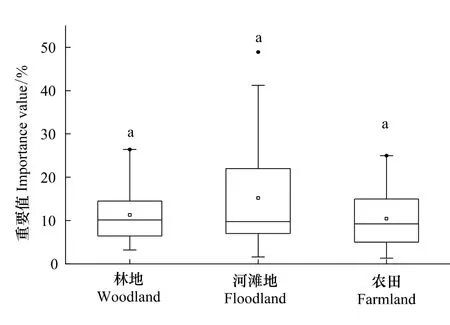
图2 外来草本植物沿伊洛河流域横向分布格局差异性分析Fig.2 Difference analysis of distribution patterns of alien herbs transverse direction of the Yiluo River watershed

表3 伊洛河流域外来草本植物不同生境间分布比较Table3 Comparison of alien invasive herbs distribution pattern in different habitats of the Yiluo river watershed
3.3 伊洛河外来草本植物纵向分布格局
流域内不同生境下外来种沿纵向梯度(河流走向)变化趋势如图3。
(1)河滩地生境二次拟合曲线模型显示从入黄河口往上,外来种出现个体数呈逐渐下降趋势。其中,入侵种空心莲子草仅在下游地区沿河道成片分布;钻叶紫菀广泛分布整个流域河滩地生境内(表3)。
(2)林地生境从入黄河口向上,外来种出现个体数呈下降趋势并在中上游交汇处达到最小值。自样地28之后,地处流域的上游山区,未发现有人工林地成片分布。山体多由栎类和油松(Pinus tabulaeformis)所组成的天然次生林所覆盖,这些是在原有自然林被砍伐的基础上自然恢复生长的次生林地,林地内很少发现有外来种分布。
(3)农田生境整体变化趋势与河滩地相同,即在下游地区最多,上游最少的分布格局。其中,样地1和样地2中有大量空心莲子草分布。
3.4 其他分布格局
研究未能对流域内进行更多生境类型及更加精细划分的外来种调查,仅对3种常见生境中可观察到的外来种进行记录。表3仅列出了流域中出现次数较多且重要值较大的物种,而个别出现较少的稀有种,如蓖麻分布在中上游区域少数荒地(包括农田荒地、河滩与农田边交错带及小路边);美洲商陆则多分布在林地边缘地带、村落附近与路边;曼陀罗多分布在农田荒地、林地与河滩地交错带挖沙残余沙石地及小路边。
4 结论与讨论
4.1 种类组成
伊洛河外来草本植物有27种,15科,以菊科、苋科、豆科等被子植物中的大科为主。分析其原因可能是因为:在全球占主导地位的植物更可能成功迁移到新环境中[22-23]。如,菊科是维管植物中种类最多的一个科,也是最为进化的科之一。其自身具有一些有利于扩散的特征,如高繁殖率、多样的繁殖方式、种子风媒传播等先进特殊机制[22]。苋科植物具有种子小、繁殖率高的特点,且全科大部分具有药用、食用和观赏价值,因此易被有意引入;豆科植物具有独特的固氮能力,通过营养的自我累积而获得资源上的优势。外来种向未分布地区迁移的过程中,有意或无意引入和引入后传播扩散是重要的两步。
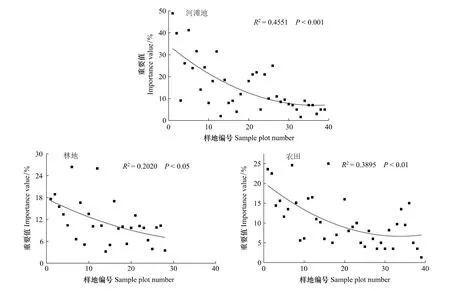
图3 外来草本植物沿伊洛河流域纵向梯度分布格局Fig.3 Variation of distribution patterns of alien herbs along the Yiluo River watershed
4.2 伊洛河外来草本植物分布格局
伊洛河流域地处半湿润、半湿润偏旱区域,位于黄河流域的中心位置,是我国原始农业发展最早地区之一,有着悠久的历史和灿烂的文明。随着当地农业、经济、旅游业的发展和人口的膨胀,其与外界的交流也日益频繁,一些外来种被引入并逐渐扩散开来。外来草本植物主要分布在资源丰富、受扰动大的地区。同时,经济发达、交通便利以及人类频繁的活动也极大地促进了物种在这些地区的扩散和传播[24]。外来种相对于本地种的生存状态可能取决于外界的环境胁迫[25]。一方面,人类活动导致土地利用方式的改变可能会增加外来种迁入并定居的机会[26-27],促进外来种的扩散和传播[28];另一方面,可获取资源的丰富程度与群落对入侵的易感性具有正相关性[29],群落有效性养分的增加可能会提高外来物种相对于本地种的适合度[30-31]。同时,养分富集和物理干扰也是人类活动影响下外来种入侵的重要特征。
流域横向梯度下各生境间外来种重要值差异并不显著,重要值平均值大体呈现河滩地>林地>农田的分布格局。Burke和Grime[32]根据生境资源的丰富程度以及人类活动干扰的强弱将各种生境类型归纳为:高养分高干扰(包括农业用地、房前屋后废弃地、水岸、沟渠边、公园等)、低养分高干扰(包括草坪、草场、人工林、天然林的间隙或边缘等)、高养分低干扰(包括森林等),未在以上几类的归为其他类型。农田生境类型中植物群落结构较为简单,稳定性较低,易受病虫草害的影响[33]。由于生物入侵主要由人为活动导致[34],因此,受人类影响最频繁和最直接的农田生境类型,也最容易遭受外来生物入侵。研究发现,农田生境类型中外来种出现个体数略低于另外两种生境类型。这一结果与Weber等[14]在对270种中国外来入侵植物研究时得出:受扰动最强但营养丰富的农田是中国入侵植物分布最多的生境的结论不同。这一原因可能是因为在干扰还没有达到物种最大耐受限度的情况下,外来种趋于分布在有干扰的环境中,但当干扰超出物种最大耐受限度时,外来种的分布种类数和个体数量则呈下降趋势。河滩地在水流带来的养分富集和季节性洪水物理干扰作用下,呈现出外来种个体数最多的分布格局。
本研究中,在伊洛河流域尺度下,沿着纵向环境梯度(海拔、水热条件以及人类活动等)上的变化,不同生境间外来草本植物重要值变化十分明显。总体上,各生境类型均呈现出:外来种在下游地区影响最为严重,上游影响较轻。形成这种格局的可能原因是:比如河滩地生境下,下游平原地区外来入侵种空心莲子草入侵严重,常沿河道大面积分布,故外来种在下游地区影响最为严重;而上游山区受地形等自然因素(如河道狭窄且密布鹅卵石等)及交通、经济等人类活动干扰较小,故外来种出现较少。林地生境类型下,外来种分布可能与林地的种植时间及种植密度有关,下游平原地区人工林地分布较少,且林龄较小,群落结构简单,但群落总盖度较高,外来种小蓬草、一年蓬分布较多;中上游地区人工林分布较多,且林龄相对较大,群落物种组成复杂,但群落总盖度较小,外来种小蓬草、一年蓬分布较少,间杂分布有红花酢浆草、圆叶牵牛、牵牛和紫苜蓿等。
伊洛河流域作为由山地、丘陵、平原等组成的相互依存、相互制约的等级自然体系,其上游山区受自然因素作用较大;下游平原地区作为人类社会经济活动中心,人为干扰强烈;而中游丘陵地区受自然因素和人为因素的双重影响景观生态系统更具有复杂性[13]。3种生境类型下,外来种均呈现在下游平原地区分布最多,上游山区分布最少的分布格局。这表明在受人类活动干扰较为强烈的地区外来种分布愈多,而在人为干扰较小地区外来种分布较少。林地中,随着人为干扰的减弱及草本层植被的自然演替,外来种呈逐渐减少趋势。流域内唯一恶性入侵种空心莲子草多出现在下游河滩地中且沿主河道成片分布,甚至在部分农田中也有发现。这可能与空心莲子草的水生和陆生的生活习性以及无性快速增殖方式有关。
不同外来种在各生境间分布差异明显。其中,小蓬草、钻叶紫菀和反枝苋广泛分布于3种类型中;稀见种有土荆芥、南苜蓿、白花草木樨、蓖麻、曼陀罗、野燕麦和节节麦等,只见于个别生境中且个体数量较少。所谓的稀见种并不是在流域中稀有,而是由于调查生境局限性造成的。如蓖麻,多分布在中上游区域少数荒地(包括农田荒地、河滩与农田边交错带及小路边);美洲商陆多分布在林地边缘地带、村落附近与路边;曼陀罗多分布在农田荒地、林地与河滩地交错带挖沙残余沙石地及小路边。虽然空心莲子草在整个流域中出现频次(出现样方数)相对较低,但在下游平原地区几乎出现在所有河滩及部分农田中,且危害较大,往往形成单优势种群,从而严重降低了当地的物种多样性。本研究仅选取了3种生境,未能涵盖所有类型。人类及其经济和非经济活动是外来种入侵的主要动因[35],裸地是外来杂草定居的主要生境,很多外来杂草,如一年蓬和小蓬草,均沿交通沿线分布[36]。调查中发现,一些人为干扰明显且地表裸露较大的道路旁、弃荒地、闲置地和暂时性裸地受外来种影响最大。
Ribeiro等在对西班牙Catalonia地区的研究中发现,土地的开发活动是影响区域尺度生物多样性变化的一个最主要因素,其中农业开发带来的威胁最大[37]。外来种成功入侵的机制可以简单地归结为外来种的入侵性特征、入侵地的生态系统特征以及人类活动等相互作用的结果,然而这又是一个悬而未决的问题[38]。分析认为,人类通过对土地开发利用等干扰活动直接改变了陆地生态系统的结构与组成,进而导致生物栖息地环境的改变是造成这种变化格局的直接原因。虽然,自然季节性洪水干扰在一定程度上干扰了群落结构的稳定型,为外来种侵入提供了机遇。但是,沿河道挖沙取石、改变土地利用方式、提取河水灌溉等强烈人为干扰使得原有物种自然分布格局被打破,导致了外来物种进入并扩散。
[1] Richardson D M,Pyšek P,Rejmánek M,Barbour M G,Panetta F D,West C J.Naturalization and invasion of alien plants:concepts and definitions.Diversity and Distributions,2000,6(2):93-107.
[2] Pimentel D,McNair S,Janecka J,Wightman J,Simmonds C,O'Connell C,Wong E,Russel L,Zern J,Aquino T,Tsomondo T.Economic and environmental threats of alien plant,animal,and microbe invasions.Agriculture,Ecosystems and Environment,2001,84(1):1-20.
[3] Perrings C,Dehnen-Schmutz K,Touza J,Williamson M.How to manage biological invasions under globalization.Trends in Ecology and Evolution,2005,20(5):212-215.
[4] Pimentel D,Zuniga R,Morrison D.Update on the environmental and economic costs associated with alien-invasive species in the United States.Ecological Economics,2005,52(3):273-288.
[5] Rudgers J A,Orr S.Non-native grass alters growth of native tree species via leaf and soil microbes.Journal of Ecology,2009,97(2):247-255.
[6] Khuroo A A,Dar G H,Khan Z S,Malik A H.Exploring an inherent interface between taxonomy and biodiversity:current problems and future challenges.Journal for Nature Conservation,2007,15(4):256-261.
[7] Lake JC,Leishman MR.Invasion success of exotic plants in natural ecosystems:the role of disturbance,plant attributes and freedom from herbivores.Biological Conservation,2004,117(2):215-226.
[8] Yang B,Zhuoga Y J,Pan X Y,Xu H G,Li B.Alien terrestrial herbs in China:diversity and ecological insights.Biodiversity Science,2010,18(6):660-666.
[9] DAISIE.Handbook of alien species in Europe.Dordrecht,Netherlands:Springer,2009.
[10] Mack R N,Simberloff D,Mark Lonsdale W,Evans H,Clout M,Bazzaz F A.Biotic invasions:causes,epidemiology,global consequences,and control.Ecological Applications,2000,10(5):689-710.
[11] Rozefelds A C,Cave F L,Morris D I,Buchanan A M.The weed invasion in Tasmania since 1970.Australian Journal of Botany,1999,47(1):23-48.
[12] Pyšek P,Richardson D M,Rejmánek M,Webster G,Williamson M,Kirschner J,Pysek P,Rejmanek M.Alien plants in checklists and floras:towards better communication between taxonomists and ecologists.Taxon,2004,53(1):131-143.
[13] Chen J,Guo Y L,Lu X L,Ding S Y,Su S,Guo J J,Li Q X.Species diversity of herbaceous communities in the Yiluo River basin.Acta Ecologica Sinica,2012,32(10):3021-3030.
[14] Weber E,Li B.Plant invasions in China:what is to be expected in the wake of economic development?BioScience,2008,58(5):437-444.
[15] Wan F H,Guo J Y,Zhang F.Research on Biological Invasions in China.Beijing:Science Press,2009.
[16] Guo Y L,Lu X L,Ding SY.The classification of plant functional types based on the dominant herbaceous species in the riparian zone ecosystems in the Yiluo River.Acta Ecologica Sinica,32(14):4434-4442.
[17] Ma Z Y.The analysis of hydrology characteristic in Yiluo River.Hydrology,1998,2:57-58.
[18] Feng D K.Analysis of the modern geomorphological prosses in the Yiluo River basin.Henan Science,1992,10(3):308-312.
[19] Ding SY,Qian L X,Cao X X,Li S,Li H M.Forest landscape patterns dynamicsof Yihe-Louhe River basin.Acta Geographica Sinica,2003,58(3):354-362.
[20] Ding SY,Liang G F.Geographic environmental factors on forest landscape dynamics of Yiluo River basin.Geographical Research,2007,26(5):906-915.
[21] Guo Y L,Su S,Ding SY,Liang GF.Species diversity of riparian herbaceous community in the upper Luo River riparian zones.Journal of Henan University:Natural Science,2011,41(1):67-71.
[22] Heywood V H.Patterns,extents and modes of invasion by terrestrial plants//Drake JA.Biological Invasions:A Global Perspective.Chichester:Wiley,1989.
[23] Pyšek P,Richardson D M,Pergl J,Jarosik V,Sixtova Z,Weber E.Geographical and taxonomic biases in invasion ecology.Trends in Ecology and Evolution,2008,23(5):237-244.
[24] Ding J Q,Mack R N,Lu P,Ren M X,Huang H W.China's booming economy is sparking and accelerating biological invasions.BioScience,2008,58(4):317-324.
[25] Alpert P,Maron J L.Carbon addition as a countermeasure against biological invasion by plants.Biological Invasions,2000,2(1):33-40.
[26] Lonsdale W M,Lane A M.Tourist vehicles as vectors of weed seeds in Kakadu Nationl Park,Northern Australia.Biological Conservation,1994,69(3):277-283.
[27] Smith M D,Knapp A K.Alien plant species in a C4-dominated grassland:invisibility,disturbance and community structure.Oecologia,1999,120(4):605-612.
[28] Hellmann J J,Byers J E,Bierwagen B G,Dukes J S.Five potential consequences of climate change for invasive species.Conservation Biology,2008,22(3):534-543.
[29] Davis M A,Grime J P,Thompson K.Fluctuating resources in plant communities:a general theory of invasibility.Journal of Ecology,2000,88(3):528-534.
[30] Green E K,Galatowitsch SM.Effects of Phalarisarundinacea and nitrate-N addition on the establishment of wetland plant communities.Journal of Applied Ecology,2002,39(1):134-144.
[31] Minchinton T E,Bertness M D.Disturbance-mediated competition and the spread of Phragmites australis in a coastal marsh.Ecological Applications,2003,13(5):1400-1416.
[32] Burke M J W,Grime J P.An experimental study of plant community invasibility.Ecology,1996,77(3):776-790.
[33] Qiang S,Chen G Q,Li B P,Meng L.Invasive alien species in Chinese agricultural ecosystems and their management.Biodiversity Science,2010,18(6):647-659.
[34] Winter M,Kuhn I,La Sorte F A,Schweiger O,Nentwig W,Klotz S.The role of non-native plants and vertebrates in defining patterns of compositional dissimilarity within and across continents.Global Ecology and Biogeography,2010,19(3):332-342.
[35] Mack R N,Lonsdale W M.Humans as global plant dispersers:getting more than we bargained for.Bioscience,2001,51(2):95-102.
[36] Auld BA,Martin PM.The autoecology of Eupatorium adenophorun Spreng in Australia.Weed Research,1975,15(1):27-31
[37] Ribeiro R,Santos X,Sillero N,Carretero M A,Llorente G A.Biodiversity and Land uses at a regional scale:Is agriculture the biggest threat for rep tile assemblages?Acta Oecologica,2009,35(2):327-334.
[38] Li B,Ma K P.Biological invasions:opportunities and challenges facing Chinese ecologists in the era of translational ecology.Biodiversity Science,2010,18(6):529-532.
参考文献:
[8] 杨博,央金卓嘎,潘晓云,徐海根,李博.中国外来陆生草本植物:多样性和生态学特性.生物多样性,2010,18(6):660-666.
[13] 陈杰,郭屹立,卢训令,丁圣彦,苏思,郭静静,李乾玺.伊洛河流域草本植物群落物种多样性研究.生态学报,2012,32(10):3021-3030.
[16] 郭屹立,卢训令,丁圣彦.伊洛河河岸带生态系统草本植物功能群划分.生态学报,2012,32(14):4434-4442.
[17] 马志有.伊洛河水系水文特性浅析.水文,1998,2:57-58
[18] 冯大奎.伊洛河流域现代地貌过程分析.河南科学,1992,10(3):308-312
[19] 丁圣彦,钱乐祥,曹新向,李爽,李昊民.伊洛河流域典型地区森林景观格局动态.地理学报,2003,58(3):354-362.
[20] 丁圣彦,梁国付.地理环境因素对伊洛河流域森林景观的影响.地理研究,2007,26(5):906-915.
[21] 郭屹立,苏思,丁圣彦,梁国付.洛河上游河漫滩草本植物群落物种多样性研究.河南大学学报:自然科学版,2011,41(1):67-71.
[33] 强胜,陈国奇,李保平,孟玲.中国农业生态系统外来种入侵及其管理现状.生物多样性,2010,18(6):647-659.
[38] 李博,马克平.生物入侵:中国学者面临的转化生态学机遇与挑战.生物多样性,2010,18(6):529-532.
