乳山近海大型底栖动物功能摄食类群
2013-09-07彭松耀李新正
彭松耀,李新正
(1.中国科学院海洋研究所,青岛 266071;2.中国科学院研究生院,北京 100049)
近年来沿海地区经济高速发展,导致近海水质污染、生态环境恶化等一系列生态功能衰退问题的出现,近海生态系统是世界上受到最大威胁的生态系统之一,因此近海生态系统健康及其相关研究也受到广泛重视。
功能摄食类群(FFGs)最初由Cummins[1]在20世纪70年代提出,海洋大型底栖动物功能摄食类群经Pearson等学者的努力[2-5],其分类系统已基本完善,主要分为6个功能摄食类群:植食者、食底泥者、滤食者、肉食者食碎屑者和杂食者。
组成功能群落的不同物种,主要是通过食物关系而结合起来的[6],因此,功能摄食类群对研究生物群落结构和功能具有重要作用。大型底栖动物是近海生态系统食物链中有机物、营养源与更高营养级生物的主要连接体,同时大型底栖动物对外界胁迫较为敏感,群落结构与功能属性均能从不同侧面反映水质状况的好坏,从而有效地指示水生生态系统的健康[7]。目前,国际上有关大型底栖动物群落功能研究,主要集中于对底栖营养类群的研究[8-12],研究方法大多运用多元统计分析等。目前,我国有关海洋大型底栖动物功能摄食类群的研究有所报道[13-14],但包括乳山近海在内的山东半岛及黄海海域却鲜有报道,仅见乳山近海大型底栖动物群落结构分析以及与环境因子关系的研究[15]。鉴于此,本研究以乳山近海大型底栖动物为对象,对大型底栖动物功能摄食类群的时空分布规律以及与环境因子的相互关系进行研究,并尝试应用基于功能摄食类群的多项参数对乳山近海生态系统进行评价,以期为进一步的生态系统变化及其与环境影响因子的相互作用研究提供基础资料。
1 材料与方法
1.1 调查区域
乳山位于东经121°31',北纬36°54',地处山东半岛东北部,濒临黄海。年平均气温在12.3℃,是典型的暖温带海洋性气候。本研究在乳山近海设置26个采样站(图1),于2006年12月和2007年5月、8月、11月对大型底栖动物进行调查采样。
1.2 底栖动物的采集及鉴定
每个采样站用0.1m2Gray-O'Hara箱式采泥器重复采样2次,用底层孔径为0.5mm的套筛筛选,两次取样获得的样品合并作为该站该次采样0.2m2内样品,所获样品用75%的酒精固定后带回实验室,随后进行种类鉴定、个体计数、称重(使用0.001g精度天平),并对所获数据进行统计分析。
1.3 理化指标的测定及方法
水温(Temp)、水深、盐度、pH值用AAQ1183-IF型多功能温盐深仪(CTD)现场同步测定。于每个样点采集2瓶600 mL底层水样,其中一瓶现场加硫酸调整pH值<2,低温保存。带回实验室后测定其化学指标,包括总氮(TN)、总磷(TP)、硅酸盐(SiO3-Si)、总有机碳(TOC)、化学需氧(COD)、溶氧(DO)、悬浮体(SS)和叶绿素 a(Chl-a)共计8项指标[16]。
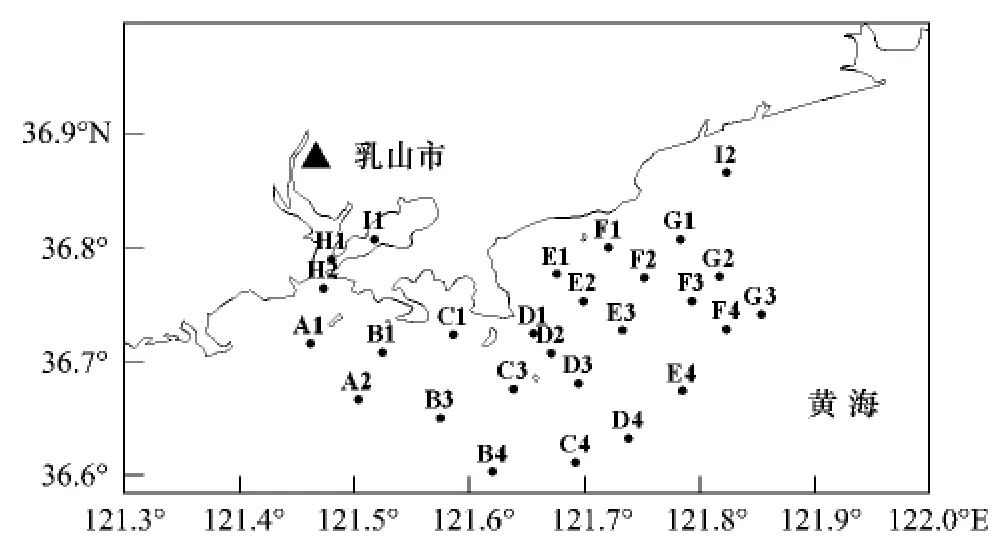
图1 乳山近海站位图Fig.1 Sampling stations of macrobenthos from the coastal water off Rushan,Shandong Peninsula
1.4 大型底栖动物功能群的划分
本文主要根据海洋大型底栖动物的食性类型划分功能群[2-6],将大型底栖动物分为以下6类功能群:(1)植食者(H),主要以某些藻类和底栖硅藻为食。(2)滤食者(FF),滤食水体中的有机质和微小浮游生物。(3)食底泥者(DF),以沉积物里的有机碎屑、细菌和底栖藻类为食。(4)食碎屑者(DTF),以沉积物表面的有机碎屑、细菌和底栖藻类为食。(5)肉食者(C),捕食者和食腐动物。(6)杂食者(O),依靠皮肤或鳃的表皮,直接吸收溶解在水中的有机物,也可以取食植物腐叶和小型双壳类、甲壳类。
1.5 大型底栖动物生态类群参数
根据Gamito和Furtado的研究[17],本研究主要从大型底栖动物功能类群的多样性以及对环境的响应等方面入手,应用基于大型底栖动物功能类群的参数对近海生态系统进行评价(表1,表2)。运用Shannon-Wiener指数(H'FD)、Pielou指数(J'FD)、底内动物指数(ITI)和海洋生物指数(AMBI)对近海生态系统进行评价。计算公式和相关参数见(表1,表2)。

表1 H'FD指数和J'FD指数的计算公式及相关参数[18]Table1 Formula and parameters for feeding diversity measured by Shannon-Wiener information index or by evenness when six feeding groups were considered
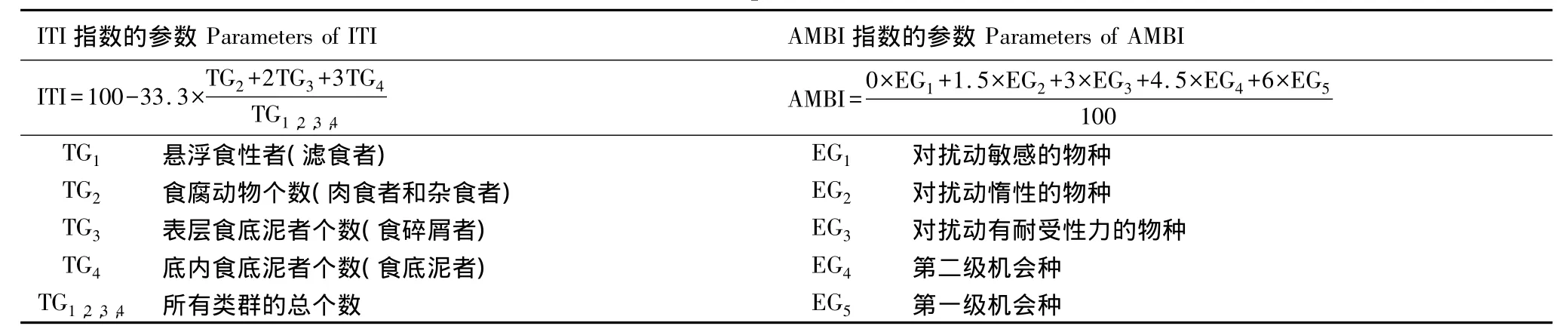
表2 ITI指数(Infaunal Trophic Index)和AMBI指数的计算公式及相关参数[19-21]Table2 Formula and parameters for ITI and AMBI
1.6 数据分析处理
大型底栖动物功能摄食类群的多样性分析采用PRIMER 5软件,AMBI指数计算采用软件AMBI 4.0(http://www.ambi.azti.es),冗余分析(RDA)采用软件Canoco 4.5。
2 结果
2.1 功能摄食类群的组成
对乳山近海大型底栖动物功能摄食类群进行分析,结果表明,食底泥者和肉食者在乳山近海占较大优势,相对丰度分别为34%和31%,其次为滤食者,相对丰度为22%、食碎屑者、杂食者、植食者相对丰度较小,分别为7%、5%、1%(图2)。
食底泥者、肉食者和滤食者的优势物种见物种排序丰度图(图3),由图可知,中蚓虫Mediomastus californiensis、不倒翁虫Sternaspis scutata、丝异蚓虫Heteromastus filiforms和轮双眼钩虾Ampelisca cyclops为食底泥者类群的优势种;寡鳃齿吻沙蚕 Nephtys oligobranchia、圆筒原盒螺Eocylichna cylindrella和纽虫Nemertinea sp.为肉食者类群的优势种;滑理蛤 Theora lubrica、菲律宾蛤仔Venerupis philippinarum、金星蝶铰蛤 Trigonothracia jinxingae和江户明樱蛤Moerella jedoensis为滤食者类群的优势种。

图2 乳山近海大型底栖动物各功能摄食类群相对丰度Fig.2 The abundant ratios of the FFGs in the total macrobenthic abundance from the coastal water off Rushan,Shandong Peninsula
2.2 功能摄食类群的时空分布
2.2.1 相对丰度和平均丰度的时间分布
周年乳山近海大型底栖动物功能摄食类群相对丰度和平均丰度见图4。食底泥者、肉食者和滤食者相对丰度和平均丰度明显高于其它功能摄食类群,优势地位明显,食底泥者相对丰度最大出现在2006年12月,为39.3%,平均丰度最大则出现在2007年5月,为107.4个/m2;肉食者和滤食者相对丰度均出现在2007年11月,分别为37.9%和27.2%,平均丰度最大均出现在2007年5月,分别为94.7个/m2和77.4个/m2;植食者相对丰度和平均丰度则较低,最大值均出现在2006年12月,分别为1.1%和2.9个/m2。
2.2.2 相对丰度的空间分布
周年乳山近海大型底栖动物功能摄食类群相对丰度空间分布见图5。由图可知,优势类群食底泥者、相对丰度最大出现在2007年8月G1站,为73.3%,最小出现在2007年8月H1和H2站,为0;肉食者相对丰度最大出现在11月A1站,为74%,最小出现在2007年8月H1站,为0;滤食者相对丰度最大分别出现在8月H1站,为100%;最小分别出现在2006年12月H2站、2007年8月E2和F1站和2007年11月A2和C3站,均为0。食碎屑者、杂食者和植食者相对丰度最大分别出现在2007年8月G3站、5月的D1站和2006年12月A1站,分别为30.6%、27.6%和5.3%。
2.2.3 丰度的空间分布
周年乳山近海大型底栖动物功能摄食类群丰度空间分布见图6,由图可知,优势类群食底泥者最大出现在2007年5月B4站,为385个/m2,最小出现在2007年8月H1和H2站,为0;肉食者最大出现在2007年11月A1站,为285个/m2和,最小出现在2007年8月H1站,为0;滤食者丰度最大出现在2007年5月D2站,丰度为325个/m2,最小出现在2006年12月H2站、2007年8月E2和F1站、11月A2和C3站,均为0。食碎屑者、杂食者和植食者丰度最大分别出现在2006年12月G2站、2007年5月B4站和2006年12月D3站,丰度分别为 195、135、20 个/m2。
2.3 应用基于功能类群的参数对乳山近海生态系统进行评价
2.3.1 各项参数的时间变化
对功能摄食类群各项参数的时间动态进行分析(图7),结果表明:乳山近海J'FD、H'FD、ITI和AMBI指数平均值最大分别为0.83(5月)、1.81(12月)、54(11月)和1.95(5月)。J'FD、H'FD、ITI和AMBI指数最小为0.8(8月)、1.65(8月)、44(12月)和1.33(11月)。
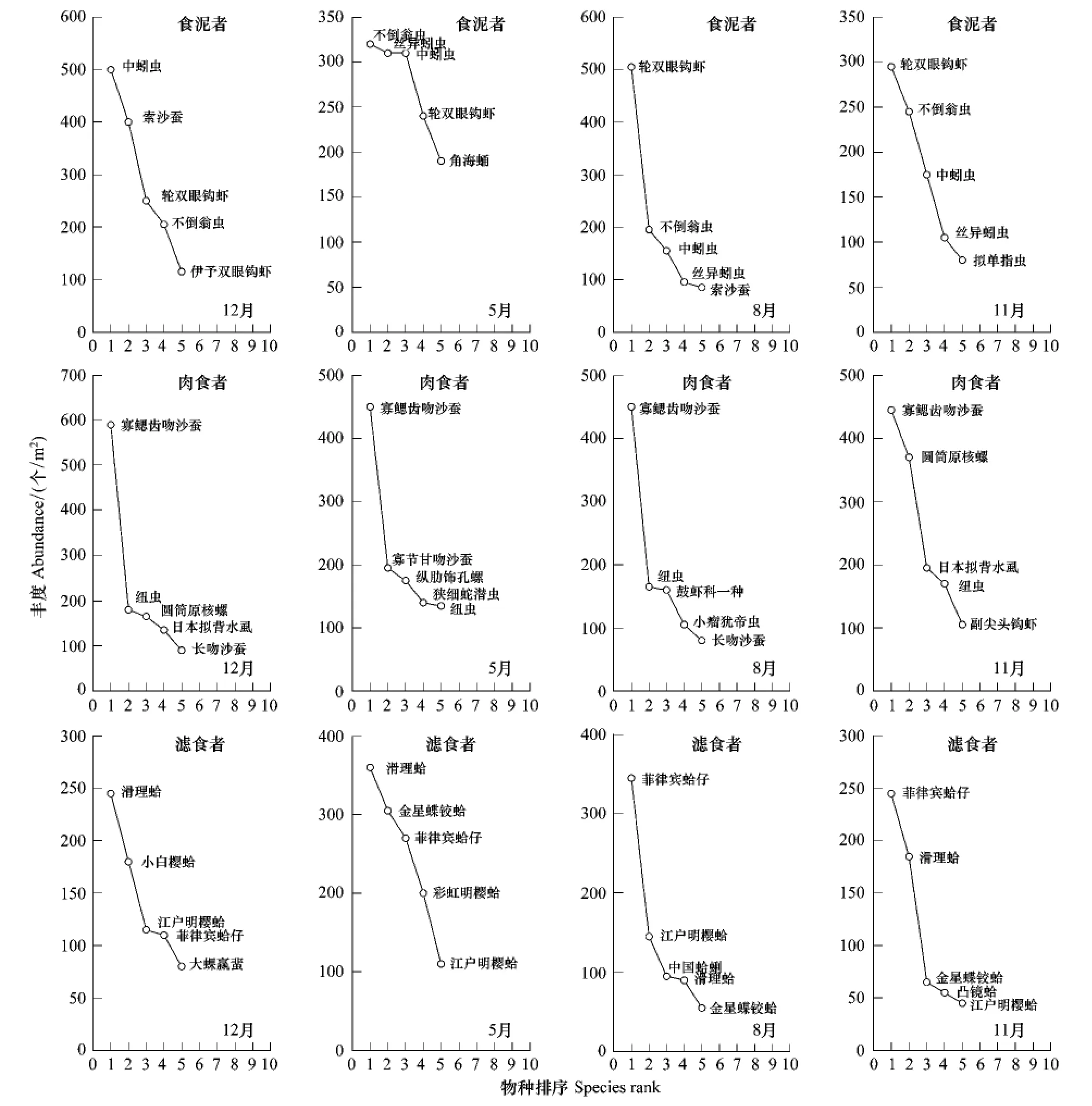
图3 乳山近海大型底栖动物食底泥者、肉食者和滤食者类群的优势种Fig.3 The dominant species of the three FFGs of macrobenthos in the coastal water off Rushan,Shandong Peninsula in different seasons
2.3.2 各项参数的空间变化
对各项参数的空间动态进行分析(图7),结果表明:J'FD、H'FD、ITI和AMBI指数最大分别出现在C3站(11月)、E4站(5月)、H1站(8月)和I2站(5月),指数值分别为1、2.15、100和4.37。J'FD指数最小出现在G1站(12月)和A1站(11月),均为0.56;H'FD指数最小出现H1站(8月),为0;ITI和AMBI指数最小均出现在G1站(8月),分别为19和0.5。
根据J'FD、ITI和AMBI指数空间分布结果和指数分级状况(图7和表3),发现A1站(11月)、G1站(12月)和I2站(8月)J'FD指数值均低于0.6;G1站(除5月)和I2站(8月)ITI指数值均低于30;I2站(5月)AMBI指数值大于4.3。
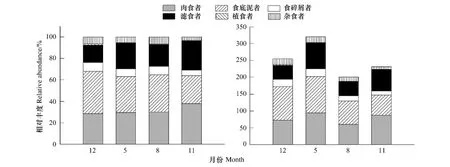
图4 大型底栖动物各功能摄食类群相对丰度和平均丰度的季节动态Fig.4 The abundant ratios of macrozoobenthic functional feeding groups in total abundance in different seasons
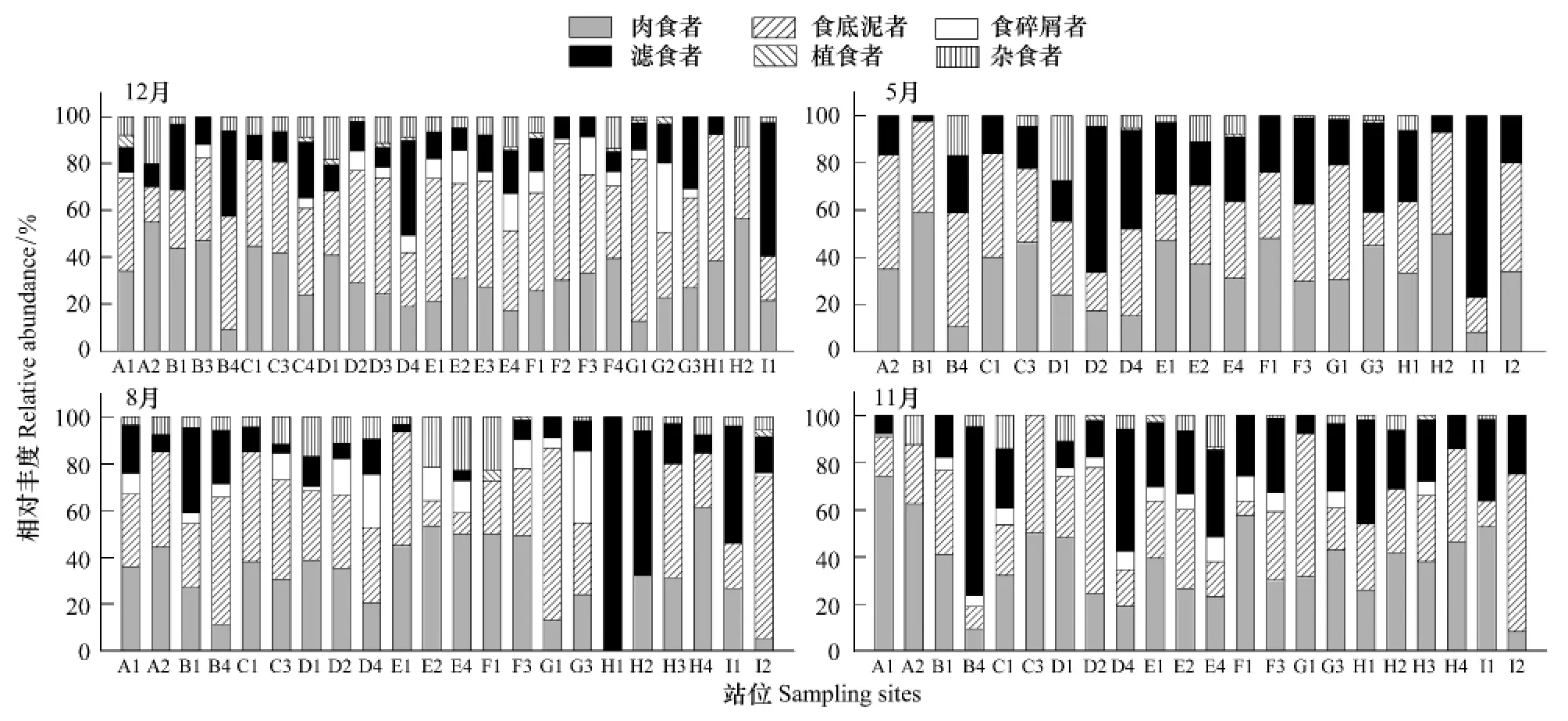
图5 各季节不同站位大型底栖动物各功能摄食类群相对丰度Fig.5 The abundant ratios of the six feeding groups in total abundance of different stations in different seasons
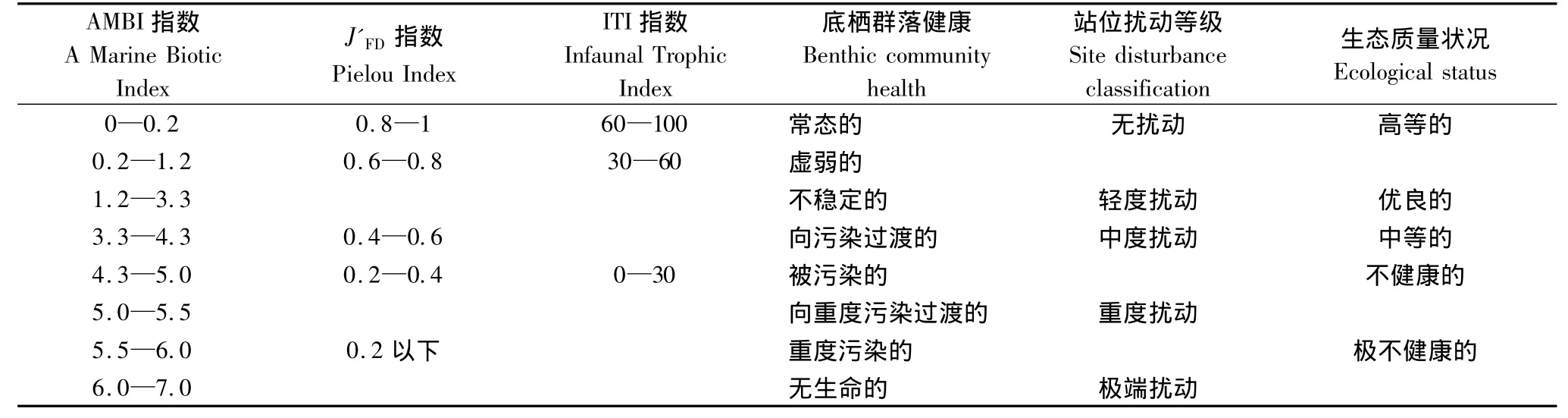
表3 AMBI指数、J'FD 指数和 ITI指数(Infaunal Trophic Index)的分级[17,22-25]Table3 Classifications of AMBI,J'FD,ITI
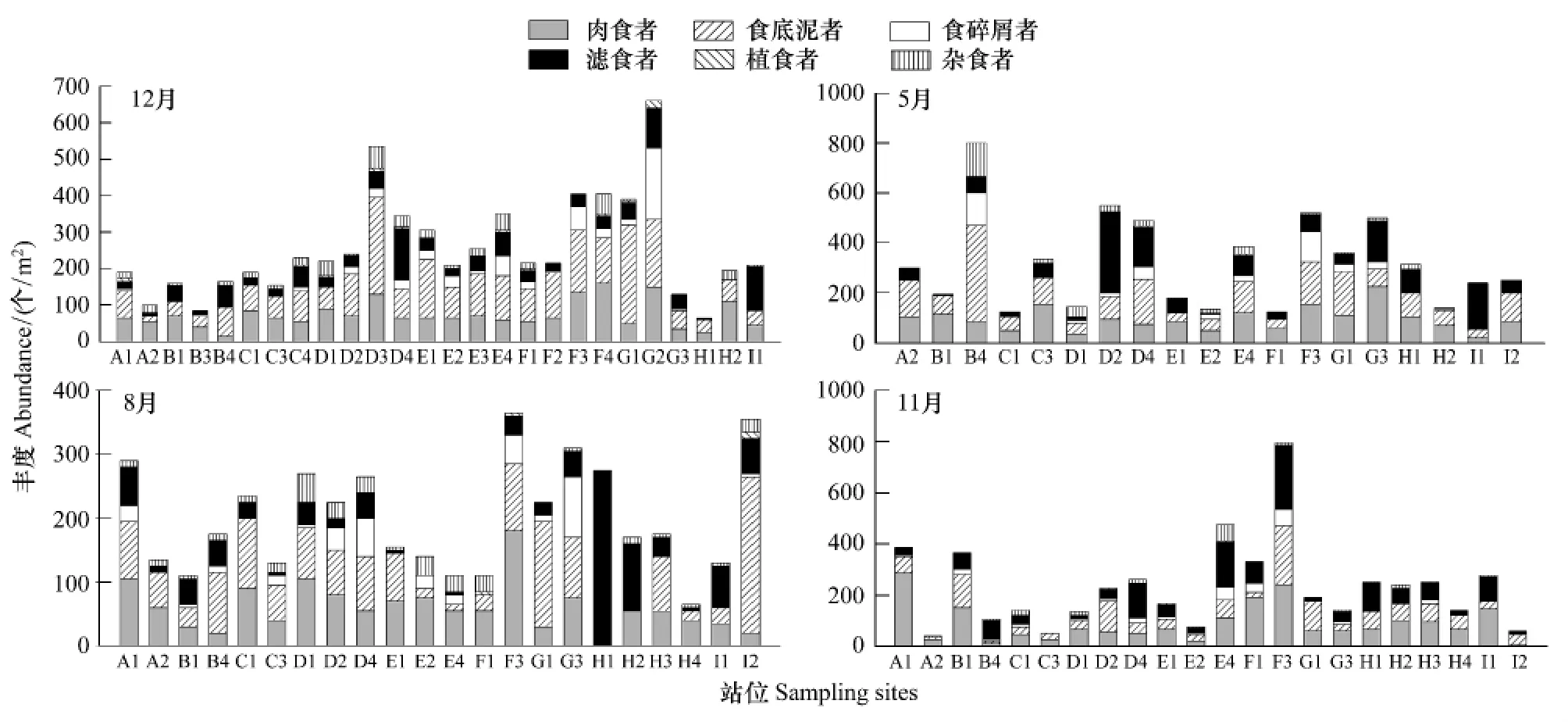
图6 各季节不同站位大型底栖动物各功能摄食类群丰度Fig.6 Abundances of the six feeding groups and total abundance of macrobenthos in different stations in different seasons
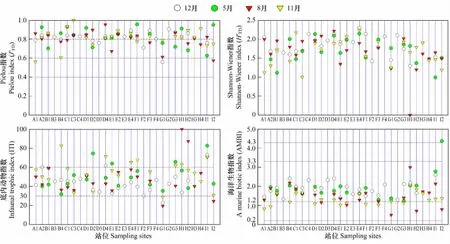
图7 研究区大型底栖动物Pielou指数、Shannon-Wiener指数、底内动物指数和海洋生物指数Fig.7 Values of J'FD,H'FD,ITI,AMBI in different stations
综合上述分析,对乳山近海底栖生态环境质量进行评价。结果表明:乳山近海底栖群落健康状况由虚弱趋于不稳定,生态质量状况较为优良,大部分站位底栖生物群落受到轻度扰动。而周年A1站、G1站和I2站生态参数值均较低,表明上述站位底栖生态环境质量较低,底栖动物群落受到中度扰动。评价结果与李新正等[15]的研究较为一致。
2.4 乳山近海大型底栖动物功能摄食类群与环境因子之间的冗余分析
为了解调查期间不同环境因子对研究海域大型底栖动物功能摄食类群的影响状况,将大型底栖动物摄食功能类群同水温、水深、盐度、pH值、总氮、总磷、硅酸盐、总有机碳、化学需氧、溶氧、悬浮体和叶绿素a等12个环境因子进行冗余分析(图8)。
通过蒙特卡洛检验(Monte Carlo Permutation Test),排除贡献小的因子(P<0.05),发现2007年5月盐度、水温、总氮和悬浮物等4个因子与乳山近海大型底栖动物功能摄食类群存在较显著的相关性;2007年8月溶解氧和水温与大型底栖动物功能摄食类群存在较显著的相关性;2007年11月和2006年12月则分别是总有机碳和硅酸盐与乳山近海大型底栖动物功能摄食类群存在较为显著的相关性。
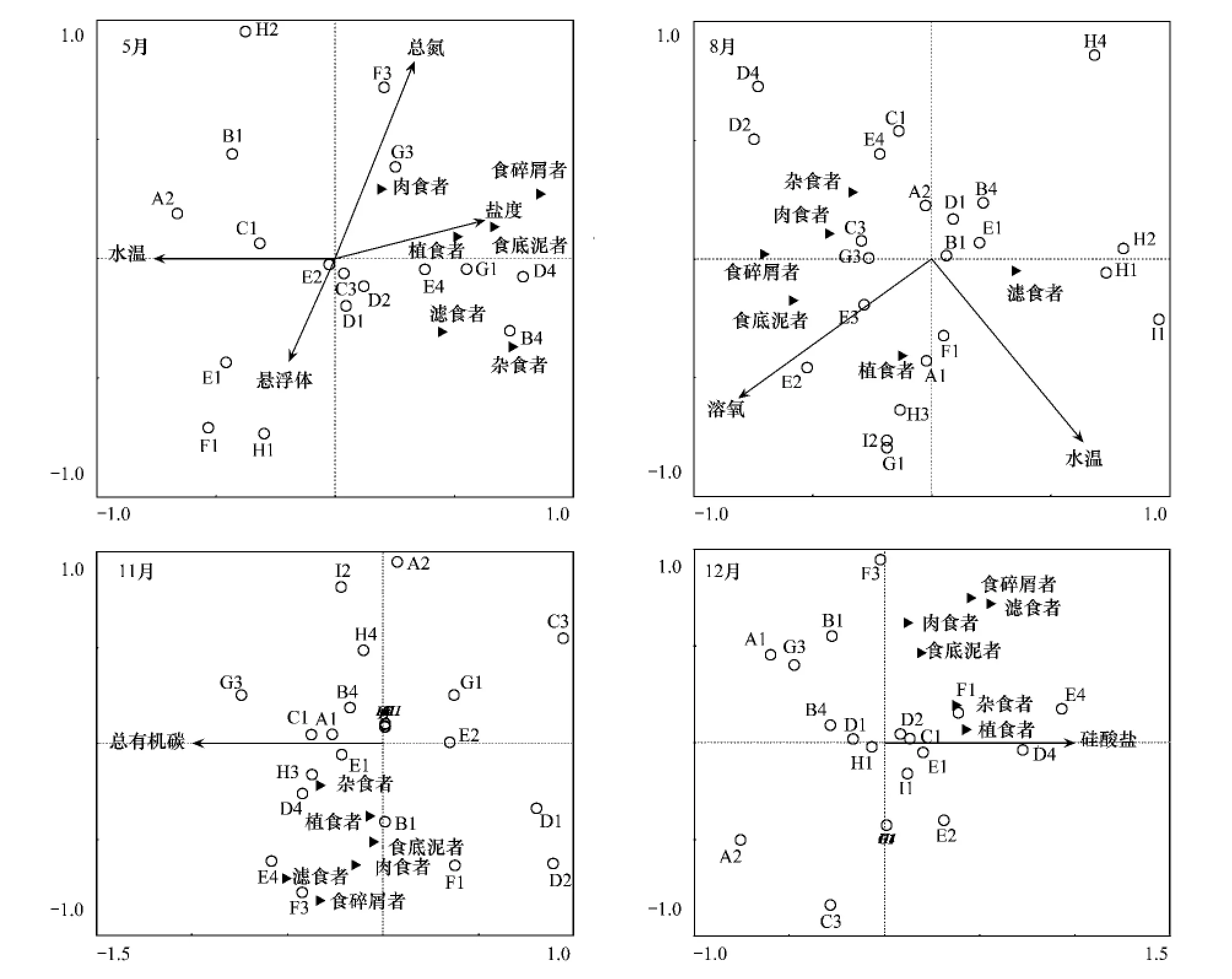
图8 乳山近海各季度大型底栖动物与环境变量之间的冗余分析Fig.8 Relationships of macrozoobenthos and environmental factors obtained by Redundancy analysis(RDA)from off Rushan,Shandong Peninsula in different seasons
3 讨论
3.1 功能摄食类群的组成
乳山近海大型底栖动物食底泥者、肉食者和滤食者功能摄食类群相对丰度较高,占优势地位,结果与葡萄牙Tagus河口[26]及西班牙西北部维哥湾[27]报道较一致。对比国内其他研究区域,乳山近海底栖动物物种数、优势功能类群和种类组成存在较大差别,主要表现为:乳山近海底栖动物物种数(236种),远高于象山港[28](95 种)、胶州湾潮滩[29](71 种)、长江口南岸潮滩[14](55 种)和灵昆岛潮滩[13](33 种)等区域。乳山近海大型底栖动物以食底泥者为优势功能类群,而灵昆岛潮滩和长江口九段沙等区域优势功能类群分别为肉食者、植食者和滤食者;与国内一些研究区域功能群代表种比较,乳山近海食底泥者代表种为中蚓虫、不倒翁虫、丝异蚓虫和轮双眼钩虾,肉食者代表种为寡鳃齿吻沙蚕、圆筒原盒螺和纽虫,滤食者代表种为滑理蛤、菲律宾蛤仔、金星蝶铰蛤和江户明樱蛤,而长江口九段沙为麂眼螺Rissoina sp.和河蚬Corbicula fluminea,灵昆岛潮滩为日本沙蚕Neanthes japonica、彩虹明樱蛤 Moerella iridescen和焦河蓝蛤 Potamocorbula ustulata,象山港为缢蛏Sinonovacula constricta、日本刺沙蚕Neanthes japonica和纵肋织纹螺Nassarius variciferus。对比发现,不同研究区,功能群的代表种不同,其主要原因可能是各研究区地理尺度不同,生境状况各异,而底栖功能群的物种组成和分布是各个物种对栖息地不同环境因子的综合反应,如葛宝明等研究显示了功能群的生境梯度变化[13],袁兴中等也发现长江口河口上游和污染严重断面,功能群单一,优势种为食底泥者霍普水丝蚓Limnodrilus hoffmeisteri和小头虫Capitella capitata,下游浏河口优势种为河蚬,属淡水种,与该处的沉积物盐度较低有关[14]。反映了底栖动物功能群物种组成和多样性与环境梯度和生境质量有着必然联系。
3.2 乳山近海大型底栖动物功能摄食类群的时空分布
底栖动物功能群丰度的季节变化与种类生殖习性、种间关系和人类干扰有关[30-32]。乳山近海5月各类群丰度最高,这可能与春季许多幼体补充进了底栖生物的群落有关,有研究发现软体动物繁殖季节多在5—6月,许多新个体进入群落,丰度达最高值[30]。8月各类群丰度为周年最低,这可能与底栖动物的自然死亡、高营养级捕食者对底栖动物的摄食和渔业捕捞有一定的关系[33]。例如,胶州湾夏季至冬季随着软体动物幼体自然死亡和被捕食,因此栖息丰度也随着降低。此外,对乳山近海拖网出现的鹰爪虾 Trachysalambria curvirostris而言,多毛类丝异蚓虫是其主要的摄食对象[34]。
沉积物类型是底栖生物空间异质性的主要影响因素[26],食底泥者在软底质中数量较多,滤食者则多分布于较粗底质中[35-36]。乳山近海B4站(5月),食底泥者丰度最大,为385个/m2,采样底质为软泥,角海蛹、丝异蚓虫和不倒翁虫对丰度贡献较大,这可能与底质有机质含量较高有关。角海蛹、丝异蚓虫和不倒翁虫这些物种多分布在底质颗粒较细、有机质较为丰富的海域,并且对有机质富集环境有较强耐受性[37-38];H1站(8月)未采集到食底泥者,而5月和11月食底泥者丰度均在70个/m2以上。H1站食底泥者丰度季节的差异可能与采样的随机性、沉积物及底栖生物的斑块分布有关。D2站(5月)滤食者丰度最大,为325个/m2,埋栖性的金星蝶铰蛤、滑理蛤和江户明樱蛤数量较多,这可能与该站底层水悬浮体浓度(22.6 mg/L)较高有关,Vale等、Boaventura研究发现滤食者多分布在有机悬浮物颗粒浓度较高的水域[39-40]。此外有研究发现浮游植物丰度与滤食者分布密切相关[41],如Saito等发现滑理蛤种群是 Maizuru湾浮游植物生产量主要消费者[42],Horikoshi等也报道了滑理蛤分布区藻华发生频率较高[43]。肉食者的丰度最大出现在A1站(11月),丰度为285个/m2,寡鳃齿吻沙蚕为该站优势种,可能与A1站(11月)底质有机质丰富有关,蔡立哲等[44]就发现寡鳃齿吻沙蚕的数量与有机质含量有正相关关系,此外Wijsman等人[45]认为肉食者高丰度区碳通量可能较高,支撑较高营养级生物生存。
3.3 乳山近海大型底栖动物功能摄食类群与环境之间的关系
冗余分析表明,调查期间底层水的盐度、总氮、水温、悬浮物浓度、有机质浓度、溶解氧和硅酸盐浓度是影响乳山近海功能摄食类群的环境因子。5月乳山近海水体盐度与各功能类群相关性较强,结果与Nanami等[46]的研究结果一致,其原因是底栖动物的分布受盐度影响[47]。5月和8月水温与乳山近海功能群丰度相关性较强,与徐勤增等对夏季南黄海大型底栖动物分布受水温影响较一致[48],也类似于Dolbeth等的研究结果[10],底栖生物生长发育和繁殖及分布范围受水温影响[27]。5月总氮与肉食者的丰度有较强相关性,可能与沉积物受扰动、生源要素氮释放和动物残体的分解有关,如航道疏浚扰动使食腐动物丰度增高[49]。此外,水体悬浮体浓度与滤食者相关性较强,结果类似于Muniz和Pires的研究[50],水体悬浮颗粒物浓度较高为滤食者提供了丰富的饵料。溶解氧是影响底栖生物分布的主要因素[3,50-51],8月乳山近海底层水溶解氧与功能群分布相关性较强,夏季底栖生物新陈代谢加强,摄食和呼吸会消耗大量的溶解氧,水体耗氧有机质分解也抑制底栖生物,进而影响底栖生物群落结构和分布。除了盐度、水体悬浮物浓度和溶解氧,沉积物有机质含量也是影响底栖生物分布的重要因素[50,52-55]。11月乳山近海底层水的有机质浓度与各功能群相关性较强,这可能与调查水域有机负荷较高及水层-底栖界面耦合作用有关。12月硅酸盐与乳山近海功能群相关性较强,尤其与植食者关系密切,这可能与植食者的饵料较为丰富有关,因为硅酸盐浓度较高区域藻类生长良好[56],此外,Bartoli等在研究水层系统与底栖系统物质循环时发现硅酸盐与菲律宾蛤仔的生物量相关性较强[57]。
总之,不同调查海域与调查时间影响大型底栖动物营养结构的环境因素不同,要深入了解非生物或生物因素对大型底栖动物的影响,需要全面了解大型底栖动物群落的功能。
致谢:本课题组王洪法、张宝琳、李宝泉、王金宝、周进、韩庆喜、董超、董栋、马林、蔡文倩、孙德海以及中国科学院海洋生物标本馆王少青、帅莲梅等参加了野外调查和实验室样品分析工作,在此一并致以衷心的感谢。
[1] Cummins K W.Structure and function of stream ecosystems.BioScience,1974,24(11):631-641.
[2] Pearson T H.Studies on the ecology of the macrobenthic fauna of Lochs Linnhe and Eil,west coast of Scotland.Ⅱ.Analysis of the macrobenthic fauna by comparison of feeding groups.Vie et Milieu,1971,1:53-91.
[3] Gaston G R.Benthic polychaeta of the Middle Atlantic Bight:feeding and distribution.Marine Ecology-Progress Series,1987,36:251-262.
[4] Sanchez-Mata A,M Lastra,JMora.Macrobenthic crustacean characterization of an estuarine area.Crustaceana,1993,64(3):337-355.
[5] Fauchald K,Jumars P A.The diet of worms:a study of polychaete feeding guilds.Oceanography and Marine Biology:An Annual Review,1979,17:193-284.
[6] He Z H.Freshwater Ecology.Beijing:China Agriculture Press,2000:101-102.
[7] Ma T W,Huang Q H,Wang H,Wang Z J,Wang C X,Huang SB.Selection of benthic macroinvertebrate-based multimetrics and preliminary establishment of biocriteria for the bioassessment of the water quality of Taihu Lake.Acta Ecologica Sinica,2008,28(3):1192-1200.
[8] Wijnhoven S and Hummel H.Patterns in macrozoobenthic assemblages indicate the state of the environment:insights from the Rhine-Meuse estuary.Marine Ecology Progress Series,2011,436:29-50.
[9] Lavesque N,Blanchet H,de Montaudouin X.Development of a multimetric approach to assess perturbation of benthic macrofauna in Zostera noltii beds.Journal Experimental Marine Biology and Ecology,2009,368(2):101-112.
[10] Dolbeth M,Teixeira H,Marques J C,Pardal M A.Feeding guild composition of a macrobenthic subtidal community along a depth gradient.Scientia Marina,2009,72(2):225-237.
[11] Macdonald T A,Burd B J,van Roodselaar A.Facultative feeding and consistency of trophic structure in marine soft-bottom macrobenthic communities.Marine Ecology Progress Series,2012,445:129-140.
[12] Carvalho S,Pereira P,Pereira F,de Pablo H,Vale C,Gaspar,M B.Factors structuring temporal and spatial dynamics of macrobenthic communities in a eutrophic coastal lagoon(Óbidos lagoon,Portugal).Marine Environmental Research,2011,71(2):97-110.
[13] Ge B M,Bao Y X,Cheng H Y,Li H H,Hu Z Y.The trophic functional groups and trophic levels of the macrobenthic community at the east tidal flat of Lingkun Island.Acta Ecologica Sinica,2008,28(10):4797-4804.
[14] Yuan X Z,Lu J J,Liu H.Distribution pattern and variation in the functional groups of Zoobenthos in the Chanjiang estuary.Acta Ecologica Sinica,2002,22(12):2054-2062.
[15] Li X Z,Li B Q,Wang H F.Community structure of macrobenthos in coastal water off Rushan,southern Shandong Peninsula,and the relationships with environmental factors.Acta Oceanologica Sinica,2009,28(5):81-93.
[16] State Oceanic Administration of China.The Specification for Marine Monitoring-Part 4:Seawater Analysis.Beijing:China Standards Press,1998:382-383.
[17] Gamito S,Furtado R.Feeding diversity in macroinvertebrate communities:a contribution to estimate the ecological status in shallow waters.Ecological Indicators,2009,9(5):1009-1019.
[18] Gamito S.Three main stressors acting on the Ria Formosa lagoonal system(Southern Portugal):physical stress,organic matter pollution and the land-ocean gradient.Estuarine,Coastal and Shelf Science,2008,77(4):710-720.
[19] Word J Q.The infaunal trophic index//Bascom W,ed.Southern California Coastal Water Research Project.California,1978:19-39.
[20] Borja A,Franco J,Pérez V.A Marine Biotic Index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments.Marine Pollution Bulletin,2000,40(12):1100-1114.
[21] Borja A,Muxika I,Franco J.The application of a Marine Biotic Index to different impact sources affecting soft-bottom benthic communities along European coasts.Marine Pollution Bulletin,2003,46(7):835-845.
[22] Kaiser M J,Ramsay K,Richardson CA,Spence F E,Brand A R.Chronic fishing disturbance has changed shelf sea benthic community structure.Journal of Animal Ecology,2000,69(3):494-503.
[23] Diaz R J,Cutter G R Jr,Dauer D M.A comparison of two methods for estimating the status of benthic habitat quality in the Virginia Chesapeake Bay.Journal of Experimental Marine Biology and Ecology,2003,285-286:371-381.
[24] Muxika I,Ibaibarriaga L,Saiz J I,Borja A.Minimal sampling requirements for a precise assessment of soft-bottom macrobenthic communities,using AMBI.Journal of Experimental Marine Biology and Ecology,2007,349(2):323-333.
[25] Bascom W,Mearns A J,Word J.Establishing boundaries between normal,changed and degraded areas//Bascom W,ed.Southern California Coastal Water Research Project Report.California,1978:81-94.
[26] Gaudencio Maria Jose,Cabral H N.Trophic structure of macrobenthos in the Tagus estuary and adjacent coastal shelf.Hydrobiologia,2007,587(1):241-251.
[27] Cacabelos E,Domínguez M,Troncoso JS.Trophic structure of soft-bottom macrobenthos in an inlet in north-western Spain.Journal of the Marine Biological Association of the United Kingdom,2009,89(3):439-447.
[28] You Z J,Tao L,Jiao H F,Shi H X,Lou D.A survey of macrobenthos in the Xiangshan Bay.Oceanologia et Limnologia Sinica,2011,42(3):431-435.
[29] Xin JH,Ren Y P,Xu B D,Zhang CL,Xue Y,Ji Y P.Macrozoobenthos functional groups in intertidal flat of northwest Jiaozhou Bay.Chinese Journal of Applied Ecology,2011,22(7):1885-1892.
[30] Li X Z,Yu H Y,Wang Y Q,Shuai L M,Zhang B L,Li X H.Study on the quantitative dynamics of macrobenthos in Jiaozhou Bay.Studia Marina Sinica,2002,44:66-73.
[31] Hussy K,St John M A,Bottcher U.Food resource utilization by juvenile Baltic cod Gadus morhua:a mechanism potentially influencing recruitment success at the demersal juvenile stage?Marine Ecology Progress Series,1997,155:199-208.
[32] Han J,Zhang Z N,Yu Z S.Macrobethic community structure in the southern and central Bohai Sea,China.Acta Ecologica Sinica,2004,24(3):531-537.
[33] Li B Q,Li X Z,Yu H Y,Wang H F,Wang JB,Xu F S,Zhang B L.Macrobenthic mollusca fauna and its relations to environmental factors in Jiaozhou Bay.Oceanologia et Limnologia Sinica,2005,36(3):193-198.
[34] Yang J M.A study on food and Trophic levels of Bohai Sea invertebrates.Modern Fisheries Information,2001,16(9):8-16.
[35] Rhoads D C,Young D K.The influence of deposit-feeding organisms on sediment stability and community trophic structure.Journal of Marine Research,1970,28:150-178.
[36] Bonsdorff E,Pearson T H.Variation in the sublittoral macrozoobenthos of the Baltic Sea along environmental gradients:a functional-group approach.Australian Journal of Ecology,1999,24(4):312-326.
[37] Wang JB,Li X Z,Wang H F.Ecological characteristics of dominant polychaete species from the Jiaozhou Bay.Acta Zoologica Sinica,2006,52(1):63-69.
[38] Callier M D,McKindsey C W,Desrosiers G.Multi-scale spatial variations in benthic sediment geochemistry and macrofaunal communities under a suspended mussel culture.Marine Ecology Progress Series,2007,348:103-115.
[39] Boaventura D,da Fonseca L C,Teles-Ferreira C.Trophic structure of macrobenthic communities on the Portuguese coast.A review of lagoonal,estuarine and rocky littoral habitats.Acta Oecologica,1999,20(4):407-415.
[40] Vale Cand Sundby B.Suspended sediment fluctuations in the Tagus estuary on semi-diurnal and fortnightly time scales.Estuarine,Coastal and Shelf Science,1987,25(5):495-508.
[41] Chícharo L,Regala J,Gaspar M,Alves F,Chícharo A.Macrofauna spatial differences within clam dredge-tracks and their implications for shortterm fishing effect studies.Fisheries Research,2002,54(3):349-353.
[42] Saito H,Ueno M,Hayashi I.Temporal fluctuation in the abundance of a semelid bivalve,Theora fragilis(A.Adams)in Maizuru Bay,Sea of Japan.Hydrobiologia,1998,375-376:151-163.
[43] Horikoshi M.Macrobenthic communities in Japanese coastal waters//Oceanographic Society of Japan,ed.Coastal oceanography of Japanese islands(Suppl vol).Tokyo:Tokai University Press,1990:283-311.
[44] Cai L Z,Li H M,Liu J J,Lin P.Distribution and polluting effects on Capitella capitata,Nephtys oligobranchia,Tharyx sp.on the intertidal mudflats in Shenzhen Estuary.Acta Ecologica Sinica 2001,21(10):1649-1653.
[45] Wijsman J W M,Herman P M J,Gomoiu M T.Spatial distribution in sediment characteristics and benthic activity on the Northwestern Black Sea shelf.Marine Ecology Progress Series,1999,181:25-39.
[46] Nanami A,Saito H,Akita T,Motomatsu K,Kuwahara H.Spatial distribution and assemblage structure of macrobenthic invertebrates in a brackish lake in relation to environmental variables.Estuarine,Coastal and Shelf Science,2005,63(1/2):167-176.
[47] Ysebaert T,Meire P,Coosen J,Essink K.Zonation of intertidal macrobenthos in the estuaries of Schelde and Ems.Aquatic Ecology,1998,32(1):53-71.
[48] Xu Q Z,Li R X,Wang Z L,Fan SL,Wang Z X.Macrobenthos distribution of the South Yellow Sea in summer.Advances in Marine Science,2009,27(3):393-399.
[49] Chícharo,M A,Chícharo L,Amaral A,Condinho S,Gaspar M.Chronic effects of dredging-induced stress on the clam(Spisula solida):nucleic acid and lipid composition.Fisheries Research,2003,63(3):447-452.
[50] Muniz P,Pires A M S.Trophic structure of the polychaetes in the São Sebastião Channel(southeastern Brazil).Marine Biology,1999,134(3):517-528.
[51] Liu L S,Meng W,Li X Z,Li Z C,Zheng B H,Lei K,Li Z Y.Studies on macrobenthos in the northern waters of Liaodong Bay:Ⅱ.Biodiversity and community structure.Research of Environmental Sciences,2009,22(2):155-161.
[52] Zhang Z N.Some progress of the study on the ecosystem dynamics for benthic-pelagic coupling.Journal of Ocean University of Qingdao,2000,30(1):115-122.
[53] Gaston G R,Rakocinski C F,Brown S S,Cleveland C M.Trophic function in estuaries:response of macrobenthos to natural and contaminant gradients.Marine and Freshwater Research,1998,49(8):833-846.
[54] Blanchet H,de Montaudouin X,Chardy P,Bachelet G.Structuring factors and recent changes in subtidal macrozoobenthic communities of a coastal lagoon,Arcachon Bay(France).Estuarine,Coastal and Shelf Science,2005,64(4):561-576.
[55] Fujii T.Spatial patterns of benthic macrofauna in relation to environmental variables in an intertidal habitat in the Humber estuary,UK:developing a tool for estuarine shoreline management.Estuarine,Coastal and Shelf Science,2007,75(1/2):101-119.
[56] Kuosa H,Autio R,Kuuppo P,SetäläO,Tanskanen S.Nitrogen,silicon and zooplankton controlling the Baltic spring bloom:an experimental study.Estuarine,Coast and Shelf Science,1997,45(6):813-821.
[57] Bartoli M,Nizzoli D,Viaroli P,Turolla E,Castaldelli G,Fano E A,Rossi R.Impact of Tapes philippinarum farming on nutrient dynamics and benthic respiration in the Sacca di Goro.Hydrobiologia,2001,455(1/3):203-212.
参考文献:
[6] 何志辉.淡水生态学.北京:中国农业出版社出版,2000:101-102.
[13] 葛宝明,鲍毅新,程宏毅,李欢欢,胡知渊.灵昆岛东滩潮间带大型底栖动物功能群及营养等级构成.生态学报,2008,28(10):4797-4804.
[14] 袁兴中,陆健健,刘红.长江口底栖动物功能群分布格局及其变化.生态学报,2002,22(12):2054-2062.
[16] 国家海洋局.海洋监测规范第4部分:海水分析.北京:中国标准出版社,1998:382-383.
[28] 尤仲杰,陶磊,焦海峰,施慧雄,楼丹.象山港大型底栖动物功能群研究.海洋与湖沼,2011,42(3):431-435.
[29] 辛俊宏,任一平,徐宾铎,张崇良,薛莹,纪毓鹏.胶州湾西北部潮滩湿地大型底栖动物功能群.应用生态学报,2011,22(7):1885-1892.
[30] 李新正,于海燕,王永强,帅莲梅,张宝琳,李笑红.胶州湾大型底栖动物数量动态的研究.海洋科学集刊,2002,44:66-73.
[32] 韩洁,张志南,于子山.渤海中、南部大型底栖动物的群落结构.生态学报,2004,24(3):531-537.
[33] 李宝泉,李新正,于海燕,王洪法,王金宝,徐凤山,张宝琳.胶州湾底栖软体动物与环境因子的关系.海洋与湖沼,2005,36(3):193-198.
[34] 杨纪明.渤海无脊椎动物的食性和营养级研究.现代渔业信息,2001,16(9):8-16.
[37] 王金宝,李新正,王洪法.胶州湾多毛类环节动物优势种的生态特点.动物学报,2006,52(1):63-69.
[44] 蔡立哲,厉红梅,刘俊杰,林鹏.深圳河口泥滩三种多毛类的数量季节变化及污染影响.生态学报,2001,21(10):1649-1653.
[48] 徐勤增,李瑞香,王宗灵,范士亮,王宗兴.南黄海夏季大型底栖动物分布现状.海洋科学进展,2009,27(3):393-399.
[51] 刘录三,孟伟,李新正,李子成,郑丙辉,雷坤,李中宇.辽东湾北部海域大型底栖动物研究:Ⅱ.生物多样性与群落结构.环境科学研究,2009,22(2):155-161.
[52] 张志南.水层-底栖耦合生态动力学研究的某些进展.青岛海洋大学学报:自然科学版,2000,30(1):115-122.
