中亚热带人工针叶林生态系统碳通量拆分差异分析
2013-09-07王绍强王辉民仪垂祥刘允芬
黄 昆,王绍强 ,王辉民,仪垂祥,周 蕾,刘允芬,石 浩
(1.中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101;2.中国科学院大学,北京 100049;3.中国科学院千烟洲红壤丘陵综合开发试验站,泰和 343700;4.美国纽约城市大学皇后学院,纽约 11367美国)
作为目前唯一的直接测定陆地生态系统与大气之间CO2净交换量的方法,涡度协方差通量观测已经成为监测和理解陆地生态系统碳循环不可或缺的工具。据全球碳通量观测研究网络(FLUXNET)统计,截至2010年全球已经建立起超过500多个通量观测站点对不同生态系统类型进行长期的CO2和水热通量的观测研究[1]。通量观测主要目标在于采用可靠的拆分方法将直接观测获得的净生态系统交换(NEE)数据分离为生态系统与大气之间的主要通量:总初级生产力(GPP)和生态系统总呼吸(Re),用以深入研究生态系统碳循环过程以及校验生态系统模型[2-3]。NEE拆分的方法主要包括以下三类:(1)基于同步观测的气象数据的方法[3-4];(2)基于稳定同位素观测的方法[5-6];(3)基于箱式法为主的呼吸观测方法等等[7-8]。
基于NEE和常规气象数据的方法作为最常用的拆分方法,又可以根据使用函数或模型的不同分为几个小类,包括:平均日变化法、查表法、非线性回归模型、人工神经网络以及遗传算法等等[9-10]。广泛采用的“非线性回归模型”拆分方法(Method 1)利用NEE和气象要素数据之间具备生理生态学意义的非线性关系,构建NEE和气象要素的回归模型,并利用完整时间序列的气象要素数据对缺失NEE数据进行插补,同时实现对于NEE的拆分。其基本原理是:植被在夜间不进行光合作用,因此涡度相关通量观测的夜间NEE数据等同于夜间总生态系统呼吸(Re),利用有效的夜间NEE数据和空气温度或表层土壤温度建立非线性模型[10],可获得针对特定生态系统的Re对温度的响应函数,再结合白天观测的相应气象数据即可估算出白天的Re[11-12]。最后,根据NEE、Re以及GPP之间的代数关系可推出GPP。但是,涡度相关通量数据质量控制、质量保证过程往往使得夜间有效NEE比例较低,进而导致采用非线性回归的方法估算白天Re可能存在较大误差。因此,利用白天NEE对光合有效辐射(PAR)的光响应曲线函数关系(Method 2)进行白天生态系统呼吸直接估算的方法得到了发展[13-15],并以该模型的截距作为白天生态系统呼吸量的均值,据此以拆分NEE[15-16]。
Reichstein等[3]认为利用夜间NEE与气象要素之间的非线性关系外推至白天呼吸的方法进行Re估算的方法亦存在显著的缺点,包括:(1)全年有效数据的夜间观测数据可能不足50%,进而显著影响对Re的准确估算[10,16];(2)利用全年的有效夜间数据进行非线性拟合获得的相应参数可能无法反映短时间尺度Re对环境变量的响应特征,而如果利用短时间尺度的少量有效夜间数据进行相应拟合又可能无法获得可靠的模型参数[3];(3)在白天光照情况下,植物叶片暗呼吸受到抑制(即“kok效应”),其呼吸速率可能低于夜间,进而导致利用夜间Re与温度的关系外推白天Re时存在高估可能接近15%[16-17]。另外,白天的生态系统呼吸值(Re)可能并不满足与温度有关的经验关系[18-19]。
国外已有研究指出:利用上述两种非线性回归方法获得的年尺度生态系统碳通量(GPP和Re)在大多数通量站点存在较好的一致性[20-21]。尽管目前国内多数研究普遍采用非线性回归(Method 1)方法进行NEE的拆分,依然有必要分析不同拆分方法对生态系统碳通量(GPP和Re)评估的影响[4]。
我国南方人工林占全国人工林面积的一半以上,深入研究我国中亚热带地区人工针叶林碳源汇时空特征及其气候异常的响应,有助于准确评价气候变化对我国人工林碳汇的影响。本研究利用中国陆地生态系统通量观测研究网络(ChinaFLUX)江西泰和县千烟洲生态试验站[22]的2003—2009年的半小时涡度通量和气象观测数据,分析上述不同NEE拆分方法下生态系统碳通量(GPP和Re)拆分结果的差异,了解造成这种差异的环境影响机制,探讨如何提高中亚热带人工针叶林生态系统NEE拆分的准确性。
1 材料和方法
1.1 研究站点简介
千烟洲生态试验站(QYZ)地处江西省泰和县,属亚热带季风气候。通量观测塔位于试验站西南部上松塘、西角塘和平坑垅等小流域的交汇点(26°44'48″N,115°04'13″E,110.8m),塔周围大约 1 km2范围内森林覆盖度高达90%,近100 km2范围内森林覆盖近70%,林分为1985年前后营造的人工林[23-24]。多年平均气温为17.9℃,多年平均降水1542.4 mm,土壤类型为红壤,主要植物种为马尾松(Pinus massoniana)、湿地松(Pinus elliottii Engelm)和杉木(Cunninghamia lanceolata),生态系统类型为人工针叶林,通量观测从2002年10月至今。
1.2 通量和气象数据的获取与处理方法
1.2.1 通量和气象数据的获取方法
千烟洲站(QYZ)的通量观测主要以开路涡度相关系统为主,辅以常规气象要素和植物生理生态要素以及植被清查等。开路涡度相关系统由三维超声风速仪(CAST3,Campell,USA)和开路式CO2/H2O红外气体分析仪(Li-7500,LiCor Inc.,USA)组成,原始采样频率为 10 Hz,由数据采集器(CR5000,Campbell Scientific,USA)进行数据采集和在线计算,并储存平均周期为30 min的CO2通量数据。常规气象要素测定包括总辐射、净辐射、光合有效辐射、空气温度/湿度、风速/风向、降水量、土壤温度/湿度、土壤热通量等。通量和其他常规观测数据均通过数据传输系统下载到计算机,以进行数据的存储和后续的处理及分析[24]。
1.2.2 通量和气象数据的处理方法
利用非线性回归模型拆分NEE需要用到完整时间序列的气象要素数据,因此有必要对常规观测的半小时气象数据缺失值进行插补,同时基于该站点太阳高度角的日变化确定白天和黑夜,将观测获得的NEE数据集划分为NEEnight(夜间)和NEEdaytime(白天)两个数据子集,并且对NEE数据进行坐标轴旋转、WPL校正和储存项校正以消除地形、空气水热传输和观测高度对于NEE观测数据的影响[12]。由于夜间大气层结稳定情况下,观测CO2交换通量可能并不能反应真实的交换通量,对夜间湍流较弱情况下的数据进行U*剔除,剔除方法参考朱治林所使用的方法[25-26]。同时,对于由于天气异常、电力不稳定以及虫禽干扰等造成的错误数据也一并进行剔除。经过上述校正及剔除过程后,2003—2009年各年缺失数据统计情况见表1。
剔除后的NEEnight和NEEdaytime数据子集,有效数据量介于20%—84%之间。其中夜间有效数据的比例在20%左右(各年夜间剔除数据量明显高于白天),白天有效数据比例大约70%—80%,各年白天有效数据比例明显高于夜间(表1)。为了进一步获得完整时间序列的NEE数据,采用查表法(Look-up Table)对缺失NEE数据进行插补,因为在利用查表法进行数据插补时,不会限定NEE与环境因子的关系,从而避免了在后续的分析中产生偏移误差[19]。数据表的具体编辑如下,查表法插补NEE缺失数据[10]:将有效数据按2个月一组分为6组,再将每组内有效数据NEE按照观测高度处空气温度分为50组(-50—50℃,2℃的间隔),利用每组内有效NEE数据的平均值插补相应组别夜间缺失数据:在空气温度分组的基础上,再将每组有效NEE数据按照入射的光合有效辐射(PAR)分为20组(0—2000μmol·m-2·s-1,100μmol·m-2·s-1的间隔),利用每组有效NEE平均值插补相应组别白天缺失数据。限于缺失数据的比例比较大(表1),利用上述方法对缺失数据进行插补后可能依然存在少量数据没有被插补完整,最后采用MDV方法以7d为窗口大小对这部分数据进行再次插补,以获得完整时间序列的NEE数据集。
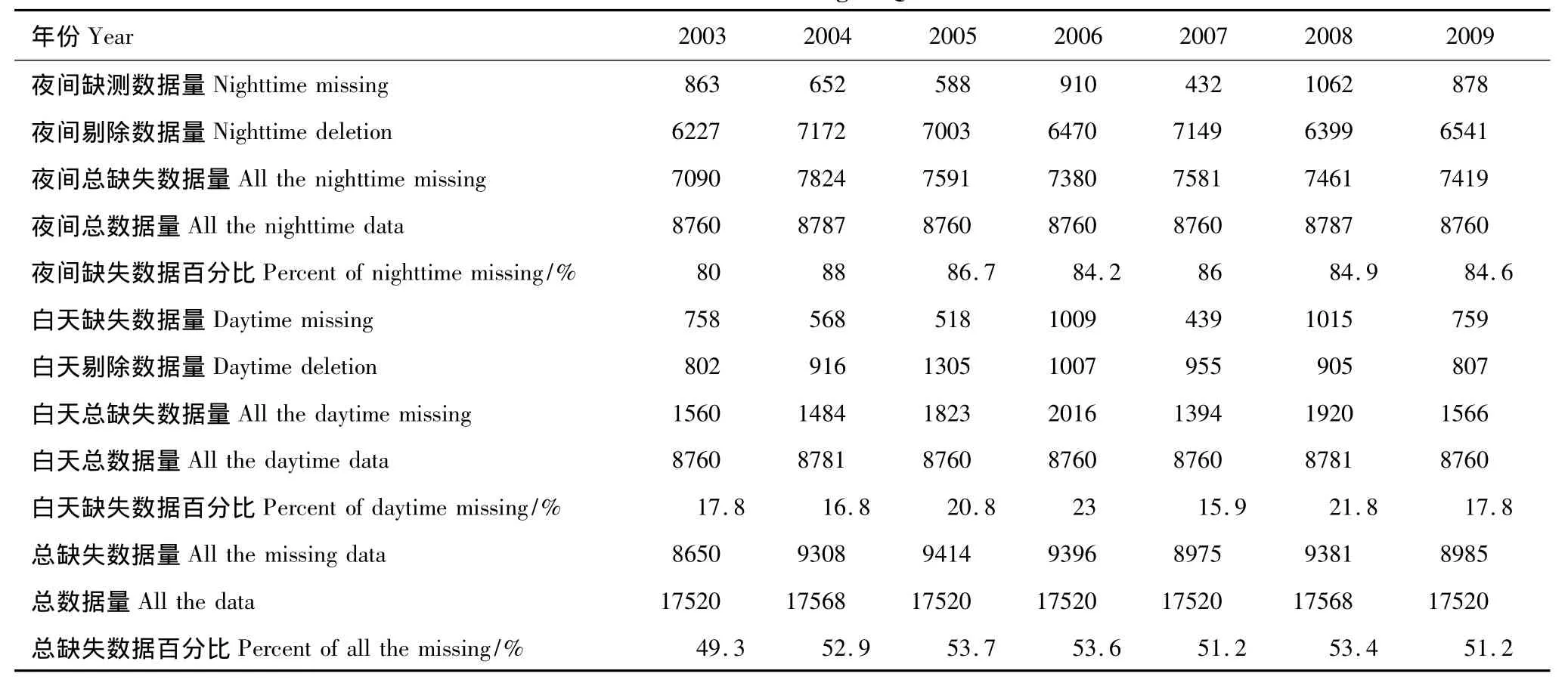
表1 2003—2009年千烟洲人工林试验站碳通量观测数据缺失情况统计Table1 Carbon flux observation data missing of QYZ station from 2003 to 2009
1.3 NEE拆分方法
采用“非线性回归模型”(Method 1)和“光响应曲线模型”(Method 2)两种方法对经过插补后的2003—2009年的半小时NEE数据进行拆分。对通量数据的谱分析表明在15d时间尺度上存在一个谱隙,说明该时间尺度通量数据受环境要素的影响较小[27],故选择15d滑动窗口。通过编写MATLAB程序进行计算,然后用ORIGIN 8.5和SPSS17.0分析作图。
1.3.1 非线性回归模型(Method 1)拆分方法
利用全年插补后的夜间NEE数据(NEEnight)和5 cm土壤温度(Ts)对非线性模型(Lloyd-Taylor方程(1))进行拟合,得到方程中相应的参数值。结合白天的土壤温度(Ts)将该方程直接外推至白天,则可估算白天半小时生态系统呼吸Re,结合已经插补完的夜间的Re(NEEnight)采用15d的滑动窗口的方法[26],拆分得到的碳通量分别表示为Re1和GPP1。

式中,Tref表示参考温度,设为283.15 K;T0为常数,设为227.13 K;ReTref和E0分别为非线性拟合的参考温度下生态系统呼吸速率(mgCO2m-2s-1)和温度敏感性参数(K);Tk为5 cm土壤温度(K)。
GPP由方程(2)计算,NEE为负表示生态系统从大气中吸收CO2。

1.3.2 光响应曲线模型(Method 2)拆分方法
采用插补后15d滑动窗口内(前后各7d)的白天半小时NEE和光合有效辐射(PAR)数据构建非线性模型(Michaelis-Menten方程,方程(3))[28-31],估算生态系统白天的平均呼吸速率,拆分得到的碳通量分别表示为Re2和GPP2。

式中,α为表观量子效率(mgCO2/μmol光子);Q为光合有效辐射(μmol光子 m-2s-1);Pmax为饱和光强下生态系统的光合速率(mgCO2m-2s-1);Reday为拟合时间段的白天的平均生态系统呼吸值(mgCO2m-2s-1)。当对不同时间段内的光响应曲线进行拟合,得到不同拟合时间段内的Re值。由此可以利用不同时间段内的Re与相应时间段内的平均温度对指数方程(1)进行拟合得到参数值,利用拟合得到的方程可对白天的Re进行估算[21]。实现对生态系统白天Re的估算,结合夜间呼吸(NEEnight)即可用方程(2)求解生态系统GPP。
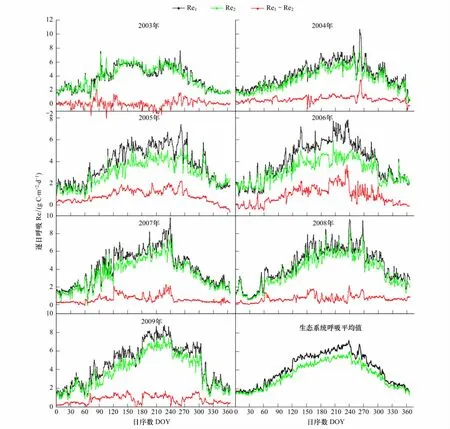
图1 逐日生态系统呼吸(Re)Fig.1 Daily ecosystem respiration(Re)partioned by different methods
2 结果与讨论
本部分将主要比较千烟洲站(QYZ)不同时间尺度两种方法拆分的生态系统碳通量(Re和GPP)异同,并分析造成这种差异可能的原因。
2.1 不同拆分方法下生态系统呼吸(Re)的估算异同
2.1.1 不同拆分方法下逐日生态系统呼吸的估算异同
从图1可以看出,不同拆分方法得到的生态系统呼吸(Re1和Re2)的季节动态变化一致:都在生长季的7月、8月份达到顶峰;2003至2007年共有7组Re1与Re2数据对应,经过双样本t检验,在5%的置信区间内,差异显著(n=365或366,P<0.01)。Re1与Re2的差值(Re1-Re2)的最大值出现时的日序数(DOY)主要落在240—290之间(2003年的第二最大值也发生在第265天),二者差值的最小值发生在温度较低的12月,1月和2月(2003年除外)。由于千烟洲人工针叶林在2003年和2007年的7月份分别发生了极度干旱和中度干旱[32],如图1所示两种拆分方法所得2003年生态系统呼吸值(Re1和Re2)在180—210天之间突然下降,随后随着8月降水量增加又继续上升;如图1所示2007年生态系统呼吸值(Re1和Re2)都在180—210d之间小幅波动以后又继续上升。2003—2004年两种不同拆分方法估算的逐日生态系统呼吸差值(Re1-Re2)的平均值小于1 gC·m-2·d-1,介于0.07—0.87 gC·m-2·d-1之间。全年逐日生态系统呼吸值中,Re1大于Re2的占多数,介于58%—100%之间。2003—2009年全年生态系统平均逐日Re1介于0.45—7.28 gC·m-2·d-1,平均逐日Re2介于0.67—6.15 gC·m-2·d-1(图1),Re1-Re2结果见表2。
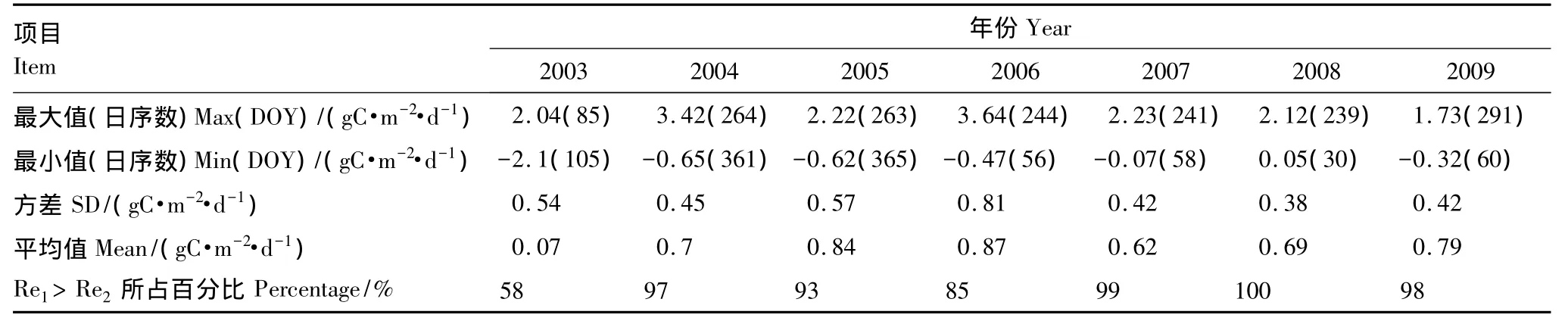
表2 (Re1-Re2)值的逐日统计分析Table2 Statistical analysis of daily(Re1-Re2)
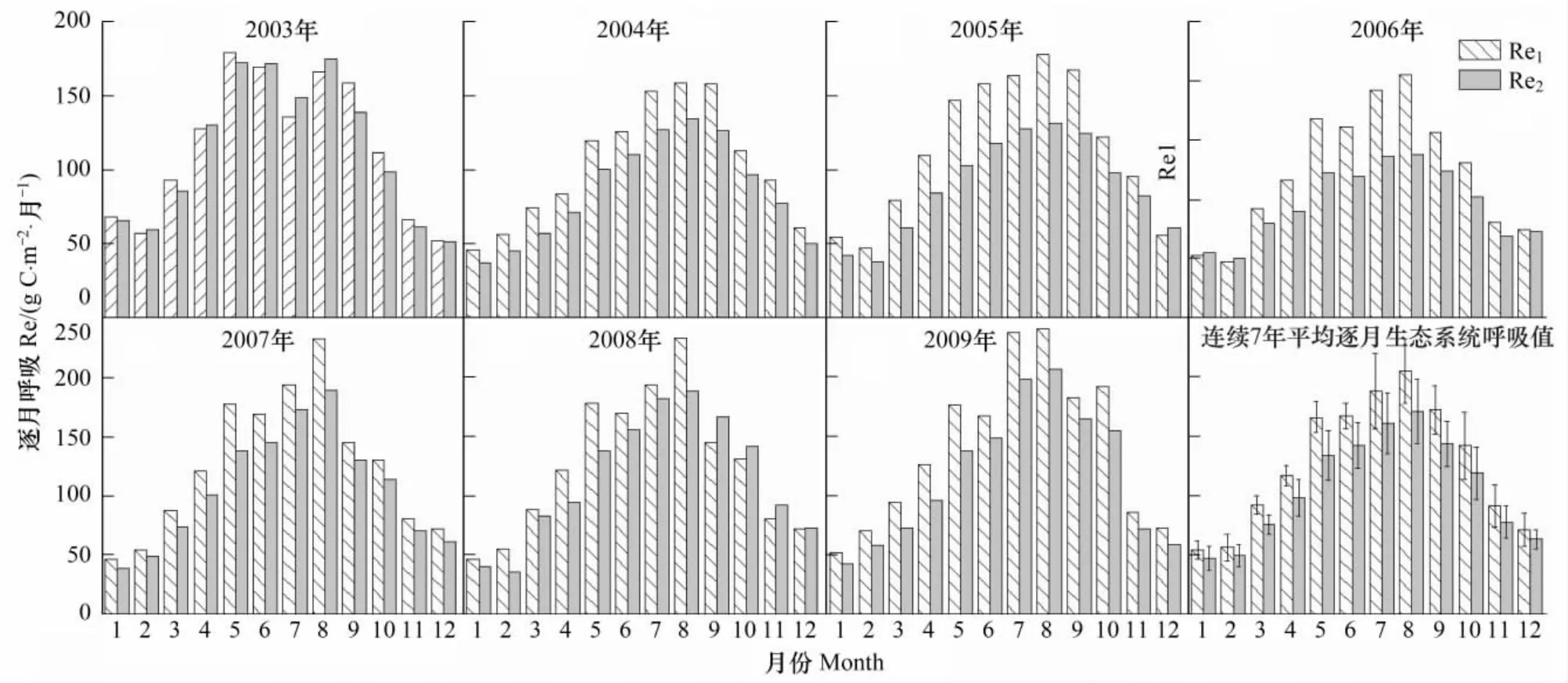
图2 2003—2009年逐月生态系统呼吸Fig.2 Monthly ecosystem respiration from 2003 to 2009
2.1.2 不同拆分方法下逐月生态系统呼吸估算异同
如图2所示,两种拆分方法估算的逐月生态系统呼吸(Re1和Re2)的季节变化动态表现一致,除了2003年以外都在8月份达到最大值,2003年因为发生极端干旱,8月份生态系统呼吸较7月份显著降低。图2h所表示的两种拆分方法所得的平均逐月生态系统呼吸值也表现出相同的季节动态特征,从非生长季开始逐渐升高在8月份达到峰值;Re1和Re2的平均逐月生态系统呼吸值的标准差第一最大值和第二最大值都发生在7、8月份,两种方法都体现了生长季的7、8月份生态系统呼吸的年际波动变化最大。连续7年逐月Re1与逐月Re2之间差值(Re1-Re2)的最大值主要集中在生长季的8、9月份(2008年出现在6月份),最小值主要发生在冬季(除2003年发生在7月份),且有些年份(2003、2005和2006)的冬季出现了逐月Re1小于逐月Re2,而其他年份的所有逐月Re1都大于Re2。平均逐月Re1范围在53.7—204.7gC·m-2·mon-1之间,平均逐月Re2介于47.1—170.9 gC·m-2·mon-1;Re1和Re2分别在7月和8月标准差最大。
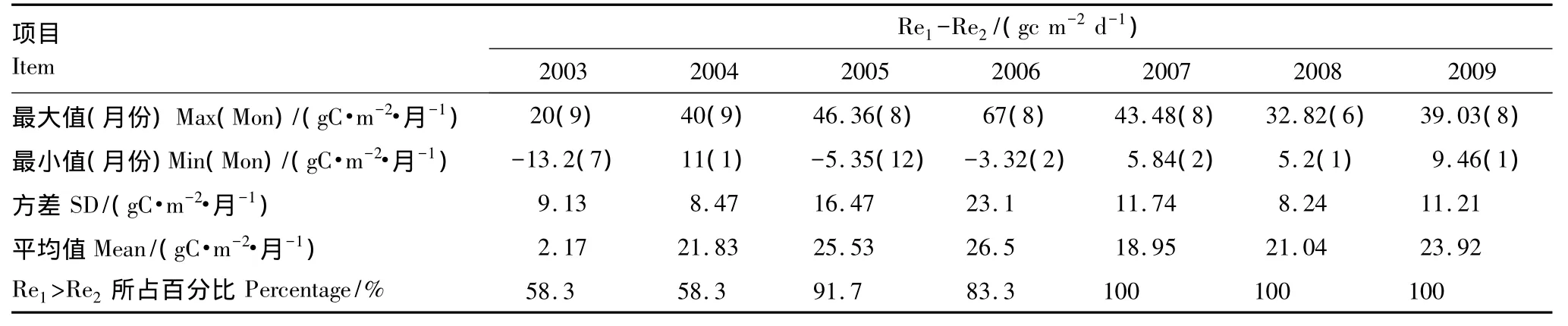
表3 2003—2009年(Re1-Re2)值的逐月统计分析Table3 Statistical analysis of monthly(Re1-Re2)
2.1.3 不同拆分方法下逐年生态系统呼吸估算异同
如图3所示,不同年份的逐年 Re1均高于逐年Re2,且2006年差值幅度最大(317.6 gC·m-2·a-1),2003年高出部分最少(30 gC·m-2·a-1),2003—2009年不同拆分方法下逐年 Re1相对于逐年 Re2高出2%—28.6%,且2005年Re1高于Re2的百分比最大。除了2005年生态系统呼吸下降以外,两种方法都表现出千烟洲人工针叶林的逐年生态系统呼吸是递增的。
由于Method 1和Method 2都采用了查表法来插补全天NEE缺失数据,夜间NEE数据即为晚间生态系统呼吸(Renight),区别在于Method 1基于夜间呼吸数据来对Lloyd-Taylor方程进行拟合估算出白天生态系统呼吸,而Method 2基于白天呼吸数据对Lloyd-Taylor方程进行拟合估算白天生态系统呼吸。从图4可以看出,千烟洲人工针叶林生态系统基于夜间(Method 1)与白天数据(Method 2)得到的夜间与白天生态系统呼吸对温度的响应可以看出,夜间的Re对温度的响应与白天的Re对温度的响应存在差异,利用夜间呼吸 (Method 1)与土壤表层温度拟合的Lloyd-Taylor方程对白天的Re估算时,会产生显著的高估现象。在千烟洲人工针叶林基于夜间数据得到的参考呼吸ReTref要大于基于白天数据的结果(表4),证实了在千烟洲人工针叶林生态系统中植物叶片呼吸收到Kok效应的影响,即光对植物叶片暗呼吸的抑制作用[16,21,33],致使植物叶片呼吸在白天要低于夜间,因此2003—2009年的逐年生态系统呼吸Re1会高出Re2(图3)。
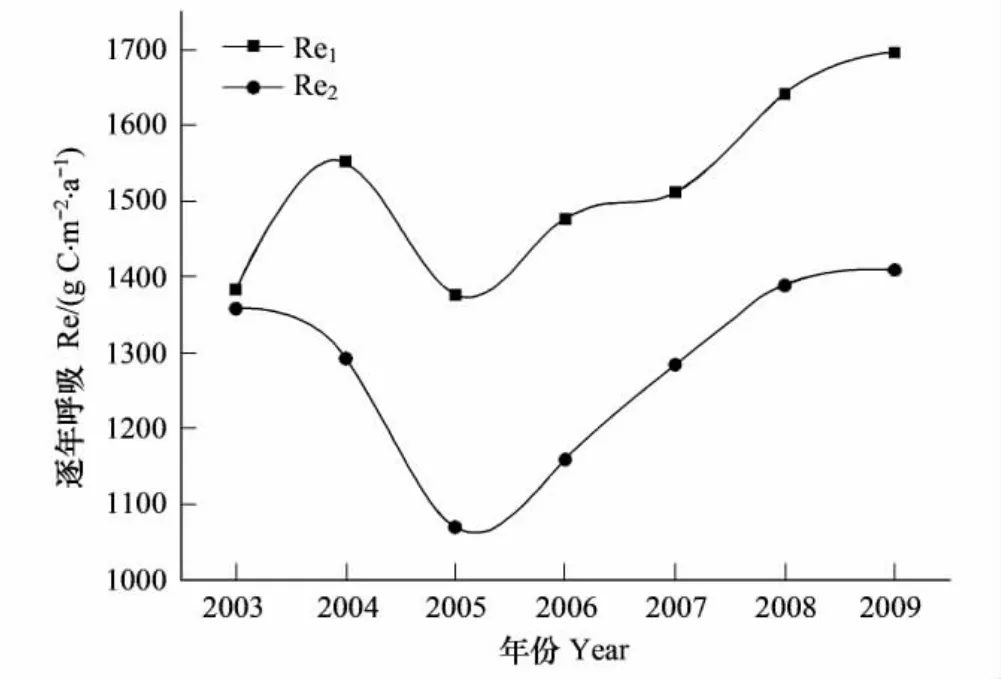
图3 2003—2009年逐年生态系统呼吸Fig.3 Annual ecosystem respiration from 2003 to 2009
2.2 不同拆分方法下生态系统总初级生产力(GPP)的估算异同
由于使用相同的方法来插补缺失的NEE数据,故两种拆分方法得到的不同时间尺度生态系统呼吸之差(Re1-Re2)和总初级生产力GPP之差(GPP1-GPP2)是相等的,这里就不再讨论GPP1-GPP2。两种拆分方法得到的千烟洲人工针叶林生态系统总初级生产力(GPP1和GPP2)的季节动态变化一致,均在每年的7,8月份生长季达到最大值(图5,图6)。千烟洲人工针叶林生态系统2003和2007年夏季发生了伏旱,两种拆分方法估算的生态系统总初级生产力(GPP1和GPP2)在7月份开始下降之后随着降水的增加而上升,最终在进入非生长季后又逐渐降低(图5,图6)。对逐日、逐月GPP1和GPP2进行两独立样本t检验,差异显著(P<0.01)。2003—2009年全年生态系统平均逐日GPP1介于1.3—8.1 gC·m-2·d-1,平均逐日GPP2介于1.2—7.3 gC·m-2·d-1(图5)。GPP1和GPP2分别在7月份和6月份达到标准差最大值,平均值的范围分别是58.3—224 gC·m-2·月-1和51.8—195.8 gC·m-2·月-1(图6)。
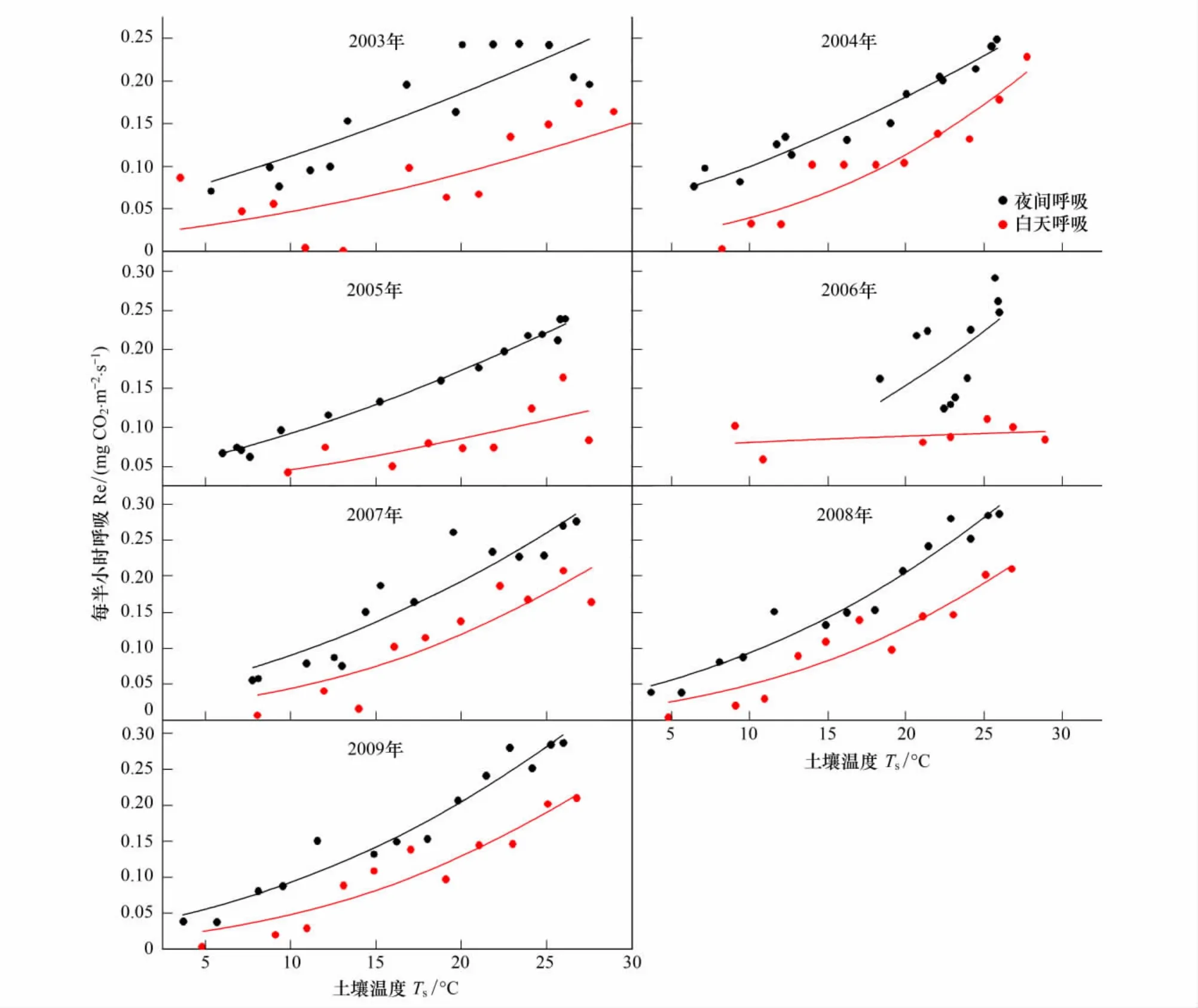
图4 2003—2009年千烟洲亚热带人工针叶林夜间及白天生态系统呼吸(Re)对土壤温度T s(5 cm)的响应Fig.4 Response of nighttime and daytime Re in QYZ plantation to soil temperature at 5cm from 2003 to 2009
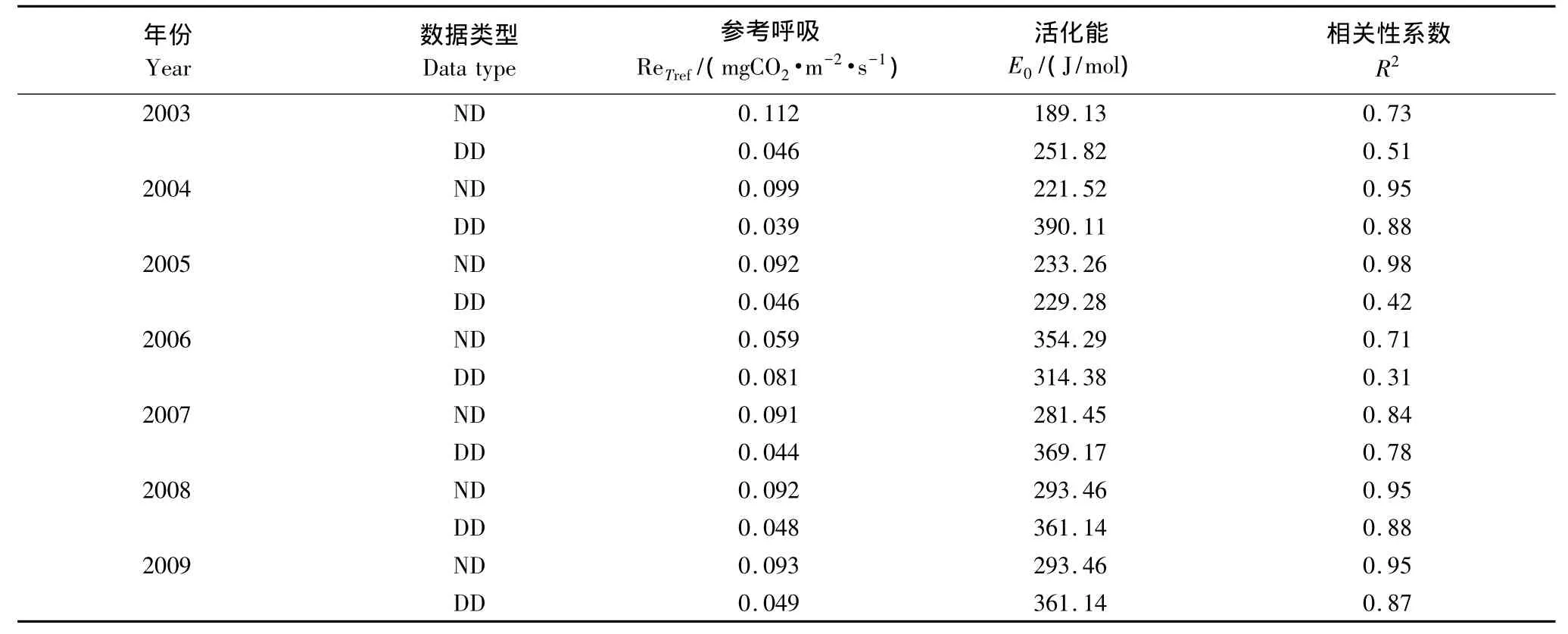
表4 千烟洲针叶林(QYZ)基于夜间数据(ND)及白天数据(DD)对Lloyd-Taylor方程拟合得到方程参数Table4 Parameters of Lloyd-Taylor equation based on nighttime data(ND)and daytime data(DD)at QYZ
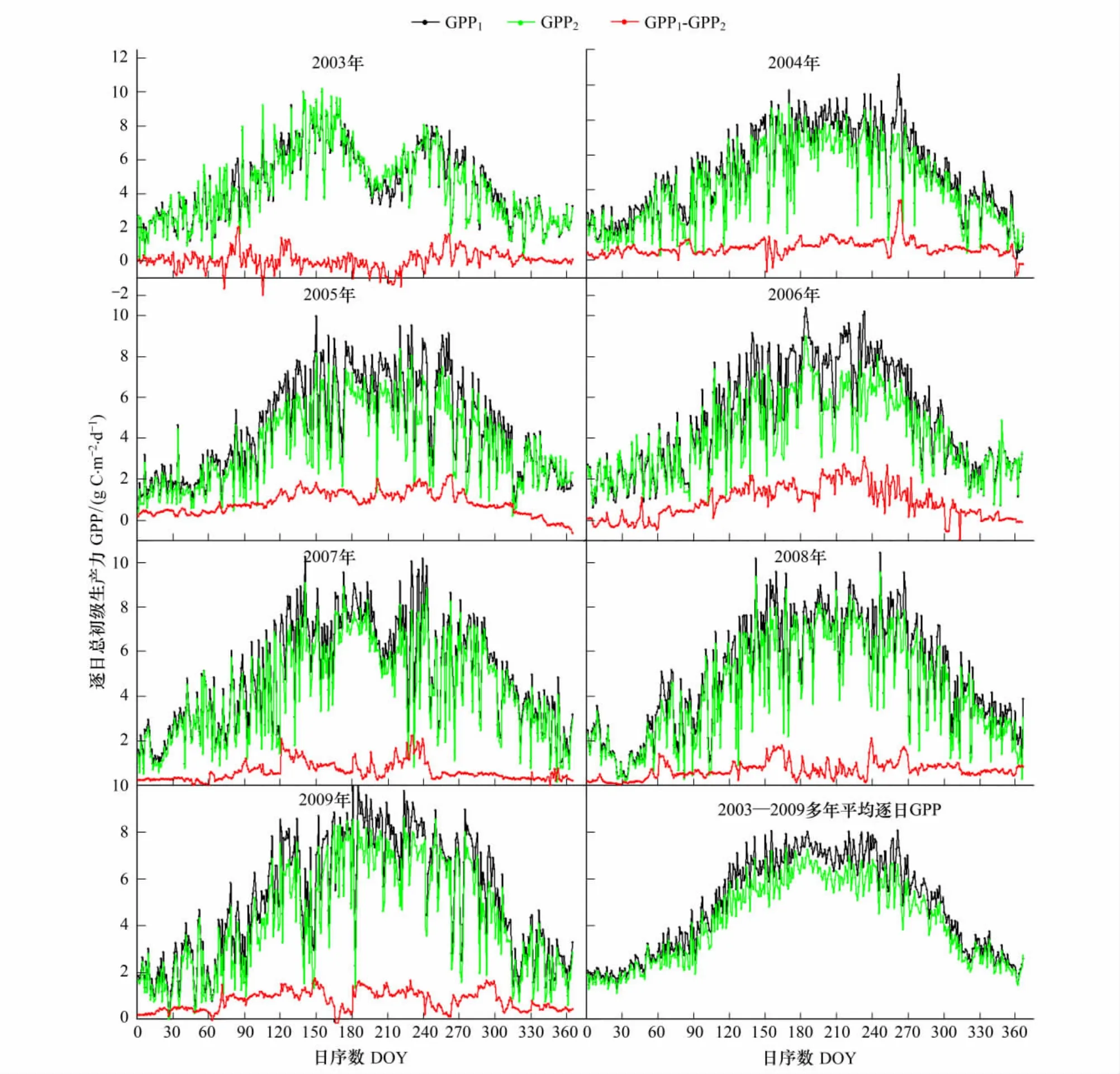
图5 2003—2009年千烟洲针叶林生态系统逐日GPPFig.5 Daily ecosystem Gross Primary Productivity(GPP)at QYZ from 2003 to 2009
如图7所示,2003—2009年的逐年生态系统总初级生产力,GPP1分别高出GPP2为26(1.6%)、262(17%)、306.3(23%)、317.6(21.3%)、227.4(14.5%)、252.5(16.4)和287 gC·m-2·a-1(18.5%),这与3.1中Re1高于Re2是一致的。
2.3 不同拆分方法下生态系统碳通量组分(Re和GPP)之差及其环境响应机制
由于不同时间尺度的Re1-Re2值与GPP1-GPP2值相同,这里只讨论生态系统呼吸差值的平均值((Re1-Re2)_avg)变化(图8)。2003—2009年,千烟洲人工针叶林生态系统的不同方法拆分估算的平均呼吸之差(Re1-Re2)的最大值于最小值分别发生在8月份(33.8 gC·m-2·月-1)和2月份(6.3 gC·m-2·月-1);2006年逐年生态系统呼吸值之差(Re1-Re2)达到最大值(317.6 gC·m-2·a-1),其次为2005年(306.3 gC·m-2·a-1),最小值发生在2003年(30 gC·m-2·a-1)。本文3.1中指出逐日和逐月的两种方法拆分所得呼吸的最小值都发生冬季(1,2,12月),同时最小呼吸差值也主要发生在冬季,这可能是因为冬季低温对呼吸的限制起主要作用,所以差值并不明显[34]。
从图9中可以看出,不同拆分方法下所得到的千烟洲人工针叶林生态系统呼吸和总初级生产力的差值(Re1-Re2)在不同时间尺度上受不同环境因子的影响限制。在日尺度上,影响因子依次为饱和水汽压差(R2=0.63)、光合有效辐射(R2=0.6);在月尺度上,影响因子依次为光合有效辐射(R2=0.89)、饱和水汽压差(R2=0.85)和降水量(R2=0.48);在年尺度上,影响因子为光合有效辐射(R2=0.78)。PAR是影响千烟洲中亚热带人工针叶林生态系统植被生长季碳吸收能力的主要因素之一[35-37],每年的生长季7—9月份光合有效辐射(PAR)和饱和水汽压差(VPD)会达到峰值,呼吸差平均值主要出现在8—9月份(图2,图6,图8)。由于2006年的光合有效辐射(PAR)最高,所以差异值最大,随着光合有效辐射和空气饱和水汽压的逐渐升高,植物叶片呼吸受到的Kok效应的影响越来越大,差值不断升高;达到一个峰值以后,Kok效应又逐渐减弱,呼吸差值又逐渐下降。当逐月光合有效辐射(PAR)接近905 mol·m-2·月-1,空气饱和水汽压差(VPD)接近1.18 kPa时,两种方法得到的逐月呼吸差值最大;当全年总的光合有效辐射(PAR)接近6323 mol·m-2·a-1时,两种拆分方法所得到的生态系统碳通量组分(GPP和Re)差别最大(图9)。
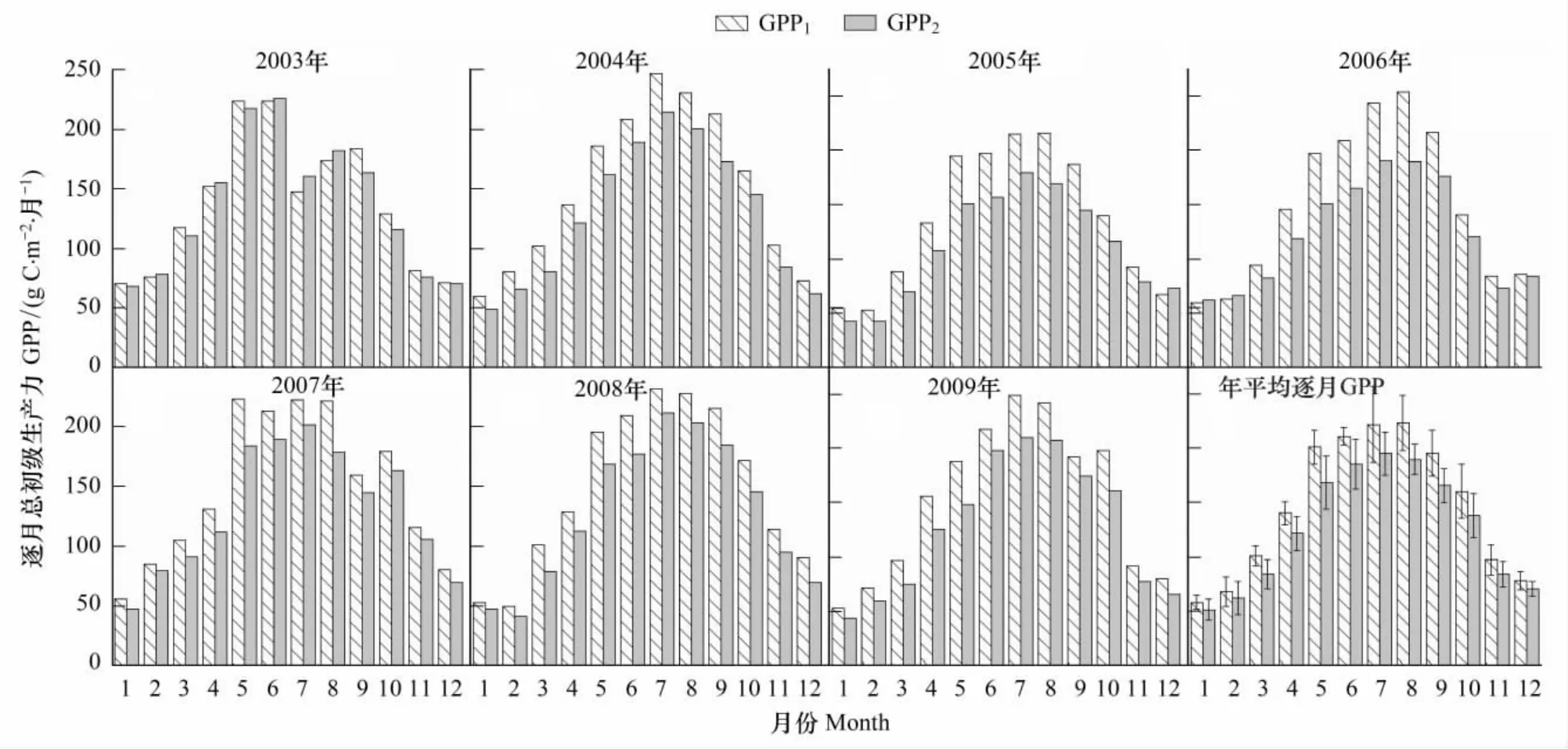
图6 2003—2009年千烟洲(QYZ)人工针叶林生态系统逐月GPPFig.6 Monthly ecosystem GPP at QYZ from 2003 to 2009

图7 2003—2009年千烟洲(QYZ)人工针叶林生态系统逐年GPPFig.7 Annual ecosystem GPP at QYZ from 2003 to 2009
3 结论
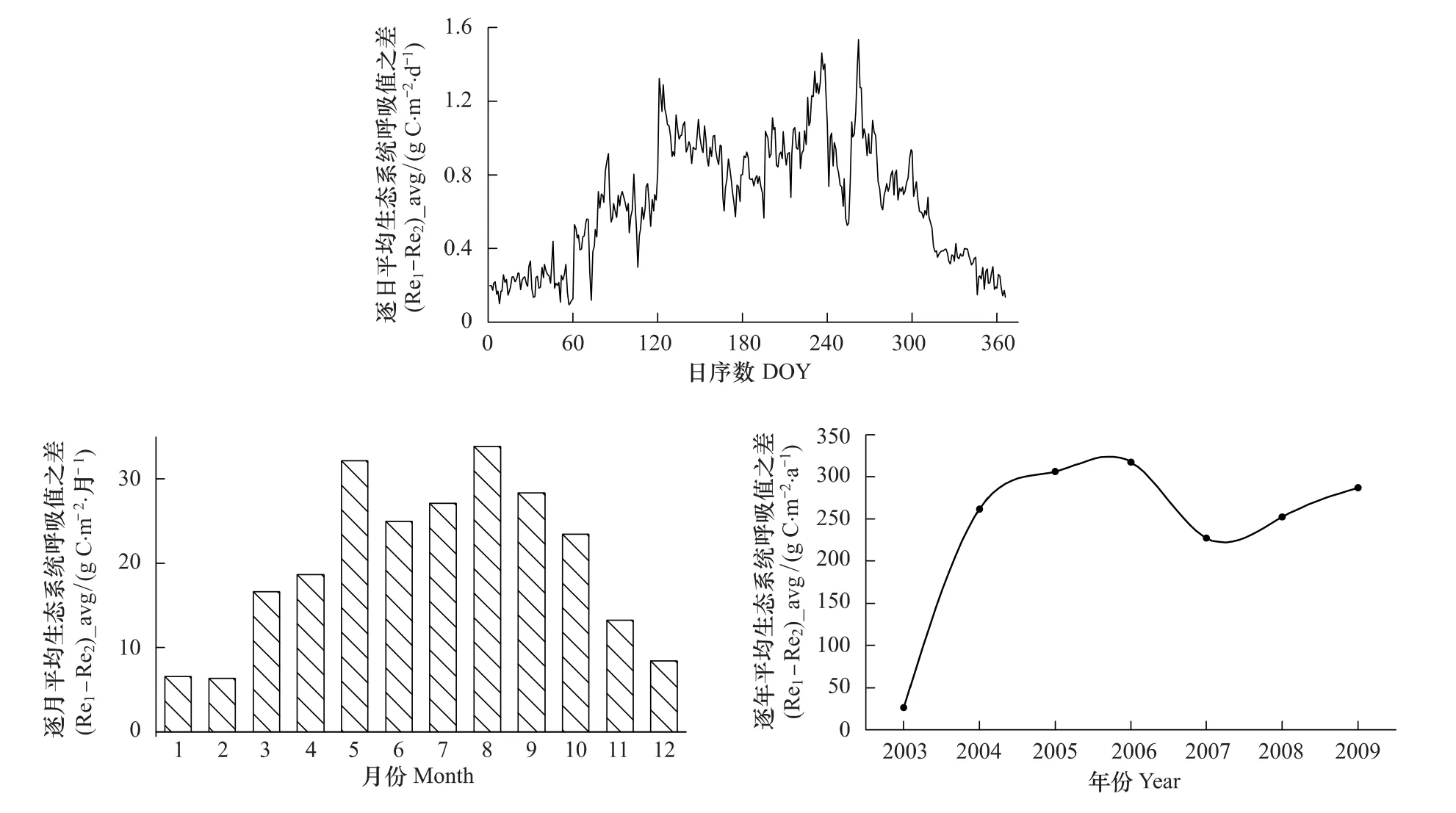
图8 2003—2009年千烟洲人工针叶林两种拆分方法下平均生态系统呼吸值差时间序列图Fig.8 Time series of average(Re1-Re2)at QYZ from 2003 to 2009
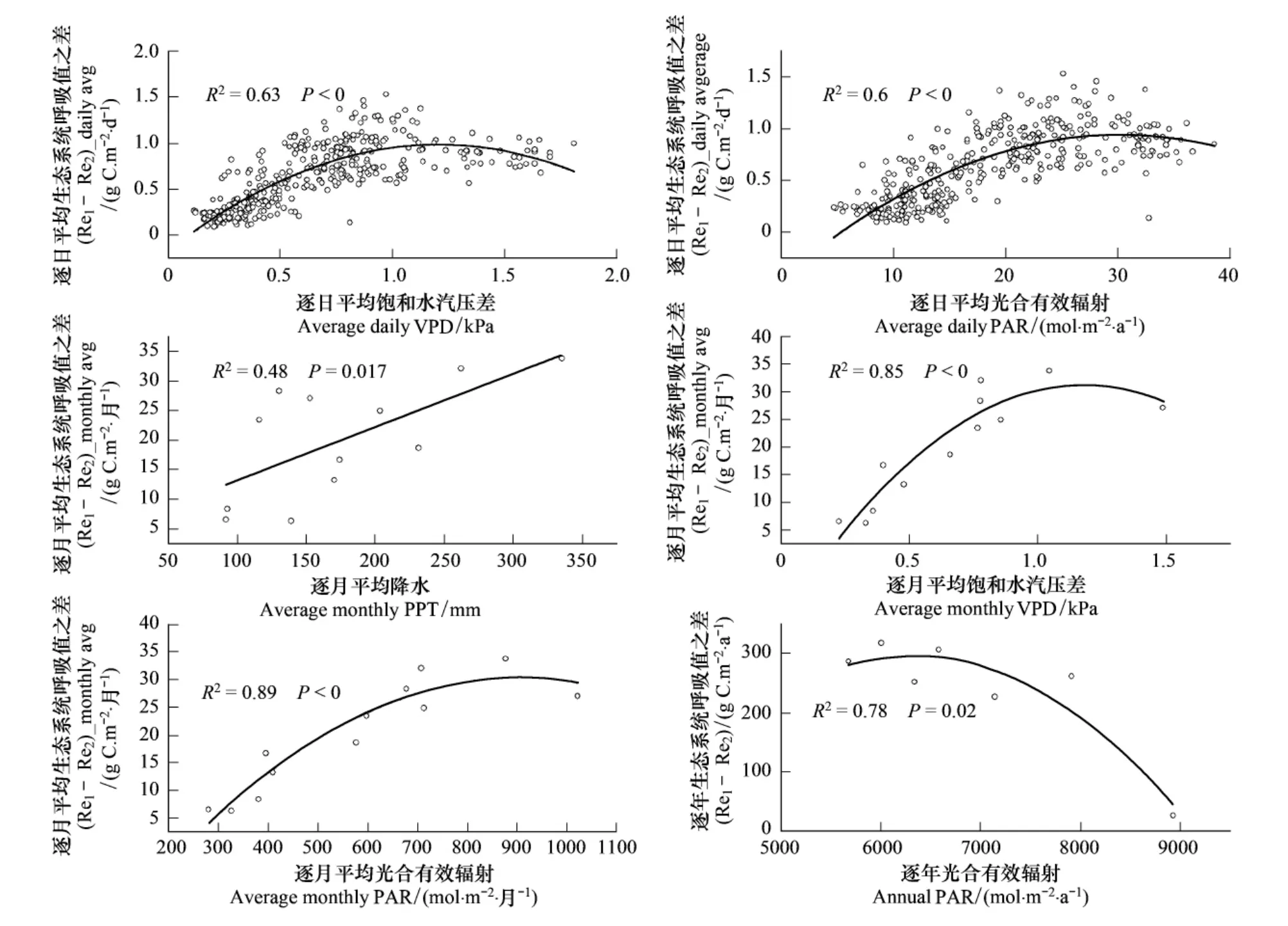
图9 千烟洲人工针叶林不同拆分方法生态系统呼吸平均差值(Re1-Re2)_avg的环境响应机制Fig.9 Environmental response mechanism of(Re1-Re2)_avg at QYZ
本研究基于2003—2009年的千烟洲人工针叶林生态系统的涡度通量观测数据和气象数据分析了两种不同方法在拆分中亚热带人工针叶林生态系统碳通量的差异及其环境响应机制,结果表明:(1)两种不同拆分方法得到的生态系统碳通量组分(GPP和Re)季节变化动态一致,在生长季达到峰值;2003和2007年伏旱的影响使得Re1和Re2、GPP1和GPP2均在8月份出现了下降随后又逐渐升高。(2)两种拆分方法所得生态系统呼吸差值(Re1-Re2)的最大值发生在生长季的8,9月份,而最小值主要发生在冬季;逐年Re1相比于比Re2高出2%—28.6%,逐年GPP1相比于比GPP2高出1.6%—23%;最大值发生在2006年(317.6gC·m-2·a-1),最小值发生2003年(30 gC·m-2·a-1)。(3)利用晚间数据(Method 1)对Lloyd-Taylor方程拟合得到的参数显著高于基于白天呼吸数据(Method 2)拟合该方程得到的参数,Kok效应确实抑制植物白天叶片的呼吸使得利用非线性回归模型在拆分NEE数据时存在高估生态系统呼吸和总初级生产力(GPP和Re)。(4)两种不同拆分方法得到的对应碳通量组分差值(Re1-Re2)在不同时间尺度上受不同环境因子影响:在日尺度上,影响因子依次为空气饱和水汽压差R2=0.63)和光合有效辐射R2=0.6);在月尺度上,影响因子依次为光合有效辐射(R2=0.89)、饱和水汽压差(R2=0.85)和降水量(R2=0.48);在年尺度上,主要影响因子为光合有效辐射(R2=0.78)。月光合有效辐射最能解释“Kok效应”对叶片白天呼吸的抑制。
不同拆分方法下千烟洲人工针叶林生态系统呼吸差值(Re1-Re2)季节差异性明显(8,9月份达到峰值),且月尺度上环境因子最能解释差异性。在利用非线性回归的方法(Method 1)处理千烟洲人工针叶林生态系统全年涡度NEE观测数据时,如果当月总光合有效辐射(PAR)接近905 mol·m-2·月-1、空气饱和水气压差接近1.18 kPa时,此时由于温度升高降水增多,“Kok效应”对呼吸的抑制作用最为强烈,应该考虑使用光响应曲线模型(Method 2)来拆分该月涡度NEE观测数据,两种方法结合使用以减小全年NEE的拆分误差。
由于碳通量的拆分方法与摩擦风速、NEE有效数据量、气象数据等密切相关,不同时间尺度上呼吸差值与环境因子响应程度的大小也对计算方法也有一定依赖性,而且不同生态系统的气候条件和碳通量季节变化趋势也有很大不同,本研究只分析了两种方法在中亚热带人工针叶林生态系统的应用,为了进一步分析和了解碳通量拆分结果的差异和不确定性,还需要将这3种方法在中国其他不同的生态系统中加以深入分析和讨论,从而为确定适合中国通量网的碳通量拆分方法提供比较和参考。
[1] Sulkava M,Luyssaert S,Zaehle S,Papale D.Assessing and improving the representativeness of monitoring networks:the European flux tower network example.Journal of Geophysical Research,2011,116:G00J04,doi:10.1029/2010JG001562.
[2] Baldocchi D.Breathing of the terrestrial biosphere:lessons learned from a global network of carbon dioxide flux measurement systems.Australian Journal of Botany,2008,56(1):1-26.
[3] Reichstein M,Falge E,Baldocchi D,Aubinet M,Berbigier P,Bernhofer C,Buchmann N,Gilmanov T,Granier A,Grünwald T,HavránkováK,Ilvesniemi H,Janous D,Knohl A,Laurila T,Lohila A,Loustau D,Matteucci G,Meyers T,Miglietta F,Ourcival JM,Pumpanen J,Rambal S,Rotenberg E,Sanz M,Tenhunen J,Seufert G,Vaccari F,Vesala T,Yakir D,Valentini R.On the separation of net ecosystem exchange into assimilation and ecosystem respiration:review and improved algorithm.Global Change Biology,2005,11(9):1424-1439.
[4] Desai A R,Richardson A D,Moffat A M,Kattge J,Hollinger D Y,Barr A,Falge E,Noormets A,Papale D,Reichstein M,Stauch V.Cross-site evaluation of eddy covariance GPP and RE decomposition techniques.Agricultural and Forest Meteorology,2008,148(6/7):821-838.
[5] Bowling D R,Tans P P,Monson R K.Partitioning net ecosystem carbon exchange with isotopic fluxes of CO2.Global Change Biology,2001,7(2):127-145.
[6] Zobitz J M,Burns S P,Ogee J,Reichstein M,Bowling R.Partitioning net ecosystem exchange of CO2:a comparison of a Bayesian/isotope approach to environmental regression methods.Journal of Geophysical Research,2007,112:G03013,doi:10.1029/2006JG000282.
[7] Griffis T J,Black T A,Gaumont-Guay D,Drewitt G B,Nesic Z,Barr A G,Morgenstern K,Kljun N.Seasonal variation and partitioning of ecosystem respiration in a southern boreal aspen forest.Agricultural and Forest Meteorology,2004,125(3/4):207-223.
[8] Law B E,Falge E,Gu L,Baldocchi D D,Bakwin P,Berbigier P,Davis K,Dolman A J,Falk M,Fuentes JD,Goldstein A,Granier A,Grelle A,Hollinger D Y,Janssens IA,Jarvis P,Jensen NO,Katul G,Mahli Y,Matteucci G,Meyers T,Monson R,Munger W,Oechel W,Olson R,Pilegaard K,Paw K T,Thorgeirsson H,Valentini R,Verma S,Vesala T,Wilson K,Wofsy S.Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation.Agricultural and Forest Meteorology,2002,113(1/4):97-120.
[9] Unger S,Maguas C,Pereira JS,Aires L M,David T S,Werner C.Partition carbon fluxes in a Mediterranean oak forest to disentangle change in ecosystem sink strength during drought.Agricultural and Forest Meteorology,2009,149(6/7):949-961.
[10] Falge E,Baldocchi D,Olson R,Anthoni P,Aubinet M,Bernhofer C,Burba G,Ceulemans R,Clement R,Dolman H,Granier A,Gross P,Grunwald T,Hollinger D,Jensen N O,Katul G,Keronen P,Kowalski A,Lai CT,Law B E,Meyers T,Moncrieff H,Moors E,Munger JW,Pilegaard K,Rannik U,Rebmann C,Suyker A,Tenhunen J,Tu K,Verma S,Vesala T,Wilson K,Wofsy S.Gap filling strategies for defensible annual sums of net ecosystem exchange.Agricultural and Forest Meteorology,2001,107(1):43-69.
[11] Moffat A M,Papale D,Reichstein M,Hollinger D Y,Richardson A D,Barr A G,Beckstein C,Braswell B H,Churkina G,Desai A R,Falge E,Gove J H,Heimann M,Hui D F,Jarvis A J,Kattge J,Noormets A,Stauch,V J.Comprehensive comparison of gap-filling techniques for eddy covariance net carbon fluxes.Agricultural and Forest Meteorology,2007,147(3/4):209-232.
[12] Fu Y L,Yu G R,Sun X M,Li Y N,Wen X F,Zhang L M,Li Z Q,Zhao L,Hao Y B.Depression of net ecosystem CO2exchange in semi-arid Leymus chinensis steppe and alpine shrub.Agricultural and Forest Meteorology,2006,137(3/4):234-244.
[13] Suyker A E,Verma SB.Year-round observations of the net ecosystem exchange of carbon dioxide in a native tallgrass prairie.Global Change Biology,2001,7(3):279-289.
[14] Griffis T J,Black T A,Morgenstern K,Barr A G,Nesic Z,Drewitt G B,Gaumont-Guay D,McCaughey J H.Ecophysiological controls on the carbon balances of three southern boreal forests.Agricultural and Forest Meteorology,2003,117(1/2):53-71.
[15] Gilmanov T G,Soussana J E,Aires L,Allard V,Ammann C,Balzarolo M,Barcza Z,Bernhofer C,Campbell C L,Cernusca A,Cescatti A,Clifton-Brown J,Dirks B O M,Dore S,Eugster W,Fuhrer J,Gimeno C,Gruenwald T,Haszpra L,Hensen A,Ibrom A,Jacobs AFG,Jones MB,Lanigan G,Laurila T,Lohila A,Manca G,Marcolla B,Nagy Z,Pilegaard K,Pinter K,Pio C,Raschi A,Rogiers N,Sanz MJ,Stefani P,Sutton M,Tuba Z,Valentini R,Williams M L,Wohlfahrt G.Partitioning European grassland net ecosystem CO2exchange into gross primary productivity and ecosystem respiration using light response function analysis.Agriculture Ecosystems and Environment,2007,121(1/2):93-120.
[16] Janssens I A,Lankreijer H,Matteucci G,Kowalski A S,Buchmann N,Epron D,Pilegaard K,Kutsch W,Longdoz B,Grunwald T,Montagnani L,Dore S,Rebmann C,Moors E J,Grelle A,Rannik U,Morgenstern K,Oltchev S,Clement R,Gudmundsson J,Minerbi S,Berbigier P,Ibrom A,Moncrieff J,Aubinet M,Bernhofer C,Jensen N O,Vesala T,Granier A,Schulze E D,Lindroth A,Dolman A J,Jarvis P G,Ceulemans R,Valentini R.Productivity overshadows temperature in determining soil and ecosystem respiration across European forests.Global Change Biology,2001,7(3):269-278.
[17] Sharp R E,Matthews M A,Boyer J S.Kok Effect and the quantum yield of photosynthesis-light partially inhibits dark respiration.Plant Physiology,1984,75(1):95-101.
[18] Tjoelker M G,Oleksyn J,Reich P B.Modelling respiration of vegetation:evidence for a general temperature-dependent Q10.Global Change Biology,2001,7(2):223-230.
[19] Baldocchi D,Falge E,Gu L H,Olson R,Hollinger D,Running S,Anthoni P,Bernhofer C,Davis K,Evans R,Fuentes J,Goldstein A,Katul G,Law B,Lee X H,Malhi Y,Meyers T,Munger W,Oechel W,Paw K T,Pilegaard K,Schmid H P,Valentini R,Verma S,Vesala T,Wilson K,Wofsy S.FLUXNET:a new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide,water vapor,and energy flux densities.Bulletin of the American Meteorological Society,2001,82(11):2415-2434.
[20] Falge E,Baldocchi D,Tenhunen J,Aubinet M,Bakwin P,Berbigier P,Bernhofer C,Burba G,Clement R,Davis K J,Elbers JA,Goldstein A H,Grelle A,Granier A,Guomundsson J,Hollinger D,Kowalski A S,Katul G,Law B E,Malhi Y,Meyers T,Monson R K,Munger J W,Oechel W,Paw K T,Pilegaard K,Rannik U,Rebmann C,Suyker A,Valentini R,Wilson K,Wofsy S.Seasonality of ecosystem respiration and gross primary production as derived from FLUXNET measurements.Agricultural and Forest Meteorology,2002,113(1/4):53-74.
[21] Stoy P C,Katul GG,Siqueira M B S,Juang JY,Novick K A,Uebelherr JM,Oren R.An evaluation of models for partitioning eddy covariancemeasured net ecosystem exchange into photosynthesis and respiration.Agricultural and Forest Meteorology,2006,141(1):2-18.
[22] Yu G R,Wen X F,Sun X M,Tanner B D,Lee X H,Chen JY.Overview of China FLUX and evaluation of its eddy covariance measurement.Agricultural and Forest Meteorology,2006,137(3/4):125-137.
[23] Liu Y F,Yu G R,Wen X F,Wang Y H,Wang Y H,Song X,Li J,Sun X M,Yang F T,Chen Y R,Liu Q J.Seasonal dynamics of CO2fluxes from subtropical plantation coniferous ecosystem.Science in China,Series D,2006,36(Supp I):91-102.
[24] Mi N,Yu G E,Wen X F,Sun X M,Wang SS.Responses of subtropical conifer plantation to future climate change:a simulation study.Chinese Journal of Applied Ecology,2008,19(9):1877-1883.
[25] Zhu Z L,Sun X M,Wen X F,Zhou Y L,Tian J,Yuan G F.Study on the processing method of nighttime CO2 eddy covariance flux data in ChinaFLUX.Science in China,Series D,2006,36(S1):34-44.
[26] Sun XM,Wen X F,Yu G R,Liu Y F,Liu Q J.Seasonal drought effects on carbon sequestration of a mid-subtropical planted forest of southern China.Science in China,Series D,2006,36(Supp I):103-110.
[27] Baldocchi D,Falge E,Wilson K.A spectral analysis of biosphere-atmosphere traces gas flux densities and meteorological variables across hour to multi-year time scales.Agricultural and Forest Meteorology,2001,107(1):1-27.
[28] Zhang L M.Ecophysiological controls on seasonal variations of ecosystem carbon exchange of typical Ecosystems along NSTEC [D].Beijing:2006.
[29] Goulden M L,Munger JW,Fan SM,Sutton DJ,Bazzaz A,Munger JW,Wofsy SC.Physiological responsesof a black spruce forest toweather.Journal of Geophysical Research,1997,102(24):28987-28996.
[30] Aubinet M,Chermance B,Vabderhaute M,Longdoz B,Yernaux M,Laitat E.Long term carbon dioxide exchange above a mixed forest in the Belgian Ardennes.Agricultural and Forest Meteorology,2001,108(4):293-315.
[31] Law B E,Falge E,Gu L H,Baldocchi D D,Bakwin P,Berbigier P,Davis K,Dolman A J,Falk M,Fuentes J D,Goldstein A,Granier A,Grelle A,Hollinger D,Janssens IA,Jarvis P,Jensen N O,Katul G,Mahli Y,Matteucci G,Meyers T,Monson R,Munger W,Oechel W,Olson R,Pilegaard K,Paw K T,Thorgeirsson H,Valentini R,Verma S,Vesala T,Wilson K,Wofsy S.Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation.Agricultural and Forest Meteorology,2002,113(1/4):97-120.
[32] Wen X F,Wang H M,Wang J L,Yu G R,Sun X M.Ecosystem carbon exchanges of a subtropical evergreen coniferous plantation subjected to seasonal drought,2003—2007.Biogeosciences,2010,7(1):357-369.
[33] Brooks A,Farquhar G D.Effect of Temperature on the CO2/O2specificity of ribulose-1,5-bisphosphate carboxylase/oxygenase and the rate of respiration in the light.Planta,1985,165(3):397-406.
[34] Zhang W J,Wang H M,Yang F T,Yi Y H,Wen X F,Sun X M,Yu G R,Wang Y D,Ning J C.Underestimated effects of low temperature during early growing season on carbon sequestration of a subtropical coniferous plantation.Biogeosciences,2011,8(6):1667-1678.
[35] Liu Y F,Song X,Sun X M,Wen X F,Chen Y R.CO2flux seasonal variations of coniferous forest in Qianyanzhou and its affection on environmental factors.Science in China,Series D:Earth Sciences,2004,34(Supp Ⅱ):109-117.
[36] Yu G R,Wen X F,Li QK,Zhang L M,Ren CY,Liu Y F,Guan D X.Seasonal patterns and environmental control of ecosystem respiration in subtropical and temperate forests in China.2004,Science in China,Series D:Earth Sciences,2004,34(Supp Ⅱ):84-94.
[37] Gu X F,Yu GR,Wen X F,Tao B,Li K R,Liu Y F.Drought effects on carbon exchange in a subtropical coniferous plantation in China.Journal of Plant Ecology,2008,32(5):1041-1051.
参考文献:
[23] 刘允芬,于贵瑞,温学发,王迎红,宋霞,李菊,孙晓敏,杨风亭,陈永瑞,刘琪璟.千烟洲中亚热带人工林生态系统CO2通量的季节变异特征.中国科学(D辑),2006,36(增刊I):91-102.
[24] 米娜,于贵瑞,温学发,孙晓敏,王树森.中亚热带人工针叶林对未来气候变化的响应.应用生态学报,2008,19(9):1877-1883.
[25] 朱治林,孙晓敏,温学发,周艳莲,田静,袁国富.中国通量网(ChinaFLUX)夜间CO2涡度相关通量数据处理方法研究.中国科学(D辑),2006,36(增刊 I):34-44.
[26] 孙晓敏,温学发,于贵瑞.中亚热带季节性干旱对千烟洲人工林生态系统碳吸收的影响.中国科学(D辑),2006,36(增刊2):103-110.
[28] 张雷明.中国东部南北森林样带典型生态系统碳收支特征及其生理生态学机制[D].北京:中国科学院研究生院,2006.
[35] 刘允芬,宋霞,孙晓敏,温学发,陈永瑞.千烟洲人工针叶林CO2不同高度碳通量季节变化及环境因子的影响.中国科学(D辑),2004,34(增刊2):109-117.
[36] 于贵瑞,温学发,李庆康,张雷明,任传友,刘允芬,关德新.中国亚热带和温带典型森林生态系统呼吸的季节模式及环境响应特征.中国科学D辑,2004,34(增刊2):84-94.
[37] 顾峰雪,于贵瑞,温学发,陶波,李克让,刘允芬.干旱对亚热带人工针叶林碳交换的影响.植物生态学报,2008,32(5):1041-1051.
