高寒山区一年生混播牧草生态位对密度的响应
2013-09-07赵成章盛亚萍
赵成章,张 静,盛亚萍
(西北师范大学地理与环境科学学院,甘肃省湿地资源保护与产业发展工程研究中心,兰州 730070)
生态位是现代生态学研究的一个主要内容,指种群在时间、空间的位置以及在群落的地位和功能作用[1]。Silvertown[2]认为生态位同种间竞争密不可分,是评价种内种间关系及种群在群落中地位的重要手段[3]。随着生态位理论在物种间关系、生物多样性和群落结构等方面的广泛应用,逐步成为生态学研究中非常活跃的领域之一[4]。近年来关于各类植物群落物种生态位的研究日益普遍,尤其在土壤盐分[5]、含水量[6]、有机质[7]和pH值[8]对植物种群生态位分化影响等方面取得了重要进展,丰富和发展了生态位理论。混播组合试验中混生物种相对密度的变化将影响植物的资源利用效率,促使植物个体生长形态发生适应性改变,植物高度的增加速率和生物量的贡献值发生变化,形成不同的空间生态位,导致物种种内种间竞争强度和转化存在差异[9]。因此,认识密度梯度上混生种群生态位宽度和重叠变化规律,是了解植物种群竞争协同机制的关键[10]。
高寒山区气候寒冷,牧草生长季短、产量低,发展优质高产的人工草地是解决冷季草畜供求矛盾、保障草地畜牧业可持续性的重要途径,研究密度制约下混播草地牧草的生态位宽度和生态位重叠及其转化规律,探讨混播草地牧草密度的最佳搭配模式,对于提高草地干物质产量和资源利用效率具有重要意义。国内学者关于高寒山区牧草种间竞争关系[11]、放牧干扰[12]和土壤盐分[13]对牧草生态位的影响等方面均进行了较深入研究。但密度变化对高寒山区人工草地混播牧草生态位宽度和生态位重叠格局,以及牧草生态位对种内种间竞争响应的适应性转化机理仍然不明晰。鉴于此,本文采用取代系列实验方法,在石羊河上游高寒山区建立1年生禾本科牧草燕麦和豆科牧草毛苕子混播草地,研究了密度制约下混播牧草生态位宽度和生态位重叠的动态规律,探讨了生态位与种间竞争之间的相互关系。
1 研究地区与研究方法
1.1 研究区概况
实验地位于石羊河上游的甘肃省肃南县皇城镇绵羊育种场(37°58'26″N,101°47'34″E),海拔2530 m,具有大陆性气候和山地垂直气候特征。年均温0℃,最热和最冷月气温分别为12—15℃和-13—-11℃,≥0℃的年积温2450—2600℃。年均降水量350 mm左右,主要集中在6—9月,年20 cm口径水面蒸发量1500—1800 mm,相对湿度65%。年日照时数2200—2800 h,无霜期80 d左右。土壤以山地栗钙土为主,有机质36.5 mg/g,全氮2.2 mg/g,全磷0.6 mg/g,全钾28.3 mg/g,代换量15.1 m·e·100 g-1土[11]。试验区地处山地荒漠草原与干旱草原过渡带,具备发展人工草地的气候和灌溉条件,已经形成了以禾本科牧草为主的人工饲草料种植模式,但是人工草地品种单一、生产能力低下,具有进一步提升的空间和潜力。
1.2 试验设计
运用取代试验方法,在单位面积植株总密度一定条件下,按1年生燕麦(Avena sativa L.)和毛苕子(Vicia villosa Roth)的密度比例设 KY(1∶0)、A(8∶2)、B(6∶4)、C(5∶5)、D(4∶6)、E(2∶8)和 KM(0∶1)7 种种植方式。所有小区总密度为300株/m2,浓硫酸处理打破毛苕子种子硬实,根据种子纯净度、千粒重以及室内发芽实验结果,确定两种牧草单播和混播的播种量(KY和KM的用种量分别为:9 g/m2和19 g/m2,其余依次类推)。小区面积2 m×5 m,3次重复,小区纵横间距均为0.50m,共21个小区,随机区组排列。2010年6月5—12日整理实验小区,去除地面植被,深翻30cm内土层,一次性施足底肥,平整地面。2010年6月15日播种,播种采用行播,行距20cm,每行均匀播种燕麦与毛苕子,播种深度2—4cm,实验过程中不施肥,分别于7月10日、8月5日、8月20日人工除草3次,7月20日、8月10日分别采用喷灌方式对供试草地进行2次3000m3/hm2的灌溉。
在2010年7月17日—8月27日隔10d取样1次,记录牧草物候期,每个处理重复3次取样,各小区取样面积50 cm×50 cm,齐地面刈割,收获样品分别记录每样方自然状态下20株燕麦和毛苕子从地面至植株最上部的植株高,然后按区组顺序将材料分别置于65℃烘箱8 h至恒重后记录两个物种干重。
1.3 数据分析
本文以所调查的样方为资源状态,使各指标综合反映物种对多种资源的利用,同时也反映植物种的空间关系[14]。以混播草地中燕麦与毛苕子的相对比例和牧草物候期为资源梯度,以株高和生物量为资源维进行生态位宽度和生态位重叠的计测。重要值采用相对密度和相对盖度之和。
1)用Levins的生态位宽度指数计测[15]:

式中,BLi是物种i的生态位宽度,pij为物种i对第j个资源状态下的重要值占该种在所有资源状态中重要值总数的比例,r为资源水平数。
2)用Pianka的生态位重叠指数计测[16]:

式中,Oik为物种i和物种j的生态位重叠,nij和nkj为种i和种k在第j个资源状态下的重要值占该种在所有资源状态中重要值总数的比例,r为资源水平数。
2 结果分析
2.1 混播草地燕麦与毛苕子的株高
在牧草生长的每个时间段,各混播组燕麦和毛苕子株高存在显著性差异(P<0.01),且在密度梯度上呈现明显的规律性。具体表现为:从密度组KY到KM随着燕麦相对密度的减小和毛苕子相对密度的增加,燕麦和毛苕子株高呈减小趋势(表1)。
2.2 混播草地燕麦与毛苕子的分蘖(枝)数
在牧草生长的每个时间段,各混播组燕麦和毛苕子分蘖(株)数存在显著性差异(P<0.01),且在密度梯度上呈现明显的规律性。具体表现为:从混播组A到E随着燕麦相对密度的减小和毛苕子相对密度的增加,燕麦分蘖数和毛苕子分枝数呈增加趋势(表2);燕麦单播(KY)草地和毛苕子单播(KM)草地中牧草分蘖(枝)数均小于混播草地(表2)。
2.3 混播草地燕麦与毛苕子的生态位宽度
生态位宽度表示物种对资源的利用程度和环境适应情况。各混播组燕麦和毛苕子的生态位宽度存在显著性差异(P<0.01),且在密度梯度上呈现明显的规律性。具体表现为:从混播组A到E随着燕麦相对密度的减小和毛苕子相对密度的增加,燕麦生态位宽度逐渐增加,毛苕子生态位逐渐减小,二者差值平均从0.40逐渐减小到0.09(图1)。随牧草生长时间延长,同一混播实验燕麦生态位宽度呈减小趋势(图1),混播组A和B中燕麦生态位宽度减小的速率大于混播组C、D和E;毛苕子生态位宽度在燕麦出苗期最低(平均为0.57),燕麦分蘖期迅速增加达到最高(平均为0.99),从燕麦分蘖期到燕麦成熟期以较小速率逐渐减小(图1)。
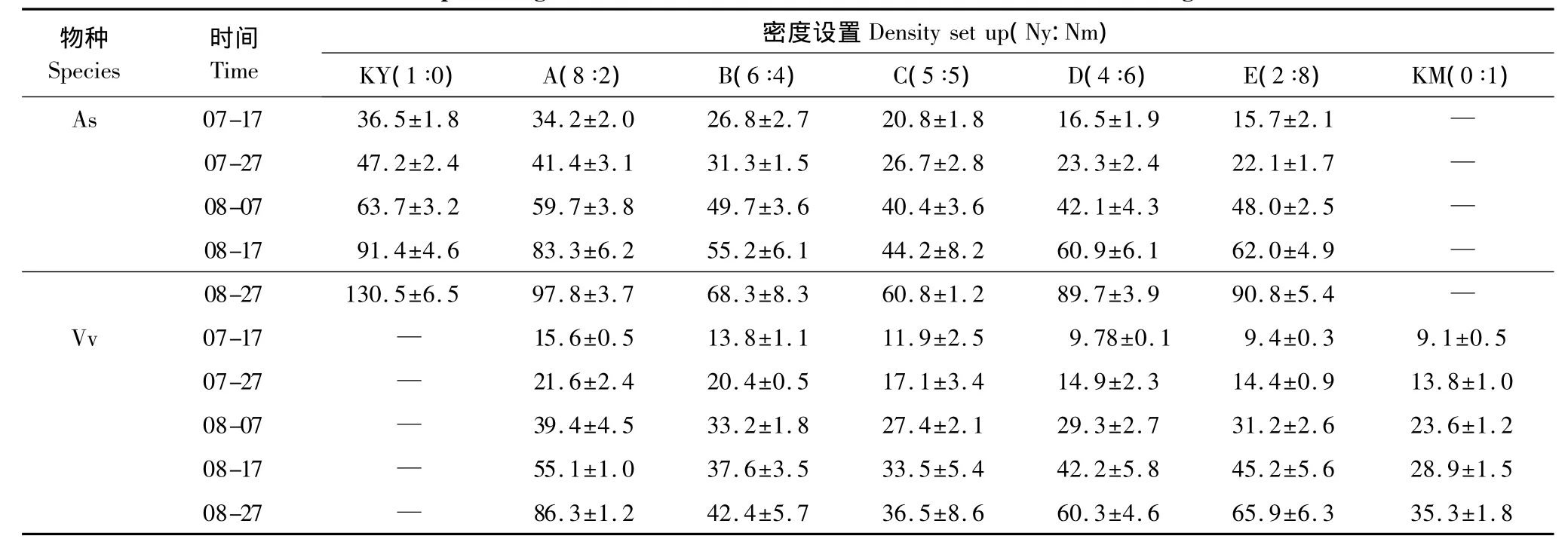
表1 混播草地燕麦与毛苕子株高Table1 The plant height of Avena sativa L.and Vicia vilosa Roth in mixed-seeding meadow
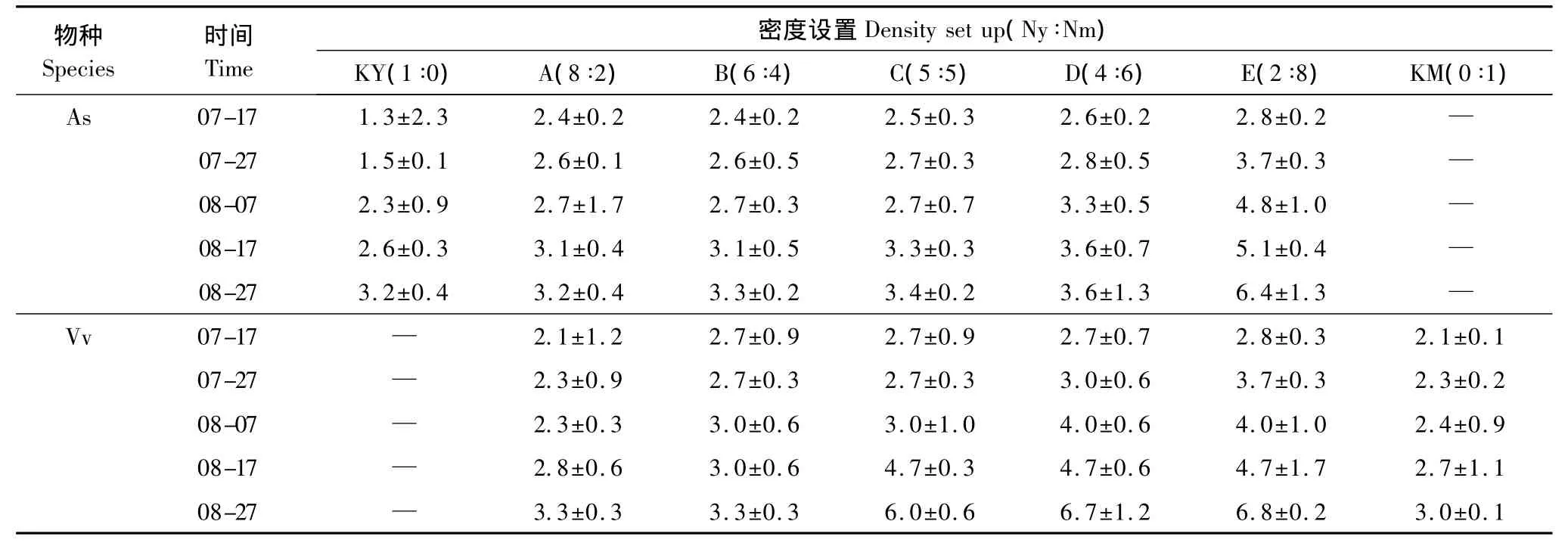
表2 混播草地燕麦与毛苕子分蘖(枝)数Table2 The tiller(branch)number of Avena sativa L.and Vicia vilosa Roth in mixed-seeding meadow
2.4 混播草地燕麦与毛苕子的生态位重叠
生态位重叠指两个或更多物种对一个资源或多个资源的共同利用。各混播组燕麦和毛苕子的生态位重叠存在显著性差异(P<0.01),且在密度梯度上呈现明显的规律性。具体表现为:从混播组A到E随着燕麦相对密度的减小和毛苕子相对密度的增加,混播草地中燕麦和毛苕子的生态位重叠平均由0.81逐渐增大到0.99(图2)。在牧草生长的各个时间段,除混播组A外其余混播组燕麦和毛苕子的生态位重叠先减小后增大(图2)。
3 讨论
3.1 密度对混播牧草生态位宽度的影响
Abrams[17]认为可通过植物利用资源的状况反映种群间相互关系,生态位宽度是度量植物对资源环境利用状况的尺度,生态位宽度越大,该物种利用资源的能力越强,其分布幅度就越大;反之则说明该种特化程度较高,其分布只能局限在特定的区域[18]。研究区降水多集中在6—9月,占全年降水的55%—70%,供试的人工草地在整地时一次性足够施用底肥,且牧草生长过程中根据土壤墒情合理进行了人工灌溉,因此1年生人工草地的水肥条件能够满足牧草生长需求,所以本试验中决定植物生长的主要因素是光、温和植物的生理、形态学特征[19]。本研究中各混播组燕麦与毛苕子生态位宽度介于0.5—1之间,随牧草生长时间的延长2者的生态位宽度发生了复杂的转变。混播草地中早萌发或早定植的植物往往具有明显的竞争优势[20],由于混播草地中禾本科牧草燕麦的出苗时间早于豆科牧草毛苕子,所以人工草地建植初期各混播组燕麦的生态位宽度大于毛苕子(图1)。燕麦从出苗期开始一直占据着明显的高度优势(表1),行内对毛苕子形成较强的荫蔽作用,由于匍匐生长型毛苕子具有非常发达的枝叶系统和明显的觅光行为,所以毛苕子逐步向行间扩展以便获得足够的光资源,毛苕子的生态位宽度逐步增加并在7月下旬且超过燕麦(图1)。燕麦在开花期随着上层叶片向行间扩展,对行间毛苕子的枝叶形成荫蔽作用,进一步影响了毛苕子的生长,毛苕子一旦找到林窗就会产生带有大型叶片的缠绕枝条,迅速爬升到林冠层,并形成自己的藤本层,致使毛苕子个别小枝的高度超过了燕麦,毛苕子通过横向和纵向扩展继续保持了生态位宽度优势。总而言之,混播草地中毛苕子在燕麦直立茎杆的支持下,毛苕子生态位宽度在牧草生长过程中逐渐增加,在毛苕子的竞争限制下,燕麦的生态位宽度在牧草生长过程中逐渐减小。
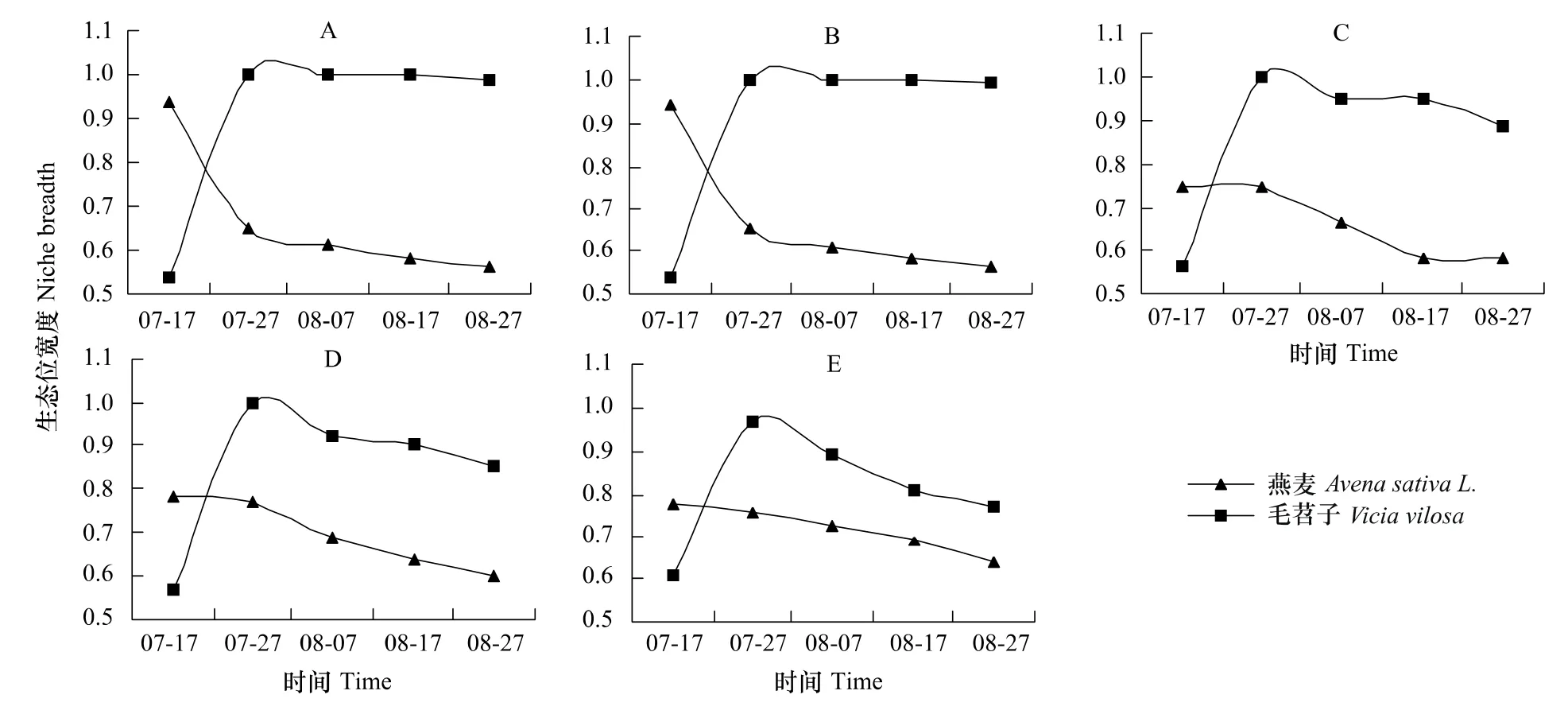
图1 混播草地中燕麦与毛苕子的生态位宽度Fig.1 The niche breadth of Avena sativa L.and Vicia vilosa Roth in mixed-seeding meadow

图2 混播草地中燕麦与毛苕子的生态位重叠Fig.2 The niche overlap of Avena sativa L.and Vicia vilosa Roth in mixed-seeding meadow
植物地上部分形态的可塑性变化主要是由于植物冠层叶片互相遮荫而造成的光环境改变所致[21],较强的荫蔽度导致植物所能接受到的光质即红光和远红光之比(R∶FR)有所减小,促使植物获取顶端优势,禾草类植物的分蘖数降低[22],匍匐草本植物的匍匐枝在地上迅速延伸[23]。密度制约是植物种群调节的重要机制之一[9],从混播组A到混播组E随着燕麦相对密度的逐渐减小,燕麦种内植株间的荫蔽度逐渐减弱,所以燕麦不需要明显的增加株高来争夺光资源,而以增加分蘖数的模式横向扩展来获得更多光资源(表1),生态位宽度逐渐增加。与此同时,在密度梯度上,虽然混播草地中燕麦对毛苕子的种间荫蔽度逐渐减弱,但是伴随着毛苕子的相对密度逐渐增大,毛苕子在较低层更加紧密的缠绕生长,其枝叶的横向扩展空间逐步缩小;混播草地毛苕子种内荫蔽度逐渐增强,使毛苕子借助叶柄伸长和增加分枝数来获得更多光资源(表2),同时纵向扩展效果逐渐减弱(表1),最终毛苕子的生态位宽度逐渐减小。总而言之,混播草地中随着牧草自身相对密度的增加生态位宽度减小,即牧草生态位宽度与密度呈负相关关系(SMA斜率小于1)。本研究关于密度制约下,燕麦与毛苕子生态位宽度的变化趋势支持密度影响和胁迫下牧草生态位会向偏离其他物种最大利用位置的方向移动[24]的观点。
3.2 密度对混播牧草生态位重叠的影响
当两个物种共同利用或占有某一资源(空间、营养成分、食物等)时,就会发生生态位重叠[18]。较高的生态位重叠说明植物间在资源利用上竞争强烈,对植物群落的结构和功能具有显著影响;而较低的生态位重叠说明各种植物种群可以充分的分享群落环境资源,主要种群之间的关系比较协调和平衡,能够相互适应,群落往往处于相对稳定的状态[1,25]。物种间生态位重叠除与环境(生物)条件相关外,还与物种间生态学特性和种间竞争强弱密切相关[26]。本研究中各混播组燕麦与毛苕子生态位重叠介于0.7—1之间,在牧草生长发育的不同时期,虽然各混播组燕麦和毛苕子的生态位重叠存在显著性差异(P<0.01),但是在密度梯度上2者的生态位重叠仍然呈现明显的规律性。随着燕麦相对密度的减小和毛苕子相对密度的增加,混播草地两种牧草应对种内、种间竞争的资源分配策略发生了调整,从燕麦相对密度较高的A组到毛苕子相对密度较高的E组,燕麦的生态位宽度逐渐增大,毛苕子生态位宽度逐渐减小,二者的差值减小(图1),使燕麦与毛苕子重叠度逐渐增强(图2),毛苕子为了获得更多光资源,依赖自身匍匐蔓生以燕麦直立茎秆为攀援体向上生长的特性,提高了混播草地资源利用水平,加剧了竞争。
随牧草生长时间延长各混播组燕麦与毛苕子种间竞争逐渐激烈,加剧了群体效应,导致牧草生态位重叠发生变化。人工草地建植初期,虽然燕麦与毛苕子个体较小,在条播种植模式下,行内毛苕子并没有扩散,上繁草燕麦对下繁草毛苕子形成完全遮阴,导致燕麦与毛苕子生态位重叠较高。较高的生态位重叠说明混播草地中结构和性质的不稳定,会促使牧草的形态特征在生长过程中发生变化,向群落稳定的方向转移或者存在转移的潜力,因此燕麦株高从出苗期到拔节期迅速增加(表1),行内燕麦对毛苕子荫蔽度增强,毛苕子为了获得更多光资源会向行间扩展,使混播草地空间生态位发生明显分离,燕麦与毛苕子重叠度有所减小。随着燕麦开花期和成熟期上层叶片对行间毛苕子枝叶遮阴程度的增强,毛苕子必须攀援燕麦向上生长,此时燕麦与毛苕子空间生态位再次重叠,并逐步增强。
综上所述,环境条件的改变降低了物种的生存适合度,导致物种对资源利用的转变和生态位的分化[27]。植物通过株高和分蘖(枝)数的可塑性调节来适应物候条件和密度制约的变化,不仅为自身生长发育提供了保护机制,同时也提高了资源利用效率[28],形成相互稳定相互影响的生态系统。本文探索了密度制约下高寒山区燕麦与毛苕子混播草地中牧草生态位的动态变化,及其对种间与种内竞争转化的响应,关于混播牧草的稳定性和适合度的变化还有待于进一步研究。
[1] Li D W,Zhang W H,Ren Z Z.Niche characteristics of dominant populations of S ophora davidii community in loess gully region.Chinese Journal of Applied Ecology,2005,16(12):2231-235.
[2] Silvertown J W.Plants in limestone pavements:tests of species interaction and niche separation.Journal of Ecology,1983,71(3):819-828.
[3] Wang Z L,Chen Y,Chen X M,Wang SY,Zhao J J.Temporal-spatial niches of Chinese White Wax Scale insect(Ericerus pela)and its three dominant parasitoid wasps.Acta Ecologica Sinica,2011,31(4):914-922.
[4] Wang P,Zhou D W,Zhang B T.Coexistence and inter-specific competition in grass-legume mixture.Acta Ecologica Sinica,2009,29(5):2560-2567.
[5] Li F,Xie Y H,Chen X S,Deng Z M,Li X.Composition of aquatic plants and their niche characteristics in wetlands of the Yellow River Delta.Acta Ecologica Sinica,2009,29(11):6257-6265.
[6] Liu JZ,Chen Y N,Zhang Y M.Niche characteristics of plants on four environmental gradients in middle reaches of Tarim River.Chinese Journal of Applied Ecology,2004,15(4):549-555.
[7] He Q,Cui B S,Zhao X S,Fu H L.Niches of plant species in wetlands of the Yellow River Delta under gradients of water table depth and soil salinity.Chinese Journal of Applied Ecology,2008,19(5):969-975.
[8] Yang Z H,Fang E T,Liu H J,Li A D,Xu X Y.Effect of water table to niche of plant population at Minqin oasis fringe.Acta Ecologica Sinica,2007,27(11):4900-4906.
[9] Li L,Zhou D W,Sheng L X.Density dependence-determined plant biomass allocation pattern.Chinese Journal of Ecology,2011,30(8):1579-1589.
[10] Watkinson A R.Density-dependence in single-species populations of plants.Journal of Theoretical Biology,1980,83(2):345-357.
[11] Sheng Y P,Zhao C Z,Gao F Y,Zhang J X,Shi F X,Qu Y.Inter-specific competition between Avena sativa and Vicia vilosa in mixed sowing grassland in alpine mountainous region.Chinese Journal of Ecology,2011,30(11):2437-2441.
[12] Dong Q M,Zhao X Q,Ma Y T,Li Q Y,Shi JJ,Wang Y L,Sheng L,Yan M Y.Niche of main plant populations on a warm-seasonal pastureland of alpine Kobrecia parva meadow.Chinese Journal of Ecology,2006,25(11):1323-1327.
[13] Wang X,Hu Y K,Rehemudula A D L,Liu Y Y,Li K H,Wang JY.Effects of soil salt on the niche of main plant species in alpine meadow.Chinese Journal of Applied Ecology,2008,19(7):1448-1454.
[14] Pielou E C.An Introduction to Mathematical Ecology.New York:Wiley-Interscience Press,1969.
[15] Levins R.Evolution in Changing Environments:Some Theoretical Explorations.Princeton:Princeton University Press,1968.
[16] Pianka E R.The structure of lizard communities.Annual Review of Ecology and Systematics,1973,4:53-74.
[17] Abtams P A.Alternative models of character displacement and niche shiftⅠ.Adaptive shifts in resource use when there is competition for nutritionally nonsubstitutable resources.Evolution,1987,41(3):651-661.
[18] Xu Z G,He Y,Yan B X,Song C H.Niche characteristics of typical marsh wetland plant populations in Sanjiang Plain.Chinese Journal of Applied Ecology,2007,18(4):783-787.
[19] Li B.Plant Competition-Experiment Research of Interactional between Crop and Weeds.Beijing:China Higher Education Press:Berlin Heidelberg:Springer-Verlag,2001.
[20] Firbank L G,Watkinson A R.On the Effects of Competition:From Monocultures to Mixtures//Grace JB,Tilman D,eds.Perspectivve on Plant Competition.London:Academic Press,1990:165-192.
[21] Schmitt J,Wulff R D.Light spectral quality,phytochrome and plant competition.Trends in Ecology and Evolution,1993,8(2):47-50.
[22] Casal J J,Sanchez,R A,Gibson D.The significance of changes in the red/far-red ratio,associated with either neighbor plants or twilight,for tillering in Lolium multiflorum Lam.New Phytologist,1990,116(4):565-572.
[23] Huber H.Architectural Plasticity of Stoloniferous and Erect Herds in Response to Light Climate[D].Utrecht:University,1997.
[24] Zhang L J,Yue M,Zhang Y D,Gu F X,Pan X L,Zhao GF.Analysis on the niche of the main plant species of Oasis-deseart ecotone in Fukang of Xinjiang.Acta Ecologica Sinica,2002,22(6):969-972.
[25] Liao B W,Li W,Zheng SF,Chen Y J,Zhong C R,Huang Z Q.Niche of several mangrove species in Dongzhai Hainan Island.Chinese Journal of Applied Ecology,2005,16(3):403-407.
[26] Tilman G D.Plant dominance along an experimental nutrient gradient.Ecology,1984,65(5):1445-1453.
[27] Tilman D.Causes,consequences and ethics of biodiversity.Nature,2000,405(6783):208-211.
[28] Silvertown J.Plant coexistence and the niche.Trends in Ecology and Evolution,2004,19(11):605-611.
参考文献:
[1] 李登武,张文辉,任争争.黄土沟壑区狼牙刺群落优势种群生态位研究.应用生态学报,2005,16(12):2231-235.
[3] 王自力,陈勇,陈晓鸣,王绍云,赵杰军.白蜡虫及其3种优势寄生蜂的时空生态位.生态学报,2011,31(4):914-922.
[4] 王平,周道玮,张宝田.禾-豆混播草地种间竞争与共存.生态学报,2009,29(5):2560-2567.
[5] 李峰,谢永宏,陈心胜,邓正苗,李旭.黄河三角洲湿地水生植物组成及生态位.生态学报,2009,29(11):6257-6265.
[6] 刘加珍,陈亚宁,张元明.塔里木河中游植物种群在四种环境梯度上的生态位特征.应用生态学报,2004,15(4):549-555.
[7] 贺强,崔保山,赵欣胜,付华龄.水、盐梯度下黄河三角洲湿地植物种的生态位.应用生态学报,2008,19(5):969-975.
[8] 杨自辉,方峨天,刘虎俊,李爱德,徐先英.民勤绿洲边缘地下水位变化对植物种群生态位的影响.生态学报,2007,27(11):4900-4906.
[9] 黎磊,周道玮,盛连喜.密度制约决定的植物生物量分配格局.生态学杂志,2011,30(8):1579-1589.
[11] 盛亚萍,赵成章,高福元,张军霞,石福习,屈勇.高寒山区混播草地燕麦和毛苕子种间的竞争关系.生态学杂志,2011,30(11):2437-2441.
[12] 董全民,赵新全,马玉寿,李青云,施建军,王彦龙,盛丽,闫明毅.高寒小嵩草草甸暖季草场主要植物种群的生态位.生态学杂志,2006,25(11):1323-1327.
[13] 王鑫,胡玉昆,热合木都拉·阿迪拉,柳妍妍,李凯辉,王吉云.土壤盐分对高寒草甸主要植物生态位的影响.应用生态学报,2008,19(7):1448-1454.
[14] 余世孝.数学生态学引论.北京:科学技术文献出版社,1995.
[18] 徐治国,何岩,闫百兴,宋长春.三江平原典型沼泽湿地植物种群的生态位.应用生态学报,2007,18(4):783-787.
[19] 李博.植物竞争—作物与杂草相互作用的实验研究.北京:高等教育出版社;海德堡:施普林格出版社,2001.
[24] 张林静,岳明,张远东,顾峰雪,潘晓玲,赵桂仿.新疆阜康绿洲荒漠过渡带主要植物种的生态位分析.生态学报,2002,22(6):969-972.
[25] 廖宝文,李玟,郑松发,陈玉军,钟才荣,黄仲琪.海南岛东寨港几种红树植物种间生态为研究.应用生态学报,2005,16(3):403-407.
