玉龙雪山白水1号冰川退缩迹地的植被演替
2013-09-07何元庆杨太保朱国锋牛贺文杜建括
常 丽 ,何元庆,杨太保,赵 勇,朱国锋,牛贺文,张 涛,杜建括,蒲 焘
(1.中国科学院寒区旱区环境与工程研究所/玉龙雪山冰川与环境观测研究站/冰冻圈科学国家重点实验室,兰州 730000;2.兰州大学西部环境与气候变化研究院,兰州 730000;3.兰州大学资源环境学院,兰州 730000;4.西北师范大学社会科学处,兰州 730070;5.西北师范大学地理与环境科学学院,兰州 730070)
近年来,由于人类活动(如开矿、修路等)的强度已大大超过环境的承载能力,使得自然界已不能够通过自我调节来恢复生态系统的原貌,于是便产生了大量人工原生或次生裸地,使生态环境更趋于恶化,那么应该如何“人为”的帮助受损生态系统恢复呢?要回答这个问题,必须要了解自然界中植物群落演替的过程和规律,并深入的分析演替的原因及发生机制,这对裸地的生态恢复具有重要的指导意义。目前,对于演替的原因和发生机制的理论主要有接力植物区系学说,初始植物区系学说,Connell-Slatyer三重机制学说,生活史对策演替学说,资源比率学说,Odunr-Margelef生态系统发展理论,McMahon系统概念模型和变化镶嵌体稳态学说共9种,从深层次演替研究历史可知,这是Clements等有机体论学派和Gleason等个体论学派围绕两种哲学观或尺度而展开的探讨[1]。
极地或高山冰川消退所形成的冰川裸地是研究植被原生演替及其环境解释的理想场所[2],也是生态学家们共同关注的热点之一,在此类研究中占有重要的位置[3-5]。主要采用空间代替时间的方法,由于不同地段冰川裸露的时间是不同的,并呈现出有一定规律的变化,所以,生态学家在一个地点就可以观察到一个完整的植被演替系列[2]。随着人们对原生演替研究的逐步深入,国外学者们的关注点也从“冰川裸地上的植物群落的构成”,“冰川裸地上植物的早期定居”,“冰碛物年龄与地衣直径的相关性”到“冰川裸地上植物群落的演替及其环境解释”[6]发生了很大的变化。其中最著名的研究是Crocker和Major在Alaska所做的工作,他们发现在Glacier Bay冰川消退不到100a的时间里,这里就形成了与周围植被非常相似的混交针叶林,同时,土壤条件发生了很大的变化,土壤容重和氮含量分别由1.5 g/cm3和100 kg/hm2变到0.7 g/cm3和1200 kg/hm2[4]。在我国,此类研究比较少,最早的是李逊在贡嘎山海螺沟冰川裸地上进行的研究,他认为植被发展过程、冰川退缩速度与气候变化密切相关,而且植被相对冰川退缩来说,对气候变化的响应更为敏感[7]。之后又有学者从气候变化对原生演替影响的模拟[8],植被原生演替的种间协变[9]和土壤的演替序列[10]等角度对海螺沟冰川裸地的演替系列进行了研究。在玉龙雪山地区还没有此类研究论文的发表。目前,国内外研究的热点主要集中在冰川前缘微生物的演替[11]及其对成土过程和植物定居的影响机制[12-17]。
本研究以期解决以下几个问题:(1)植被演替各个阶段植物群落的组成特征;(2)植被演替的过程及植被类型的变化和解释;(3)冰碛物的年龄。
1 研究区概况
玉龙雪山(26°59'—27°17'N,100°04'—100°15'E)位于青藏高原东南缘、横断山的最南端,南北长 35 km,东西宽18 km,南距丽江市25 km,是欧亚大陆距赤道最近的雪山,最高峰海拔5596 m。金沙江从玉龙雪山西南到东北环绕而过,从金沙江水面到玉龙雪山主峰垂直跨度近4000 m,具备了从亚热带、温带到冰川寒漠完整的7个自然带。玉龙雪山现在分布有15条现代冰川,东坡14条,西坡1条,总面积7.65 km2。最大的是白水1号冰川,长2.26 km,面积1.32 km2,平衡线高度4850 m,冰川末端海拔4365 m。该海洋型冰川区位于季风交错地带,主要受高原季风、季风环流以及西风环流的交错影响,夏季风期降水占全年降水总量的90%以上[18]。20世纪90年代后期丽江—玉龙雪山地区以冰雪景观为特色的旅游业进入快速发展期,于2007年成为首批国家5A级景区,带动和促进了玉龙雪山区域社会经济的全方位发展。
青藏高原的现代冰川是小冰期最盛时以后冰川进退变化的近期表现[19],玉龙雪山最大的一条冰川——白水1号冰川的具体变化过程如表1所示[20]。17—19世纪最盛时,白水1号冰川末端到达现在的森林上限海拔3800 m处,由于冰川前缘坡度较大,冰碛物沿冰舌外缘零散分布,分不出道数[19],这是近期白水1号冰川冰舌部分所能延伸到的最低的海拔高度。从冰川变化近一百年的历史来看,除了1957—1982年间,冰川前进了800 m之外[21],冰川基本上都处于强烈退缩的状态,据玉龙站的相关资料显示,2010年冰川末端处于海拔 4365 m 处的位置[22]。
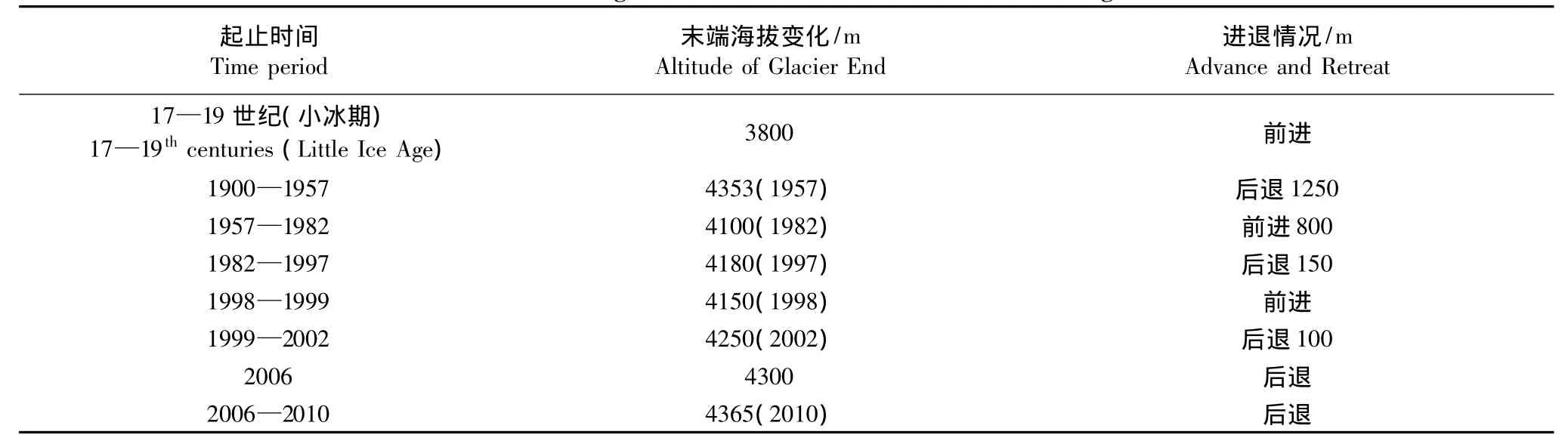
表1 自小冰期以来白水1号冰川的进退变化[20]Table1 The changes of Baishui No.1 Glacier since Little Ice Age[20]
植被调查的地点位于自小冰期以来白水1号冰川进退变化的冰川裸地上(海拔3800—4300 m),地跨亚寒带潮湿山地气候带和苔原气候带2个区域:其中海拔3800—4000 m的地带属于亚寒带潮湿山地气候带,位于夏季云雾线以上。年均气温为0—7℃,年降水量1500—1800 mm,空气湿度大,日照时数少,年均日照百分率26%,年均风速小,但多阵性大风,可达18 m/s以上。土壤有季节性冻土。海拔4000—4300 m属于苔原气候,年均温度为0—5℃,气温日较差极大,雪封时间长,土壤冻结时间长,生长季中最低气温常为负值或接近0℃,雨量大,风速大,气候多变;从植被带上来看,该区主要有耐寒垫状植物、高山草甸、杜鹃、柳树灌丛和冷杉等[23]。
2 野外调查和研究方法
2.1 样地设置
出于冰川末端地形险峻的考虑,尽量选取一些便于到达并能真实反映演替现象的地点进行植被调查,主要采用空间代替时间的方法,如图1所示,在冰川主流线的左侧,按照冰川退缩的不同年限,依次从海拔4250 m、4150 m、4000 m、3900 m和3800 m处选择5个宽约100 m,长200 m的样条(编号为Z1—Z5)作为研究地点。各样条距离2011年冰川末端的水平距离分别为:Z1—199 m,Z2—297 m,Z3—733 m,Z4—904 m,Z5—1152 m。每个样条不仅代表冰川曾经对此区域覆盖过的历史时间序列,也代表了土壤物理、化学和植物群落演替的时间序列,其中已知年限的有:Z1—10a,Z2—14a,Z5—约250a。在每个样条内选取具有代表性的10个50 cm×50 cm的样方用于调查草本,并在后两块灌木和乔木大量出现的样条内用10 m×10 m的样方调查乔木,用4 m×4 m的样方调查灌木。
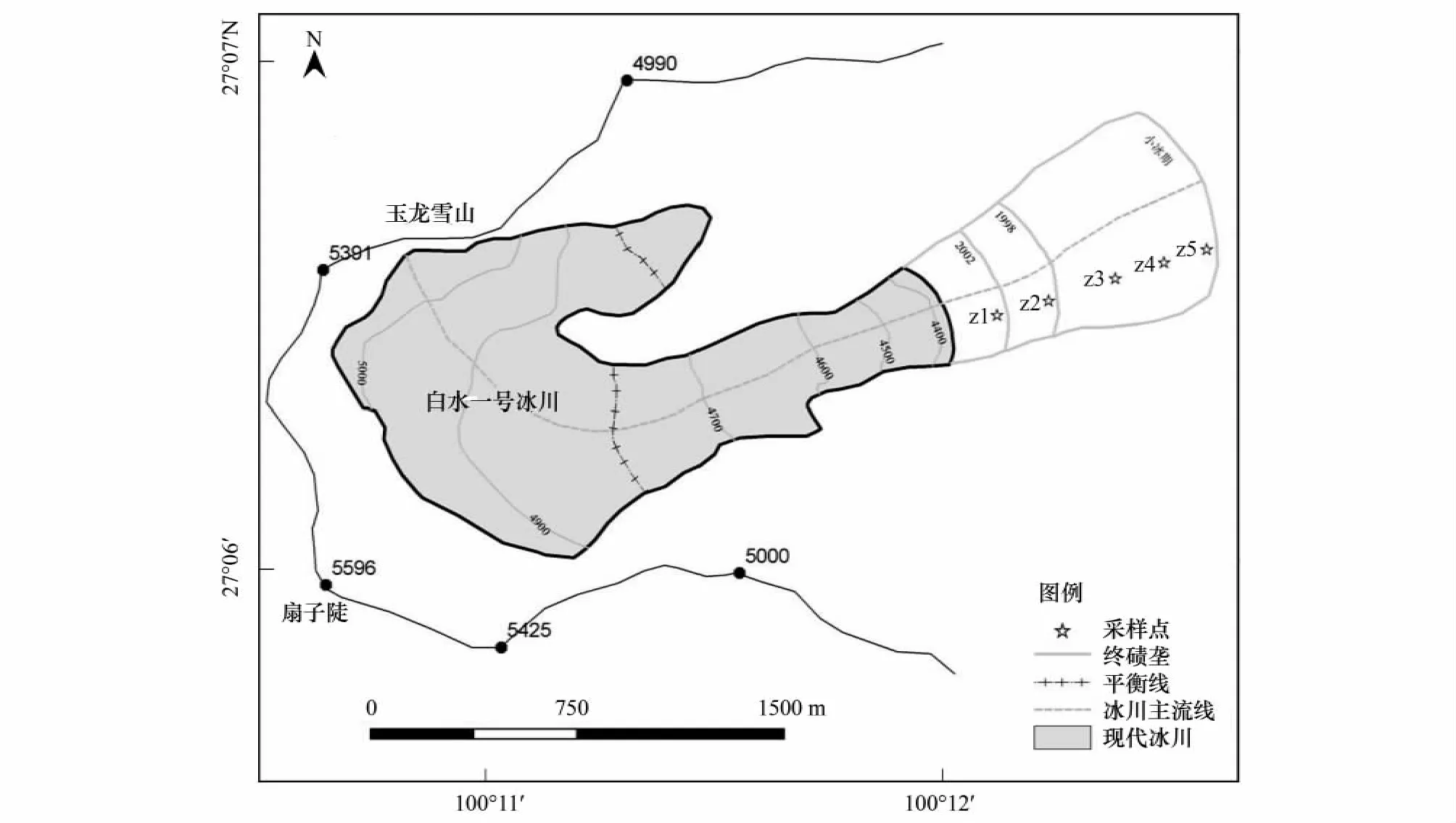
图1 玉龙雪山白水1号冰川前缘采样点分布图Fig.1 The scattergram of sampling point on the Foreland Baishui No.1 Glacier Mt.Yulong
2.2 调查内容
调查内容包括:(1)样方基本信息,包括:经纬度和海拔高度等;(2)对草本层样方,调查记录草本、灌木物种名、高度、多度、盖度和地上生物量等;(3)对乔木层,记录样方内所有活立木的种名、胸围和树高;(4)对灌木层样方,调查记录灌木、幼树的物种名、平均高度、多度和盖度。
盖度:采用估测法,地上生物量:齐地面剪割,并置于农业气象观测干燥箱内,在温度为105℃下杀青半小时,后将温度调到80℃烘干至恒重,称取干重。
3 数量分析
3.1 重要值的计算方法
重要值=(相对频度+相对高度+相对盖度)/3
3.2 因子分析方法
因子分析[24]的主要目的是用来描述隐藏在一组测量到的变量中的一些更基本的,但又无法直接测量到的隐性变量。最早由英国心理学家C.E.斯皮尔曼提出。本文的有关不同演替阶段植物群落的因子分析过程是在spss18.0里进行的,其录入数据是各个演替阶段不同物种的重要值,在分析过程中,为了使分析的数据更具代表性,剔除了一个在50个样方中只出现过1次的少见种毛果萎陵菜。
4 结果
4.1 植物群落调查结果
此次野外调查时间是2011年8月6日到12日。累计调查55个样地,发现了91种植物,已鉴定出62个种,分属于28个科,46个属,其中双子叶植物最为丰富,有23科40属54种,占冰川退缩迹地种子植物科、属、种总数的百分比分别为82.14%、86.96%、87.1%,单子叶植物3科3属5种次之,分别占 10.71%、6.52%、8.06%;裸子植物1科2属2种,分别占3.57%、4.35%、3.23%;蕨类植物1科1属1种,所占比重最小,分别是3.57%、2.17%、1.61%。而且,这些植物大多数都是多年生的草本,少有1年生草本,如总状绿绒蒿Meconopsis racemosa和多刺绿绒蒿Meconopsis horridula,包括乔木、灌木和草本3种生活型的植物。另外,从特有性上来说,有14个中国特有种,1个珍稀、濒危种(卵羽玉龙蕨Sorolepidium ovale),1个渐危种(长苞冷杉Abies georgei)。
4.2 玉龙雪山白水1号冰川裸地的植被演替
4.2.1 各演替阶段植物群落因子得分模型分析结果
对玉龙雪山白水1号冰川裸地不同演替阶段的植物群落进行因子提取,得到累计贡献率大于65% 的数个公因子,为了保证模型拥有足够信息量,对模型进行了验证,各演替阶段群落的因子分析模型均通过了KMO检验,如下表2(表中已列出单个因子贡献率)。因子得分模型的分析结果是按照因子得分大小对各演替阶段的植物进行作图(图2—图6)

表2 因子得分模型参数表Table2 The parameter list of factor analysis
4.2.2 玉龙雪山白水1号冰川裸地的植被演替
2010年白水1号冰川末端位于海拔4365 m处,从冰川末端一直到海拔4300 m(2006年冰川末端)处,到处都是比较松散的冰碛石的堆积物,没有发现任何植物,大约过了5—6a的时间,海拔4300m处的冰碛物上开始出现了高等植物(1年生的总状绿绒蒿),它们是冰川末端最早的定居者,起初它们是间隔2—3 m的距离呈线性排列的,不久,便出现了第1种多年生的草本植物——簇生卷耳,它与总状绿绒蒿是相间分布的,随后便进入所调查的演替的第1阶段。
植被演替第1阶段大约经历了10a时间,从群落外貌上来看,植物已经由线性或散点状分布发展成小斑块状分布,而且由于这是离冰川末端最近的样带,所以这里的环境异质性很高,植物一般会在背风处或岩石的缝隙等能够储备水分且利于植物扎根的地方生长,这一阶段的植物群落如图2所示,以簇生卷耳Cerastium caespitosum为优势种,多刺绿绒蒿和短柱灯芯草Juncus brachystigma为亚优势种,山景葶苈Draba oreodoxa和圆穗蓼Polygonummacrophyllum为伴生种的群落,其中簇生卷耳是在前一个阶段就在冰川裸地上定居的物种,这充分说明在演替的早期阶段它具有较强的竞争力,能够适应冰川末端较为贫瘠和严酷的自然环境。这同时也印证了因子分析的结果,簇生卷耳处于第1因子的重要地位。
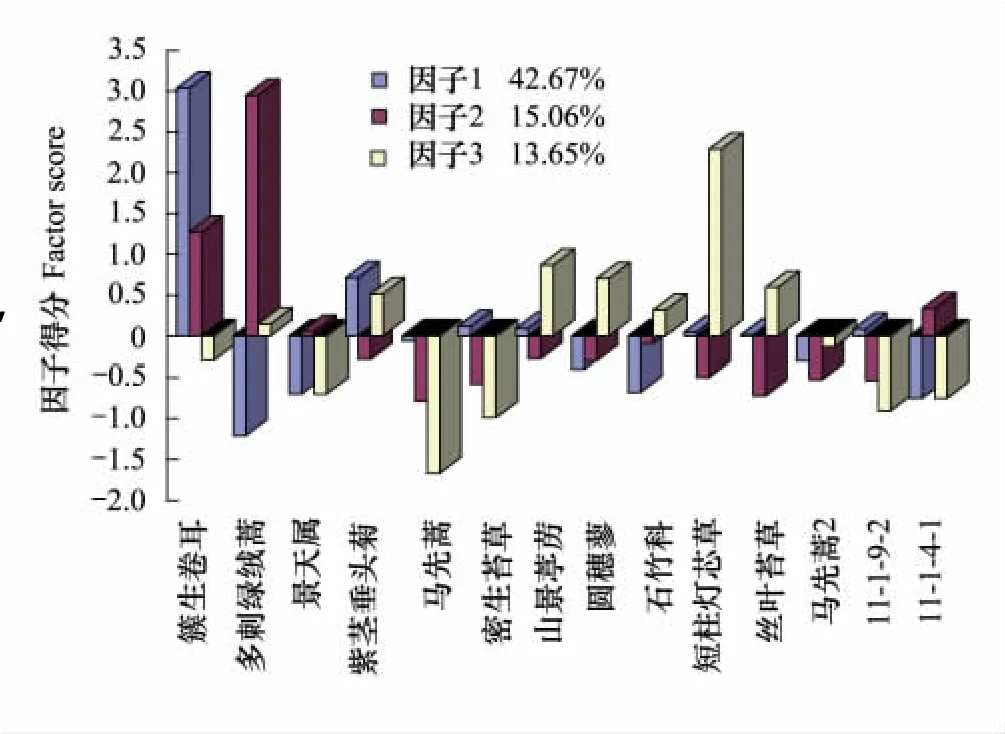
图2 白水1号冰川末端第1演替阶段植物群落结构因子得分图Fig.2 The factor score of plant community structure during the first stage of succession in Baishui No.1 glacier end
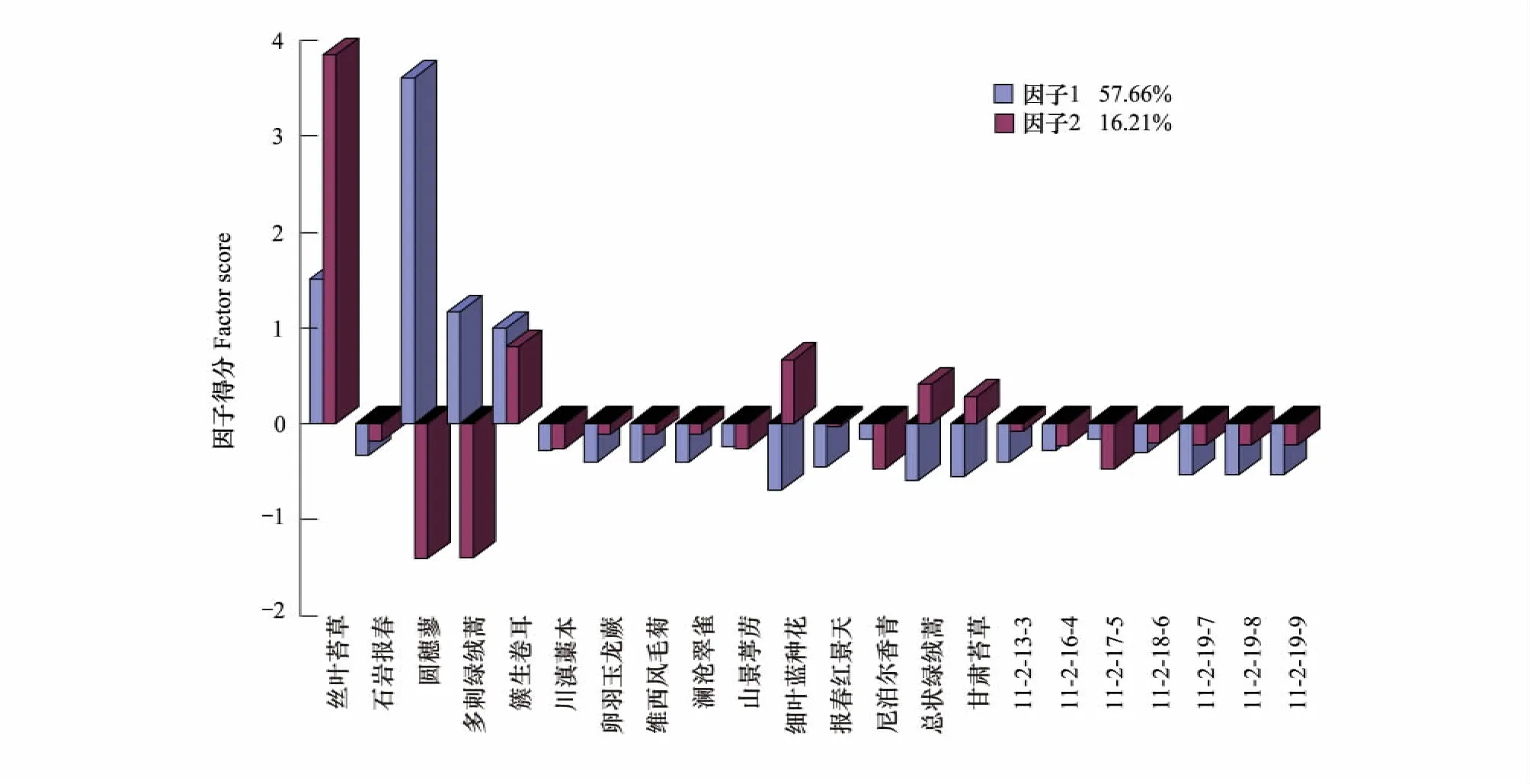
图3 白水1号冰川末端第2演替阶段植物群落结构因子得分图Fig.3 The factor score of plant community structure during the second stage of succession in Baishui No.1 glacier end
植被演替第2阶段历时14a,从群落外貌上来看,斑块变大呈丛簇状镶嵌分布,植被盖度明显增大达到20%,而且物种数比上一阶段增加了近1倍,草本层又出现了10种植物,多为莲座状、叶片和茎部密被绒毛或者在岩缝中生长的植物,如维西风毛菊 Saussurea spathulifolia、尼泊尔香青 Anaphalis nepalensis、细叶蓝钟花Cyananthus delavayi和石岩报春Primula dryadifolia等,这些植物都是能够适应流石滩的恶劣环境的典型植物。另外,在这一阶段首次出现了半灌木、灌木和乔木,分别为小垫柳Salix brachista、栎叶杜鹃Rhododendron phaeochrysum和长苞冷杉,在同海拔参照样地上的植被类型是高山杜鹃灌丛,但是却没有发现乔木的存在。同时这一阶段还第1次出现了蕨类植物和地衣附生在一起,这是流石滩上植被演替的一个主要特征。(图3)该阶段植物群落变为圆穗蓼和丝叶苔草Carex capilliformis分别形成的单优群落,其中多刺绿绒蒿和簇生卷耳为圆穗蓼群落的主要伴生种。圆穗蓼在这一阶段植物群落中占据绝对的优势(累计贡献率达到57.66%),逐渐代替了前一阶段的簇生卷耳和多刺绿绒蒿群落,使其处于从属地位。
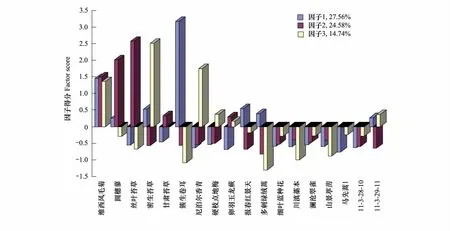
图4 白水1号冰川末端第3演替阶段植物群落结构因子得分图Fig.4 The factor score of plant community structure during the third stage of succession in Baishui No.1 glacier end
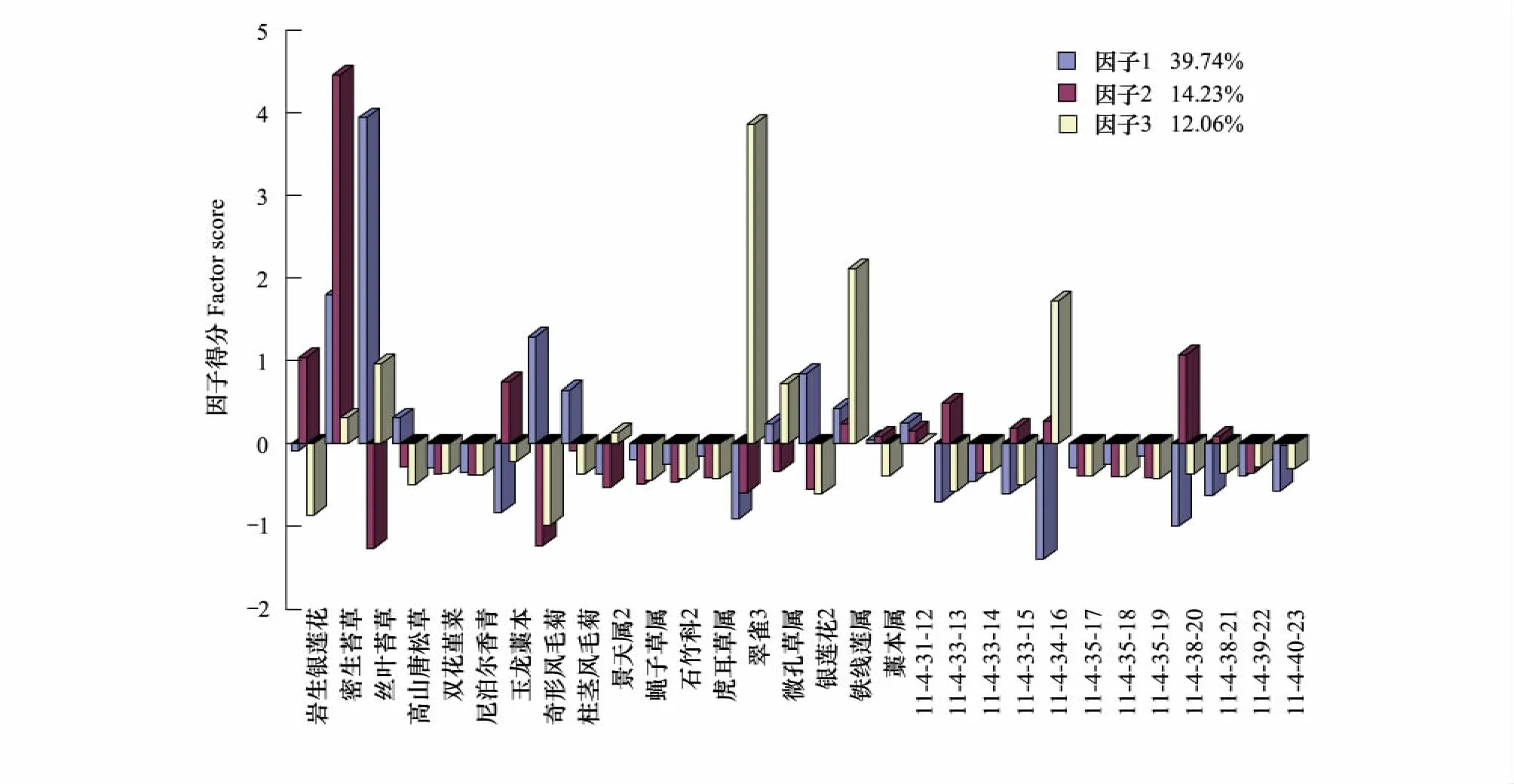
图5 白水1号冰川末端第4演替阶段植物群落结构因子得分图Fig.5 The factor score of plant community structure during the fourth stage of succession in Baishui No.1 glacier end
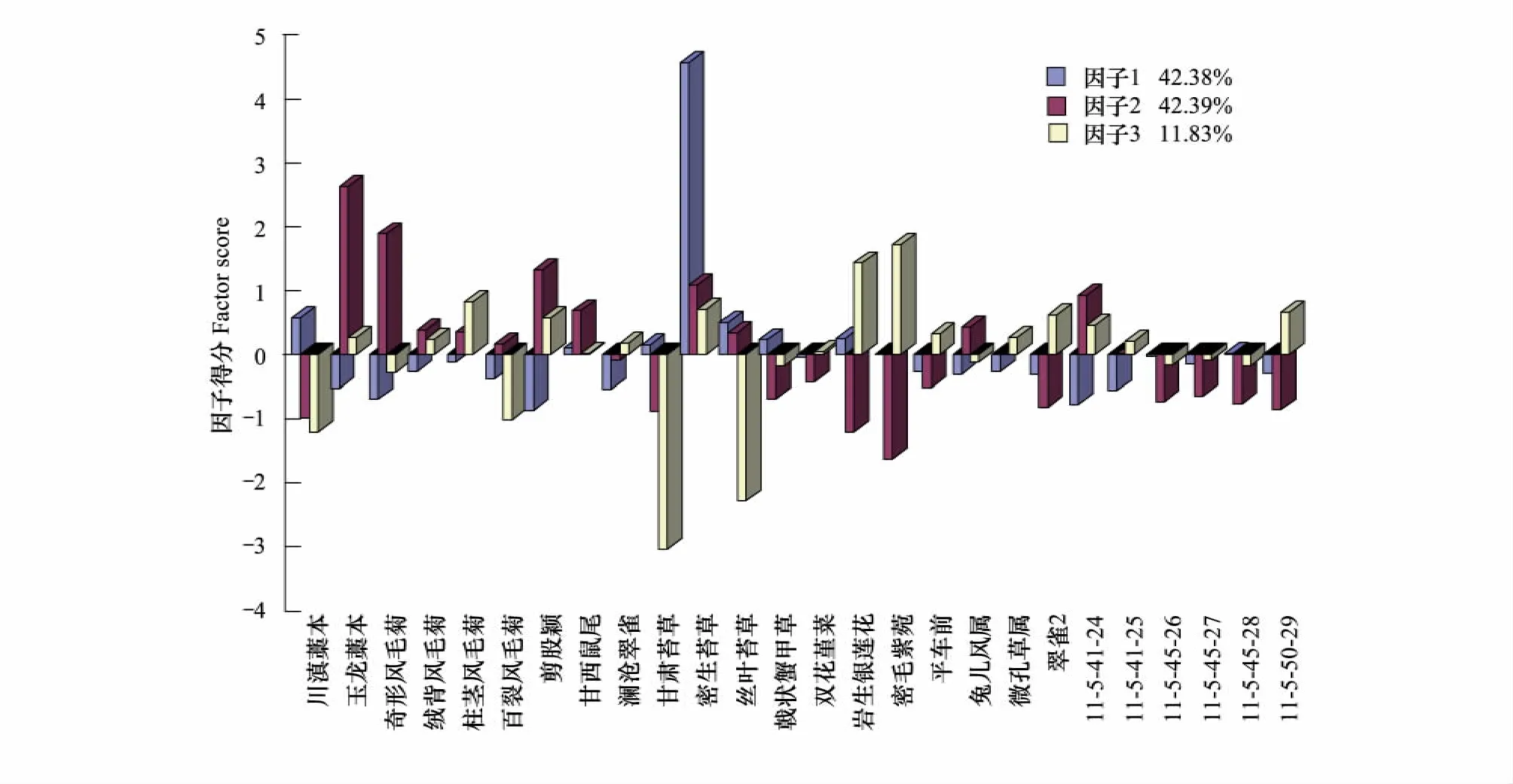
图6 白水1号冰川末端第5演替阶段植物群落结构因子得分图Fig.6 The factor score of plant community structure during the fifth stage of succession in Baishui No.1 glacier end
植被演替第3阶段的群落外貌基本上与前一阶段相似,所不同的是又出现了一种乔木矮柳Salix sp.,这是世界上最矮的树,它的适应能力很强,能在极端严酷的环境中生长。还出现了岩桦Betula calcicola这种灌木,草本又出现了喜阳的毛果萎陵菜Potentilla eriocarpa和莲座状成簇生长的硬枝点地梅Androsace rigida。该阶段群落结构发生了很大的变化,如图4所示,簇生卷耳由主要伴生种演变为极具竞争力的单优群落、圆穗蓼的竞争力逐渐减弱,趋向于与丝叶苔草形成共优群落、密生苔草Carex crebra和尼泊尔香青越来越能够适应周围的环境,形成了共优群落,并且还出现了马先蒿Pedicularis sp.群落,一共4个主要的群落。其中维西风毛菊是主要的常见种,川滇藁本Ligusticum sikiangense、澜沧翠雀Delphinium thibeticum和山景葶苈是偶见种。这一带已经位于林线的边缘,植被的盖度能达到40%,属于林缘稀疏草地。
植被演替第4阶段的植物群落完成了从草本、灌木到乔木等生活型的过渡,植被的郁闭度大大增加,从前一阶段到这一阶段似乎有些断层,因为群落外观一下子发生了飞跃式的变化,前几个阶段的植被均是比较零散而呈块状分布的,此时一下子进入了长苞冷杉的纯林分布区。造成这种断层的原因很有可能是由温度所致,由于林区上部的气温限制了大规模的森林向冰川末端扩散的速度。这一阶段草本的物种更加丰富,而且开始向阔叶和高大的方向过渡,如柱茎风毛菊Saussurea columnaris和奇形风毛菊Saussurea fastuosa,同时又出现了17个新物种,数量剧增,有7个物种消失,如卵羽玉龙蕨、硬枝点地梅、圆穗蓼等。此时灌木大量出现,又出现了6种,如小蘗Berberis sp.、毛喉杜鹃Rhododendron cephalanthum、刺红珠Berberis dictyophylla、绒叶黄花木Piptanthus tomentosus等。乔木又出现了大果红杉Larix potaninii,不过数量比较少。此阶段草本植物群落组成如图5所示,是丝叶苔草群落,密生苔草群落和翠雀Delphinium sp.群落的组合。丝叶苔草群落的主要伴生种是奇形风毛菊和银莲花Anemone sp.,翠雀群落的主要伴生种是铁线莲Clematis sp.,密生苔草的主要伴生种是岩生银莲花Anemone rupicola和玉龙藁本Ligusticum rechingerana。在这几个群落中丝叶苔草群落占有绝对优势,其累计贡献率是39.74%,因此可以认为该阶段是丝叶苔草的最适生长带,其竞争力在经历了前几个阶段后逐步增强;密生苔草比尼泊尔香青更能适应这一阶段的环境,因而由共优群落,变成密生苔草的单优群落。
植被演替第5阶段的群落外观与前一阶段基本相似,乔木层的优势种仍然是长苞冷杉,草本层的优势种是密生苔草,而且草本又出现了10种新的更加喜湿、高大、阔叶的植物,如密毛紫菀Aster vestitus、兔儿风Ainsliaea sp.、剪股颖Agrostis matsumurae、甘西鼠尾Salvia przewalskii等。如图6所示这一阶段的主要植物群落是密生苔草的单优群落,玉龙藁本+奇形风毛菊的共优群落和甘肃苔草 (Carex kansuensis)+丝叶苔草的共优群落3个主要的植物群落。其中,密生苔草单优群落的竞争力最强,其累计贡献率为42.38%,而其他两个群落均处于劣势。此外,从叶片形态上来看,密生苔草的叶子较丝叶苔草的来说,已不是针形的而是较窄的片状,这说明这一地带的气温比前一阶段有所升高。此阶段出现的新物种数量较前一阶段有明显减少,可能是由于随着林冠层郁闭度的增大,草本层植物能够利用到的阳光越来越少,使得喜阳的草本逐渐减少所致。
总的来说,在经历了从草本—灌木—乔木的250a的演进后,维管束植物的演替已趋于完成。演替早期植物的定居速度是比较缓慢的,演替中期植物的侵占速度陡然增加,到演替后期植被覆盖裸地的速度又开始重新缓和,也就是说演替后期已进入生态平衡阶段即,演替的顶级阶段。此外,随着演替时间的推进,各个优势种的高度、密度、生物量和盖度都是呈增大的趋势,但频度的变化相对来说是比较平稳的。这说明伴随着植被的演替,土壤、温度、水分、辐射等环境因子均变得更为缓和,更加适合植物的定居及生长。
5 讨论与结论
5.1 植被演替中植被类型的变化及解释
一般来讲,旱生原生演替要经过地衣、苔藓、草本植物和木本植物4个阶段[25]。然而白水1号冰川裸地植被演替过程中植被类型的变化不是完全遵循这样的规律,而是按照草本—灌木—乔木的顺序变化的,区别在于:首先从高等有花植物而不是地衣或苔藓开始进行演替,随后一年生草本和多年生草本共生,直到第3个阶段(海拔4166 m处)出现灌木的同时,才出现了苔藓,这是流石滩植物群落形成的一个特点。因为在石砾及碎石间细碎物质积累较快,有花植物的繁殖体能够借助稳定的土壤条件生根定居[26]。由此看来,自然演替过程更倾向于个体演替论的观点,即演替并非一定要按照固定的顺序进行,而是取决于早期定居的物种是什么。
那么植物定居下来之后,各物种及种群间是如何替代的呢?从生态位理论和竞争排斥原理相结合的角度来看,其适应性及生态幅的大小有如下顺序:乔木>灌木>多年生草本>1年生草本,因而演替的过程总会朝着适应性强及生态幅大的物种的方向演进,而且在演替的整个过程中各群落及种对之间通常是相互独立并共存着的,符合R.H.Whittaker的群落中全部物种间的关联类型呈正态分布的观点[27],说明它们之间的生态位是分离的,重叠较少,存在直接种间竞争的物种较少,群落内部竞争以种内竞争为主。以草本植物演替为例我们可以作更进一步的解释:第一演替阶段,虽然环境比较恶劣,土壤养分也比较贫瘠,但是相对于物种数目来说,资源还是相对丰富的,因而各个群落间仍是能够共存的,物种间的关系以种内竞争为主,如簇生卷耳,多刺绿绒蒿和短柱灯芯草这3个群落,它们能共存是因为其生态位较狭,且重叠较少,那么生境中一定有未被利用的资源,它们将通过激烈的种内竞争来扩展资源利用的范围,同时能够适应该机制的物种必将在进化中获得好处,而且种内竞争的结果又促使两物种间生态位的接近[27]。而山景葶苈和圆穗蓼是短柱灯芯草的主要伴生种,说明它们之间生态位是比较靠近的,而且对资源利用具有相似的诉求,于是它们之间的种间竞争占优势,将会导致两种结果:一是某一物种的灭亡,二是通过生态位的分离而使两物种得以共存[27]。虽然这几个种对间是竞争关系,但是该群落在这一阶段却处于次要地位,所以该阶段仍主要表现为种内竞争。进入第2阶段后,圆穗蓼和丝叶苔草成为最为主要的两个群落,圆穗蓼在与短柱灯芯草和山景葶苈的竞争中胜出,圆穗蓼与山景葶苈的生态位分化后得以共存,而短柱灯芯草却灭亡了,这时圆穗蓼与簇生卷耳和多刺绿绒蒿的生态位开始接近,它们之间出现了种间竞争。总之,演替过程中每个阶段群落的组成特征都是不尽相同的,印证了生态位理论的观点,不同物种具有不同的生态位,所以在不同的演替阶段构成了不同的植物群落,那些能够成为优势种的物种通常是生态位较宽,适应性较强,在多个演替阶段能够存活,并能利用多种资源的物种。在演替过程中,随着土壤的养分、水分和气温等外界条件的改善,能够被植物利用的资源越来越多,同时,外界环境条件也更能适应植物的定居,于是在此定居的物种就越来越多,为了避免直接的竞争,使得群落组成更加稳定,植物自身的生态位不断发生分化,从草本—灌木—乔木的转变就是生态位分化的一种表现,物种通过占据不同的生态位,使得各自都能尽可能最大化的利用更多资源,以期在进化过程中获得更大的好处。因此,我们可以认为演替过程中物种替代就是在其生态位不断的分化与接近,激烈的种内与种间竞争过程中持续下去的,直到处于某种动态平衡为止,这时群落的外观基本不变,然而群落内部却一直处于变化之中。
5.2 小冰期终碛垄年龄的估计
表3列出了玉龙雪山白水1号冰川区和贡嘎山海螺沟冰川区自然地理概况的异同点,虽然两个区域的海拔分布范围及受控的天气系统是不同的,而且白水1号冰川前缘从完全裸露到有草本(海拔4300 m处发现总状绿绒蒿)出现经历了约6a的时间,其草本地被占优势的阶段持续了约20—30a的时间,比贡嘎山海螺沟冰川前缘出现草本植物的时间提前了2a,但是海螺沟冰川草本地被占优势的时间较短(8—15a)[7]。这可能是因为白水1号冰川区分布的海拔高度更高的原因,低温条件使得森林无法快速入侵并占据主导地位。但是从区域角度上来看,二者均位于青藏高原东南缘的温冰川区,从演替方向来看,植被演替都是按照裸地-草本-灌木-乔木的顺序演进的,并最终形成了云、冷杉纯林。基于此笔者认为通过对比二者的演替过程再结合何元庆等用树轮及冰川变化资料重建的小冰期以来的气温变化资料[20],来大致估计小冰期终碛垄的年龄是可行的。由此可以推断出白水1号冰川前缘小冰期最盛时的冰碛垄(海拔3800—3900 m)的年龄最早在170a左右,甚至比海螺沟冰川的170a更短[7]。

表3 玉龙雪山白水1号冰川区与贡嘎山海螺沟冰川区的自然地理概况对比Table3 The comparison of the geography overview for the areas Baishui No.1,Glacier Mt.Yulong and Hailuogou Glacier,Mt Gongga
致谢:玉龙雪山冰川与环境观测研究站,昆明植物所丽江高山生态站和西北师范大学生命科学学院的工作人员在样品采集、物种鉴定及论文思路等方面给予帮助,特此致谢。
[1] Ren H,Cai X A,Rao X Q,Zhang Q M,Liu SZ.The theory on succession of plant community.Ecologic Science,2001,20(4):59-67.
[2] Xu Z Q,Li W H,Bao W K,Xu Q.A review on primary succession of vegetation.Acta Ecologica Sinica,2005,25(12):3383-3389.
[3] Jones GA,Henry GH R.Primary plant succession on recently deglaciated terrain in the Canadian High Arctic.Journal of Biogeography,2003,30(2):277-296.
[4] Crocher R L,Major J.Soil development in relation to vegetation and surface age at Glacier Bay,Alaska.Journal of Ecology,1955,43(2):427-448.
[5] Hodkinson ID,Webb N P,Coulson SJ.Primary community assembly on land-the missing stages:why are the heterotrophic organisms always there first?Journal of Ecology,2002,90(3):569-577.
[6] Stork A.Plant immigration in front of retreating glaciers,with examples from the Kebnekajse Area,Northern Sweden.Geografiska Annaler,1963,45(1):1-22.
[7] Li X,Xiong SF.Vegetation primary succession on glacier foreland in Hailuogou,Mt.Gongga.Mountain Research,1995,13(2):109-115.
[8] Huo C F,Cheng G W,Lu X Y,Fan J H,Xiao F P.Dynamic simulation of climate change impacts on forest primary succession in Gongga Mountain,southwestern China.Journal of Beijing Forestry University,2012,32(1):1-6.
[9] Luo J.Interspecific covariation during primary succession on the east slope of Gongga Mountain.Mountain Research,1996,14(4):235-238.
[10] He L,Tang Y.Soil development along primary succession sequences on moraines of Hailuogou Glacier,Gongga Mountain,Sichuan,China.Catena,2008,72(2):259-269.
[11] Wang X X.Primary succession of microflora along glacier forefield of Urumqi Glacier No.1.[D].Urumqi:Xinjiang University,2010.
[12] Sigler W V,Crivii S,Zeyer J.Bacterial succession in glacial forefield soils characterized by community structure,activity and opportunistic growth dynamics.Microbial Ecology,2002,44(4):306-316.
[13] Sigler W V,Zeyer J.Microbial diversity and activity along the forefields of two receding glaciers.Microbial Ecology,2002,43(4):397-407.
[14] Nicol G W,Tscherko D,Embley T M,Prosser J I.Primary Succession of Soil Crenarchaeota across a Receding Glacier Foreland.Environment Microbiology,2005,7(3):337-347.
[15] Shinpei Y,Masaki U,Takayuki N,Hiroshi K,Hiroshi K.Characterization of Soil Microflora on a successional glacier Foreland in the high Arctic on Ellesmere Island,Nunavut,Canada using Phospholipid fatty acid analysis.Polar Bioscience,2006,19:73-84.
[16] Schütte U M E,Abdo Z,Bent SJ,Williams CJ,Schneider G M,Solheim B,Forney L J.Bacterial succession in a glacier foreland of the High Arctic.The ISME Journal,2009,3(11):1258-1268.
[17] Wang X X,Zhang T,Sun J,Zhang X B,Ling Z Q,Lou K.Ecological characterization of soil microflora in primary succession across glacier forefield:a case study of Glacier No.1 at the Headwaters of Urumqi River.Acta Ecologica Sinica,2010,30(23):6563-6570.
[18] Li Z X,He Y Q,He X Z,Jia W X,Wang S J,Chang L,Zhu G F,Xin H J.Environmental records from a shallow profile,Baishui No.1 Glacier,Mt.Yulong.Earth and Environment,2009,37(4):360-365.
[19] Zheng B X.Quaternary glaciation and glacier evolution in the Yulong mount,Yunnan.Journal of Glaciology and Geocryology,2000,22(1):53-61.
[20] He Y Q,Zhang Z L,Yao T D,Chen T,Pang H X,Zhang D.Modern changes of the climate and glaciers in China's Monsoonal temperate-glacier region.Acta Geographica Sinica,2003,58(4):550-558.
[21] Li J J,Su Z.Glaciers in the Hengduan Mountains.Beijing:Science Press,1996
[22] Li ZX,He Y Q,Pu T,Jia WX,He X Z,Pang H X,Zhang NN,Liu Q,Wang SJ,Zhu GF,Wang SX,Chang L,Du JK,Xin H J.Changes of climate,glaciers and runoff in China's monsoonal temperate glacier region during the last several decades.Quaternary International,2010,218(1/2):13-28.
[23] Zhao X T,Zheng B X,Xiao Z R,Li T S.Research on investigation,development and programmings of tourism resources at Glacier Park of Mt.Yulong.Beijing:Instisute of Geology,Chinese Academy of Sciences,1998.
[24] Liu W W.A primary study on vegetation in the Water-Level-Fluctuating Zone of Three Gorges Reservoir[D].Wuhan:Wuhan Botanical Garden,Chinese Academy of Sciences,2011.
[25] Kimmins J P.Forest Ecology.New York:Mcmillan,1987.
[26] Jiang H Q,Zhu W M.A studying on vegetation of the flowstone beach in Mt.Yulong.Journal of Yunnan University,1957,(4):163-178.
[27] Li B,Yang C,Lin P.Ecology.Beijing:Higher Education Press,1999.
参考文献:
[1] 任海,蔡锡安,饶兴权,张倩媚,刘世忠.植物群落的演替理论.生态科学,2001,20(4):59-67.
[2] 许中旗,李文华,鲍维楷,许晴.植被原生演替研究进展.生态学报,2005,25(12):3383-3389.
[7] 李逊,熊尚发.贡嘎山海螺沟冰川退却迹地植被原生演替.山地研究,1995,13(2):109-115.
[8] 霍常富,程根伟,鲁旭阳,范继辉,肖飞鹏.气候变化对贡嘎山森林原生演替影响的模拟研究.北京林业大学学报,2010,32(1):1-6.
[9] 罗辑.贡嘎山东坡植被原生演替的种间协变.山地研究,1996,14(4):235-238.
[11] 王晓霞.乌鲁木齐河源1号冰川前缘微生物原生演替[D].乌鲁木齐:新疆大学,2010.
[17] 王晓霞,张涛,孙建,张雪兵,李忠勤,娄恺.冰川前缘土壤微生物原生演替的生态特征——以乌鲁木齐河源1号冰川为例.生态学报,2010,30(23):6563-6570.
[18] 李宗省,何元庆,和献中,贾文雄,王世金,常丽,朱国锋,辛惠娟.我国典型季风海洋型冰川积累区冰雪剖面的环境记录.地球与环境,2009,37(4):360-365.
[19] 郑本兴.云南玉龙雪山第四纪冰期与冰川演化模式.冰川冻土,2000,22(1):53-61.
[20] 何元庆,张忠林,姚檀栋,陈拓,庞洪喜,章典.中国季风温冰川区近代气候变化与冰川动态.地理学报,2003,58(4):550-558.
[21] 李吉均,苏珍.横断山冰川.北京:科学出版社,1996.
[23] 赵希涛,郑本兴,肖泽榕,李铁松玉龙雪山冰川公园旅游资源调查,规划和深层次开发研究报告[R].北京:中国科学院地质研究所,1998.
[24] 刘维伟.三峡水库消落区植被初步研究[D].武汉:中国科学院武汉植物园,2011.
[26] 姜汉侨,朱维明.丽江玉龙山流石滩植被初步观察.云南大学学报:玉龙雪山植被调查专号,1957,(4):163-187.
[27] 李博,杨持,林鹏.生态学.北京:高等教育出版社,1999.
