五鹿山国家级自然保护区物种多样性海拔格局
2013-09-07何艳华张钦弟张直峰苗艳明毕润成
何艳华,闫 明,张钦弟,张直峰,苗艳明,毕润成
(山西师范大学生命科学学院,临汾 041004)
生物多样性格局与生态因子之间的关系一直是生态学研究的热点领域[1]。物种多样性可以和许多生态梯度结合起来[2],海拔梯度包含了温度、水分和光照等因素的影响[3],而且在海拔梯度上温度的变化速率要比在纬度梯度上快1000倍[4]。物种多样性的海拔梯度格局,可反映出物种的生物学、生态学特性、分布状况及其对环境的适应性[5]。近年来对植物多样性沿海拔梯度分布格局的研究较多[6-9]。一些研究表明物种多样性在中海拔达到最大(单峰格局)[6]、而另一些研究则表明随海拔升高物种多样性逐渐下降[7]或与海拔无关[8]不同山地和不同生活型的物种多样性海拔分布格局不同,这可能与山地所处的区域环境条件、山体的相对高度和地质地貌等众多因素有关[9]。五鹿山自然保护区先后被批准成立为省级、国家级自然保护区[10],主要保护以褐马鸡为主的野生动物和白皮松林、辽东栎林为主的森林生态系统[11]前人已经对它的主要植被类型和分布[12],种子植物名录[13]等进行了研究。五鹿山保护区地理位置的特殊性使得其在植物区系组成上表现为南北过渡、东西交汇的特点,具有复杂性、多样性、过渡性和特殊性。但近年来由于人口压力以及大规模的经济开发,保护区生物多样性逐年锐减,白皮松、辽东栎林的破坏严重,褐马鸡最适宜的栖息地逐渐缩小[11],因此加强保护迫在眉睫。本文以五鹿山自然保护区植被垂直带谱的调查数据为基础,分析其乔木层、灌木层和草本层的α多样性、β多样性以及总物种的γ多样性在海拔梯度上的分布格局。以期为五鹿山自然保护区的保护提供理论基础同时也为阐明山地植物多样性的垂直分布格局积累基础资料。
1 研究区概况
五鹿山自然保护区地处吕梁山南端,蒲县和隰县交界处,位于36°23'45 ″—36°38'20″N,111°8'—111°18'E 之间。全区面积20617.3 hm2,主要山峰有尖山、红军寨、沙冒顶等,山势险要,沟大谷深,主峰五鹿山最高海拔1946.3 m。该区位于暖温带季风型大陆性气候区,是东南季风区的边缘。年平均气温8.7℃,极端最高气温36.4℃,极端最低气温-23.2℃。无霜期150—180 d。降水量年均500—560 mm左右,最高899.5 mm,最低346.4 mm。保护区内的土壤呈明显的垂直分布,在区内自上而下土壤类型共分为棕壤、褐土、草甸土、山地草甸土4 个土类[11]。
2 研究方法
2.1 样地设置
2010年10月在研究区内选择有代表性的山体,从山麓1040m到山顶1877m沿海拔梯度设置样地,每隔100m设置3个样地,在物种比较丰富的地段增设1个样地。每个样地包含4个10m×10m的乔木样方,每个乔木样方的对角线两个角上分别设一个5m×5m的灌木样方,四角及中心分别设一个1m×1m的草本样方。共设置样地30个(图1)。其中包括乔木样方120个,灌木样方240个,草本样方600个。具体调查指标包括:乔木层的种类、胸径、树高、冠幅;灌木层的种类、高度、盖度;草本层的种类、平均高度、盖度;样方所在地的经度、纬度、海拔、坡度、坡向、坡位等指标。
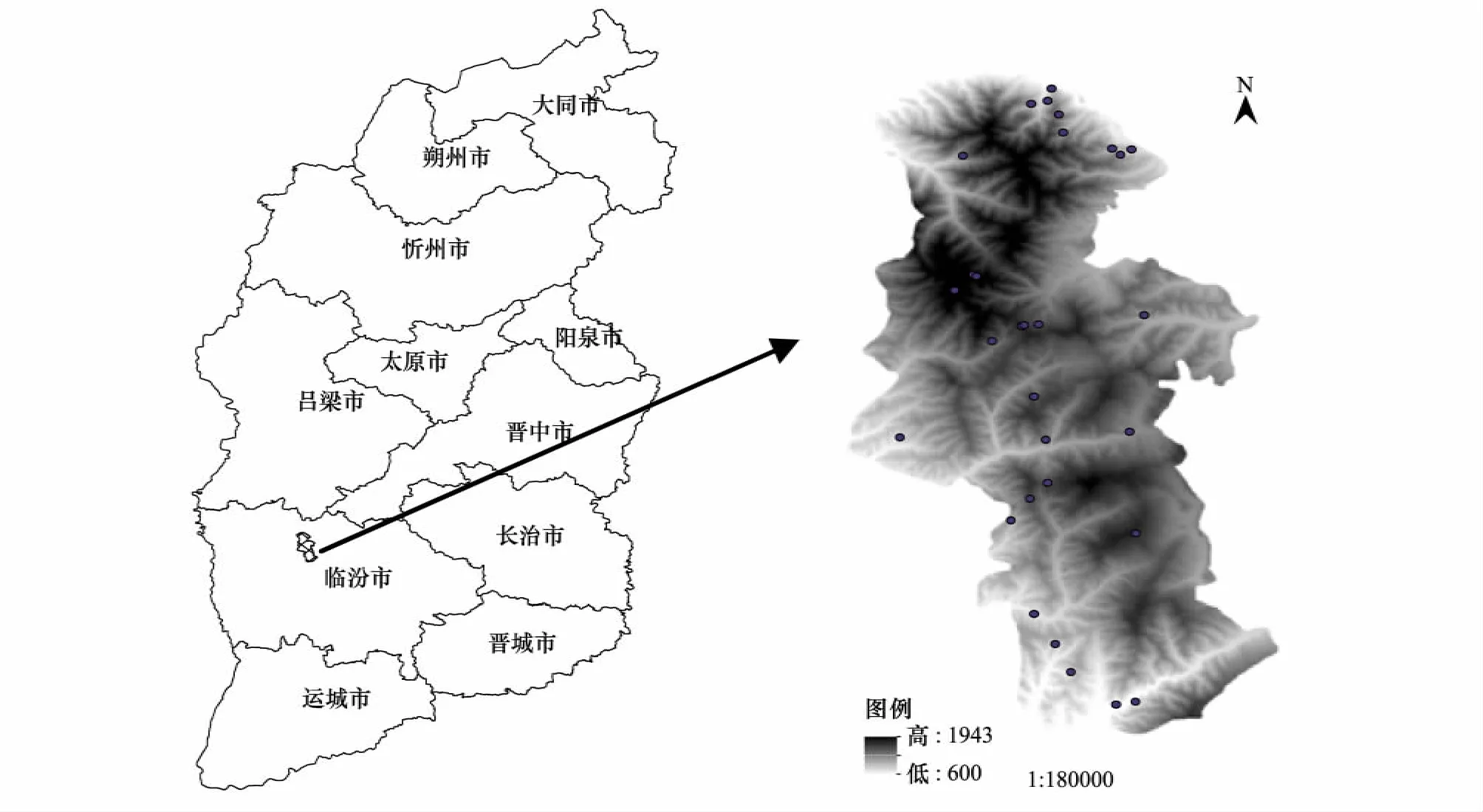
图1 研究区及样地的地理位置Fig.1 The location of study area and the investigated plots
2.2 数据处理
分别计算每个样地内乔木层、灌木层、草本层的α多样性、β多样性、总物种的γ多样性以及乔木层的胸高截面积。计算方法如下:
α-多样性

β-多样性
β多样性指沿环境梯度不同生境群落之间物种组成的相异性或物种沿环境梯度的更替速率[14]。文中选取Sørensen相异性指数来反映β-多样性,公式如下:

式中,S为Sørensen相异性指数,a和b分别为样方i和j内物种数目,W为两个样方内的共有种数目。研究中将五鹿山自然保护区植物群落以100梯度为间隔分段,计算海拔间群落物种相异性指数。
γ-多样性
1972年Whittker将γ多样性定义为在一个地理范围(如一个景观、一个岛屿等)内各生境中的物种丰富度[15]多指区域或大陆尺度的物种数量。该方法一直沿用至今,将五鹿山自然保护区植物群落从山脚至山顶以100梯度为间隔划分为9个海拔带,以每个海拔梯度中总物种数目(总物种丰富度S)来表示区域内的γ多样性。
乔木层的胸高截面积
乔木层的胸高截面积(cm2/m2),即每个样方内所有乔木胸高(距地面高1.3m)处的树干截面积之和。
2.3 回归分析
广义可加模型(GAM)是一种拟合非参数函数来估计响应变量与解释变量之间关系的模型,是广义线性模型(GLM)的进一步推广,优点是拟合曲线能最大限度的符合原始数据[16]。前人已经运用该模型对贺兰山木本植物物种多样性的海拔格局[17]、伊犁河谷山地植物群落物种多样性分布格局[18]环境因子对巴山冷杉-糙皮桦混交林物种分布及多样性的影响[19]等进行了研究,并得出了较好的结果。其一般形式为:
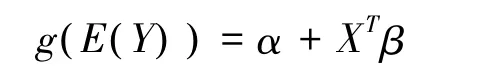
式中,α、β分别相当于线性回归模型中的截距和回归系数矩阵,XT表示预测变量矩阵,E(Y)为响应变量Y的期望,g()表示联结函数,而S()表示平滑函数。GAM分析在CANOCO软件(Version4.5)中完成。
GAM预测模型拟合结果的评价指标为模型的偏差(D2),其计算公式如下:

式中,D2相当于线性回归模型中的回归系数R2,反映了模型可解释的偏差。而无效偏差是指模型在只剩截距时的偏差,剩余偏差指拟合状态下仍然不能解释的偏差。理想状况下的D2值应当为1,表明没有剩余偏差,模型完全解释[20]。
3 结果分析
3.1 群落特征概况
采用重要值作为各物种在群落中的优势度指标,以表达不同物种在群落中的功能地位,以及反映其在群落中的分布格局状况。乔木层是森林生态系统的主要成分,其组成成分决定了林下的灌木层和草本层的组成和结构[7],因而首先选用乔木的重要值进行分类。对研究区森林生态系统组成种重要值排序(表1)分析发现五鹿山自然保护区内建群种主要是刺槐、油松、白皮松、辽东栎等;灌木层主要优势种为黄刺玫、水栒子、陕西荚蒾等;草本层主要优势种为披针叶苔草、铁杆蒿、淫羊藿等。
五鹿山自然保护区内沿海拔升高,受水热组合的变化,地带性植被发生着有序的变化(表2)。群落内的建群种和优势种也发生着有序的变化,大体上,从低海拔到高海拔依次为刺槐、白皮松、油松、茶条槭、华北落叶松、辽东栎,其中白皮松、辽东栎和油松的生态幅较宽、分布范围较广,刺槐、茶条槭、华北落叶松等只在一些小生境中成为优势种或建群种。灌木层中主要优势种为黄刺玫、陕西荚蒾、水栒子;草本层,除在较低海拔下优势种为铁杆蒿,其余海拔下优势种均为披针叶苔草。
运用GAM模型拟合乔木的胸高截面积沿海拔的分布格局(图2),可看出乔木的胸高截面积沿海拔梯度大体呈递增的变化趋势。因为从低海拔到高海拔群落类型逐渐由针叶林向辽东栎林过渡特别是进入辽东栎林以后(从海拔1610m开始)乔木的胸高截面积随海拔升高急剧增大,而低海拔(<1110m)群落类型为刺槐疏林,林冠郁闭度较小。
3.2 α多样性变化
区内α多样性随海拔梯度变化如图3从图中可看出不同层次的物种丰富度对环境变化的敏感程度不同。
物种丰富度
(1)乔木层,总体上中海拔区域物种丰富度高,低海拔、高海拔区物种丰富度较低。其中海拔1260—1480 m的几个样地最高,这是因为该区域的群落类型为辽东栎+白皮松混交林和茶条槭+油松混交林,乔木层物种比较丰富,含有较多的亚优势种和伴生种,如:油松、白桦、华北落叶松。
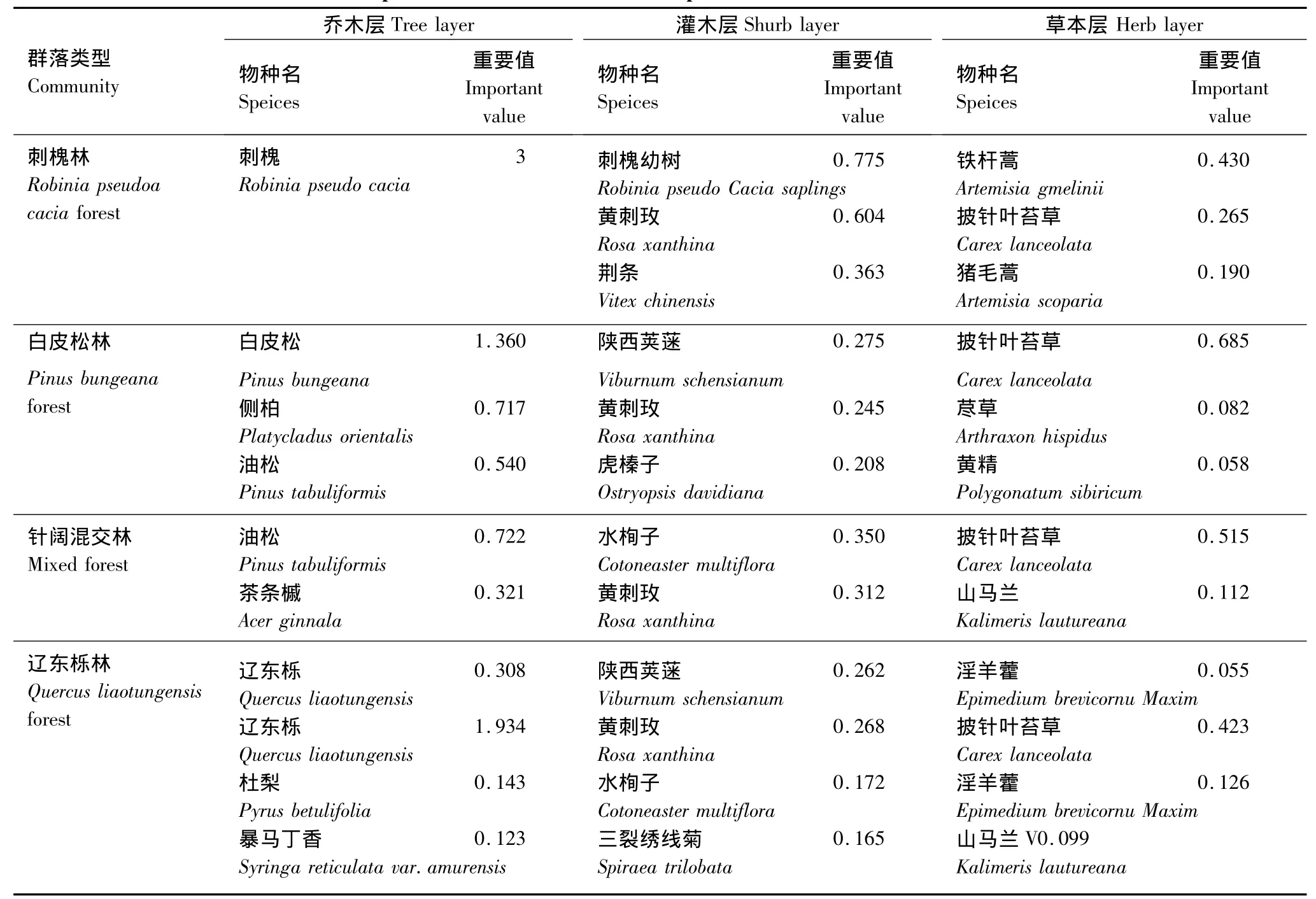
表1 五鹿山自然保护区不同群落类型主要优势种重要值排序Table1 Importance values of the dominant species of wulu Mountain Nature Reserve

表2 沿海拔梯度主要群落类型及主要组成种Table2 Both community type&community composition vary along the altitudinal gradient
(2)灌木层,除海拔1160和1260 m处由于受小生境(沟谷空气湿润、土壤肥沃)的影响物种丰富度指数较高分别为19、20外,其余海拔下物种丰富度大体呈以中高海拔1507 m处为峰值的偏峰分布格局,是因为该处的水热组合较好,且乔木层物种丰富度较低(D=3),为灌木层物种的发育提供了良好的光环境。
(3)草本层,在海拔在海拔1040—1400 m之间之间草本层的物种丰富度随海拔升高逐渐下降,大于海拔1400m时,丰富度指数变化不大。这是因为,乔木层的胸高截面积随海拔的升高呈递增的趋势(图1)而草本层的物种丰富度与乔木层的胸高截面积呈明显的负相关关系(图4)。即林木对地面的遮蔽越严重,草本层的物种丰富度就越低[8,21]。到海拔1400m以后除了受乔木层的胸高截面积的影响外还受水热组合、自然选择等多方面因素的影响,变化趋势变得明显。
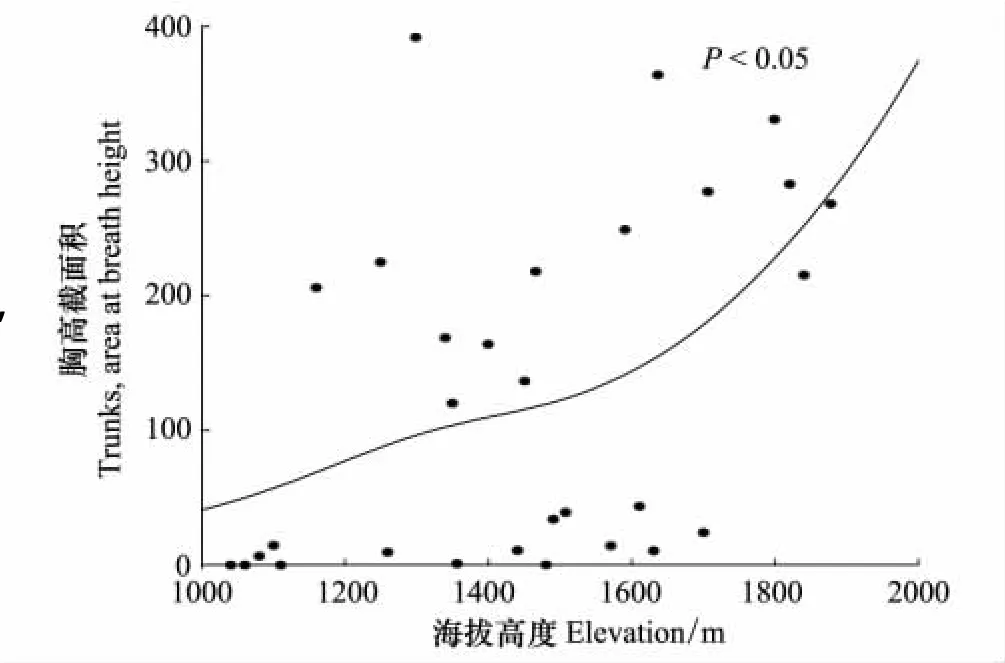
图2 森林群落乔木胸高截面积沿海拔梯度的变化Fig.2 Changes of total Trunks,area at breath height in all forest plots along altitude
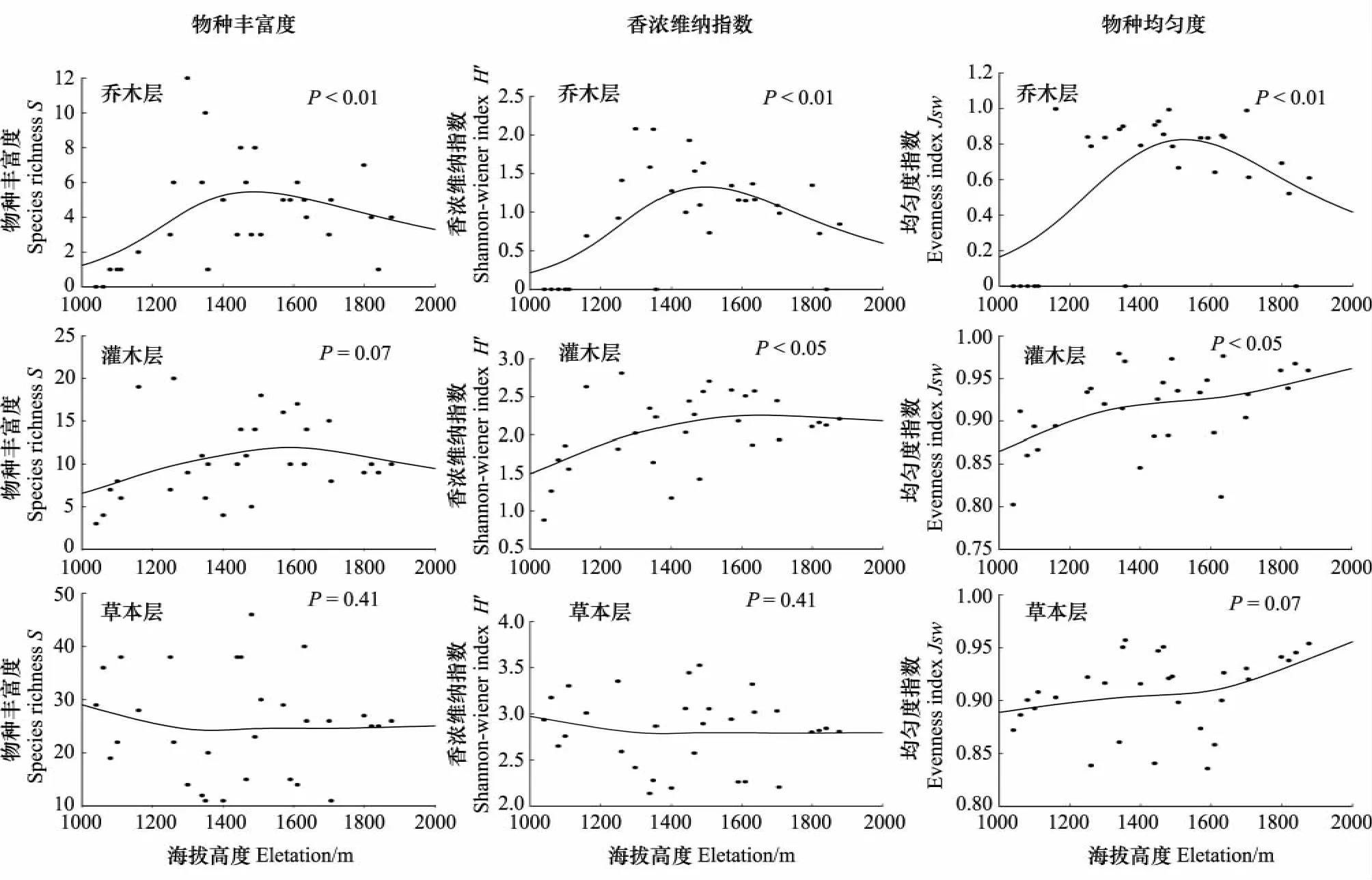
图3 五鹿山自然保护区多样性海拔梯度的变化Fig.3 Varition of α-diversity along altitudinal gradient of wulu Mountion nature reserve
Shannon-Wiener多样性指数:Shannon-Wiener多样性指数与物种丰富度变化趋势大体相同。
物种均匀度:
乔木层的均匀度指数变化趋势与其丰富度指数、香浓维纳指数的变化趋势相同,呈显著的单峰分布格局(P<0.01),这是因为中海拔群落类型为针阔混交林,物种丰富,优势种作用不明显。灌、草层随海拔升高常见种与稀少种的差距减小,均匀度指数大体呈递增的趋势。
3.3 草本物种丰富度与乔木胸高截面积的关系
以不同海拔乔木的胸高截面积为横坐标,以草本的物种丰富度为纵坐标作图(图6)结果表明,随着乔木胸高截面积的增大,草本的物种丰富度大体呈递减趋势,即草本的物种丰富度与乔木胸高截面积呈负相关关系。样方内乔木的胸高截面积越大,林冠郁闭度也越大[22]。林冠郁闭度通过光照、局部温湿度、紫外线强度等方面的差异对林下物种尤其是草本层的分布产生明显影响[23]。
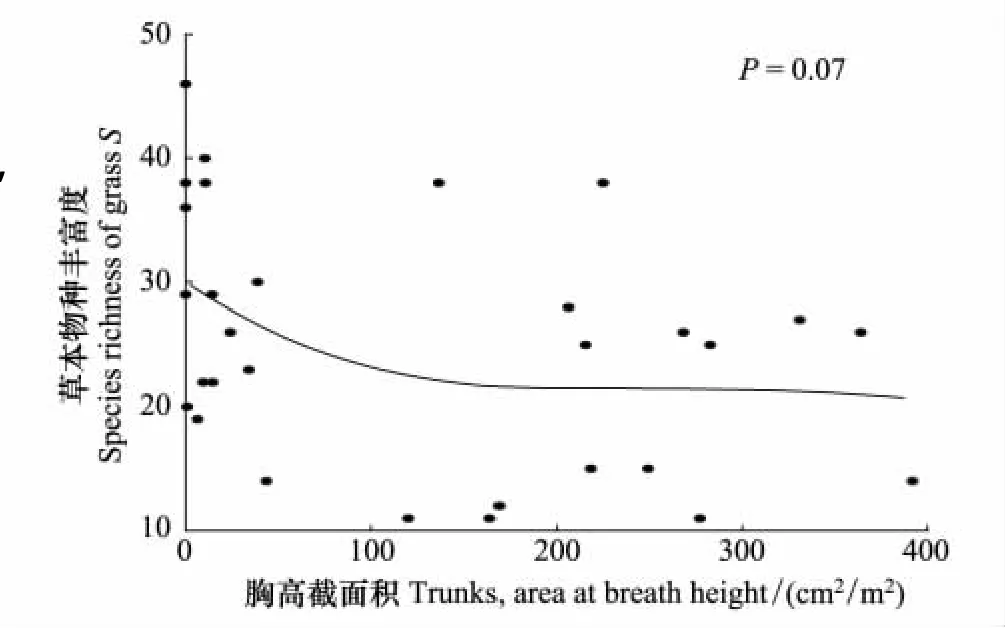
图4 草本物种丰富度与乔木胸高截面积的关系Fig.4 The relationships of species richnesses of grass and total Trunks,area at breath height in all forest plots
3.4 β多样性
五鹿山自然保护区β多样性(图5)沿海拔梯度大体呈“~”型变化。较低海拔和山顶附近的物种更替速率较快,较低海拔白皮松林向针阔混交林过渡带物种变化速率较快,β多样性出现一个峰值,较高海拔(>1750 m)由于水热组合的影响,适生物种的种类发生变化,同时数量急剧减少(例如:油松主要分布在海拔1200—1700 m的地方,槭、椴树分布的最高海拔为1750 m[11])物种变化速率较快,β多样性较大。其余海拔下变化比较平缓。
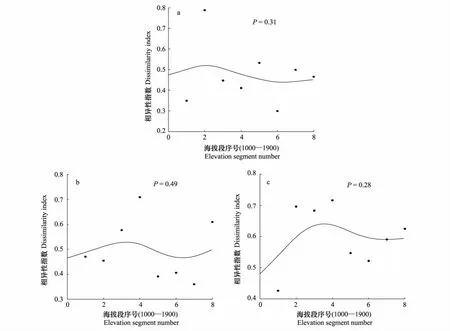
图5 五鹿山自然保护区群落间物种相异性(β-多样性)沿海拔梯度的变化Fig.5 variation ofβ-diversity along the altitudinal gradient of Wulu Mountion narure reserve
3.5 γ多样性
文中将五鹿山自然保护区按海拔100m为区间分段,以各海拔段内出现的物种数为纵坐标作图,具体结果如图6所示,五鹿山自然保护区物种丰富度沿海拔梯度大体呈单峰分布格局,在中高海拔1400—1500m之间出现最大值。较低海拔区,人为干扰较大,降水量较少,土壤贫瘠,适合生存的物种少,随着海拔升高,降水量增加,同时气温降低,地表蒸发量减少,土壤中有机物质的含量增加,适合生存的物种增多,物种丰富度逐渐达到峰值,到高海拔区域,气候寒冷,同时生长季较少的降水量也限制了植物的生存,物种丰富度逐渐降低。
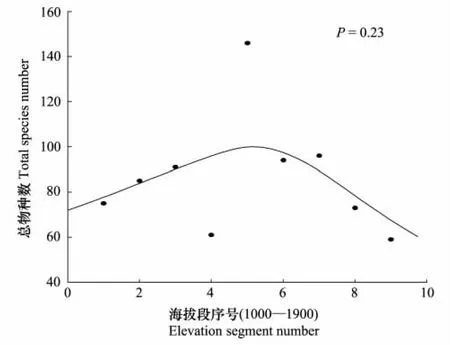
图6 五鹿山自然保护区总物种丰富度(γ多样性)沿海拔梯度的变化Fig.6 Variation of γ-diversity along the altitudinal gradient of Wulu Mountion nature reserve
3.6 海拔与物种分布相关性分析
GAM拟合模型中D2相当于相关系数R2,反映了模型可解释的偏差。从表3可看出海拔与不同层次的各多样性指数的相关程度不一,其中与乔、灌层的α多样性的相关性程度较大,D2介于18.2%到63.2%之间,而与草本层的α多样性的相关性程度较小,D2介于3.8%到17%之间,这是因为,除海拔外,乔木层的胸高截面积也对草本层的物种分布产生一定的影响(D2=17.9%);与乔、灌、草 β多样性的相关系数分别为14.7%、24.7%、39.4%;与总物种γ多样性的相关系数为38.8%。
4 结论和讨论
4.1 讨论
α多样性具有多种环境梯度(海拔梯度、纬度梯度等)格局,其中海拔梯度是环境梯度中的主导梯度之一[24-28]。物种多样性沿海拔梯度的分布格局一般有5种形式,分别是随海拔升高先降低后升高、先升高后降低(单峰曲线)、单调升高、单调下降和没有明显格局。[8-9]且不同生活型的植物种类沿海拔梯度呈现不同的多样性格局[29]。
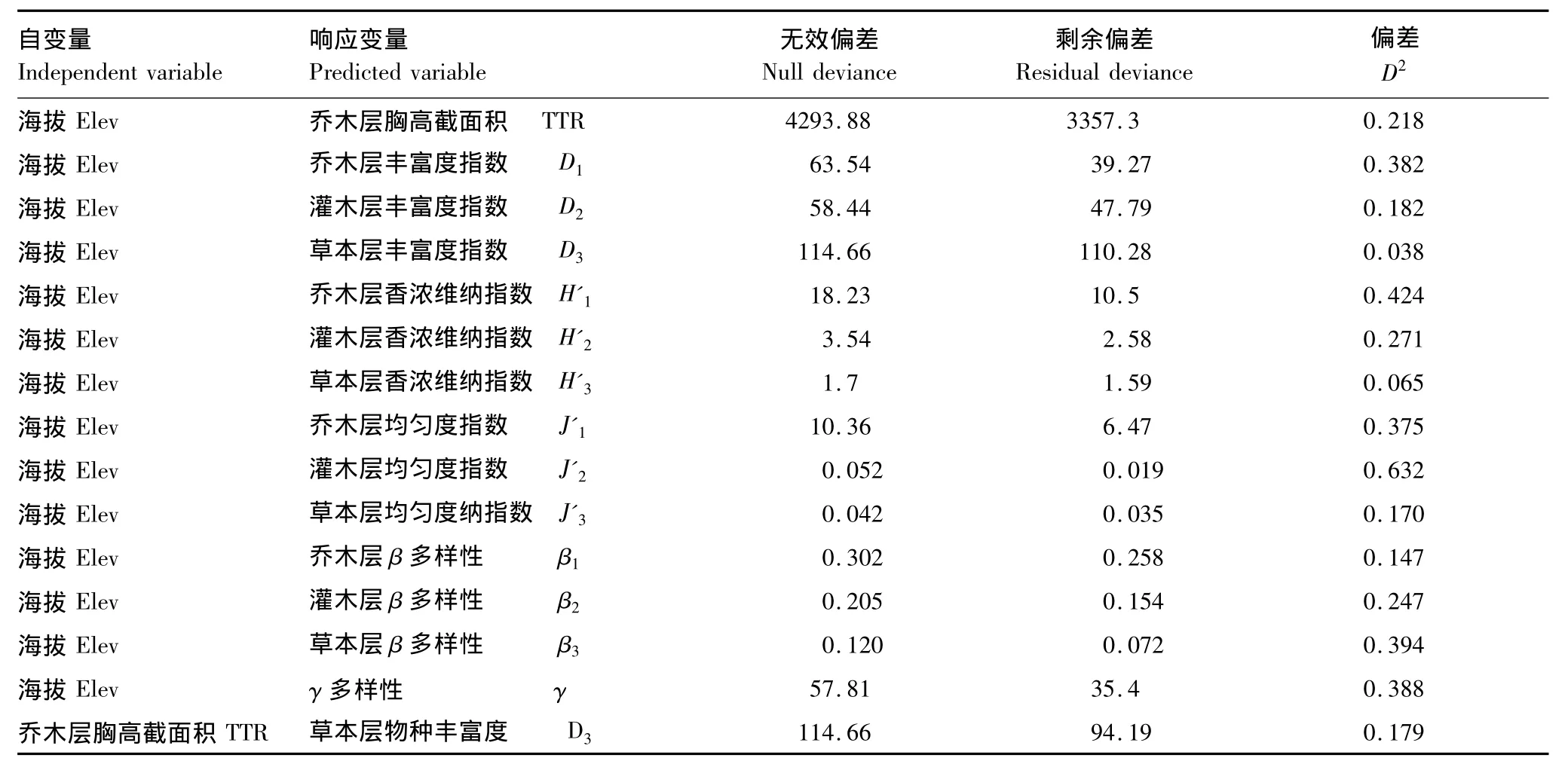
表3 GAM预测模型的偏差表Table3 D2 value of GAM model
五鹿山自然保护区α多样性受群落类型更替的影响,沿海拔梯度变化发生一定的波动。且乔灌草3层的变化规律及敏感程度都不同。物种丰富度指数在多数海拔下为草本层﹥灌木层﹥乔木层,在低海拔区,人类活动影响较大、降水量少、土壤贫瘠[11]乔灌层物种组成单一,群落类型为纯林(刺槐林、白皮松林),物种丰富度较低,随着海拔的升高,人类活动减少,水热组合发生变化,地带性植被发生更替,建群种和伴生种也发生变化,形成针阔混交林,乔灌层的物种丰富度升高,到一定海拔,温度降低到只适合部分物种生长时,地带性植被由混交林逐渐变为纯林(辽东栎林),乔灌层丰富度又开始降低,只有在中间海拔时人为干扰较小,且水热组合最佳,乔灌层丰富度最高。草本层的物种丰富度在较低海拔下,由于受乔木层郁闭度的影响,随海拔升高,物种丰富度逐渐减低,到海拔1400m以后由于受乔木层的郁闭度、水热组合、自然选择等多方面因素的综合影响,变化趋势逐渐变得不明显。
Shannon-Wiener指数与丰富度指数变化趋势大体相同,与相关研究[28,30]结论相同。
乔木层的均匀度指数与其丰富度指数、香浓维纳指数的变化趋势大体相同也呈单峰分布格局,这是因为中间海拔的水热组合适宜、干扰较小为乔木层创造了有利的生存条件,优势种的作用不明显,均匀度较高。灌木层和草本层的均匀度指数,随海拔升高逐渐变大,这是因为,均匀度指数除与丰富度指数和Shannon-Wiener多样性指数有一定的相关性之外,还与个体数目或生物量等指标在各个物种中分布的均匀程度有关[30]。研究区随海拔升高,灌草层中常见种与稀少种的差距逐渐减小,群落向着均方向发展,所以随海拔升高,均匀度指数逐渐增大,这与实际调查时研究区现状相符。
β多样性反映了生境间的多样性,控制β多样性的主要生态因子有水分、土壤、地貌及干扰等[14],它可以反映较大距离上群落组成的差异[31-33],也可以是小范围内环境梯度快速变化下物种组成的更替[6,34]。在对β多样性随海拔梯度变化格局的研究中,不同学者也得出了不同结论。大致可分为3种:(1)β多样性性沿海拔梯度的无规律变化,β多样性的高值一般出现在群落的交错带[35];(2)植物的β多样性随海拔升高而单调下降[35];(3)在低海拔变化不大,而在高海拔地区随着海拔升高急剧上升[36]。本研究中用Sørensen指数反映β多样性,该指数可以很好地反映研究区内群落物种组成沿海拔梯度发生的变化,群落间Sørensen指数沿海拔梯度峰值出现的区域为建群种发生更替的过渡带,即不同群落类型的交错带[37]。研究发现五鹿山自然保护区β多样性沿海拔梯度大体呈“~”型变化,较低海拔白皮松林向针阔混交林过渡带物种变化速率较快,β多样性出现一个峰值,较高海拔由于水热组合的影响,适生物种的种类发生变化,同时数量急剧减少物种变化速率较快,β多样性较大。
γ多样性随海拔梯度呈现两种较为普遍的分布格局:1)偏峰分布格局(不同研究中峰值出现的区域不同);2)线性递减的负相关格局[25]。五鹿山自然保护区γ多样性的峰值出现在中海拔地区。这是因为,低海拔的干旱、人为干扰和高海拔的严寒限制了物种的生长[11]中间海拔区域温度和降水量配置适宜,环境和资源条件优越,形成了物种多样性的峰值区域[17,25,38]。这与Abrams提出的单峰关系学说比较相似,在生产力较低时,多样性随生产力的增加而增加,但当生产力达到足够高时多样性会下降,亦即在群落生产力达到中等水平时出现最高的多样性[39]。
运用GAM模型拟合各多样性指数与海拔之间的相关关系,除草本层的α多样性由于受乔木层郁闭度影响与海拔拟合模型的偏差较小,其余模型的偏差介于17%—63.2%之间,说明海拔是影响物种分布的重要因素。但拟合模型的偏差均小于1表明除海拔外还有其他的因素影响物种的分布,例如在较小的海拔范围内地形因子可能会对物种分布产生一定的影响[18]这有待进一步的研究。
4.2 结论
在自然因素和人为因素共同作用下,五鹿山自然保护区乔、灌、草及总物种的多样性随海拔梯度发生一定的变化,中间海拔人为干扰较小,同时水热资源配置良好,导致乔、灌层及总物种多样性在中间海拔出现峰值,而草本层在水热组合、人为干扰和乔木层郁闭度的共同影响下多样性指数呈现随海拔升高递减的趋势。
调查中发现:五鹿山中海拔地段(1400—1500 m)生物多样性较高,白皮松在黑虎沟海拔1110—1260 m地段生长良好,辽东栎从海拔1260m到山顶都有分布,其中海拔1610m以上几乎为纯林,在瓦罐庙、红军寨等地分布典型。白皮松是我国特有树种,分布面积非常狭小,辽东栎是五鹿山的地带性植被,是中国落叶阔叶林的典型群落,也是褐马鸡的重要栖息地[11]。因此建议不同海拔段采取不同的保护措施,以取得较好的效果。
致谢:感谢北京师范大学Lamber Nyobe博士对英文摘要的润色。
[1] Zhao Z Y,Wang R H,Yin CH,Zhang H Z,Wang L.Species diversity and spatial heterogeneity of plant communities in piedmont plain of South Slope of Tianshan Mountains.Acta Botanica Boreali-Occidentalia Sinica,2007,27(4):784-790.
[2] Wang G H,Zhou GS,Yang L M,Li ZQ.Distribution,species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains,Gansu,China.Plant Ecology,2002,165(2):169-181.
[3] Gaston K J.Global patterns in biodiversity.Nature,2000,405(6783):220-226.
[4] Walter H.Vegetation of the Earth.2nd ed.New York:Springer,1979:1-35.
[5] Deng H B,Hao Z Q,Wang Q L.The changes of co-possession of plant species between communities with altitudes on northern slope of Changbai Mountain.Journal of Forestry Research,2001,12(2):89-92.
[6] Liu Z L,Zheng C Y,Fang J Y.Changes in plant species diversity along an elevation gradient on Mt.Xiao Wutai,Hebei,China.Biodiversity Science,2004,12(1):137-145.
[7] Zhao SQ,Fang JY,Zong Z J,Zhu B,Shen H H.Composition,structure and species diversity of plant communities along an altitudinal gradient on the northern slope of Mt.Changbai,Northeast China.Biodiversity Science,2004,12(1):164-173.
[8] He J S,Chen W L.A review of gradient changes in species diversity of land plant communities.Acta Ecologica Sinica,1997,17(1):91-99.
[9] Liu X L,Shi Z M,Yang D S,Liu S R,Yang Y P,Ma Q Y.Advances in study on changes of biodiversity and productivity along elevational gradient in mountainous plant community.World Forestry Research,2005,18(4):27-34.
[10] Zhang X L,Jing S H.Biodiversity research and development measures for Five-deer Mountain nature reserve.Journal of Shanxi Agricultural Sciences,2011,39(7):696-698.
[11] Bi RC,Wei X Z,Duan Z S,Yu WL,Cui BY.Scientific Investigating Report for Nature Reserves in Wulu Mountain of Shanxi.Beijing:Chinese Science and Technology Press,2004:1-249.
[12] Mao F F,Xu B S.The main vegetation types and their distribution in the Wu Lu Mountainous region.Journal of Shanxi Normal University:Natural Science Edition,1988,2(1):61-67.
[13] Wei X Z,Su JX,Liu WZ,Wang Y Y,Zhang L P,BI R C,Li X F,Huang L,Li JF,Cui B Y.A List of the Plants in the Wulushan.Journal of Shanxi Normal University:Natural Science Edition,2003,17(3):72-79.
[14] Cui B S,Zhao X S,Yang Z F,Tang N,Tan X J.The response of reed community to the environment gradient of water depth in the Yellow River Delta.Acta Ecologica Sinica,2006,26(5):1533-1541.
[15] Whittaker R H.Evolution and measurement of species diversity.Taxon,1972,21(2/3):213-251.
[16] Shen Z H,Zhao J.Prediction of the spatial patterns of species richness based on the plant topography relationship:an application of GAMs approach.Acta Ecologica Sinica,2007,27(3):953-963.
[17] Zhu Y,Kang M Y,Jiang Y,Liu QR.Altitudinal pattern of species diversity in woody plant communities of mountain Helan,northwestern China.Journal of Plant Ecology,2008,32(3):574-581.
[18] Xu Y J,Chen Y N,Li W H,Fu A H,Ma X D,Gui D W,Chen Y P.Distribution pattern and environmental interpretation of plant species diversity in the mountainous region of Ili River Valley,Xinjiang,China.Chinese Journal of Plant Ecology,2010,34(10):1142-1154.
[19] Ren X M,Yang GH,Wang DX,Qin X W,Liu ZX,Zhao SX,Bai Y.Effects of environmental factors on species distribution and diversity in an Abies fargesii-Betula utilis mixed forest.Acta Ecologica Sinica,2012,32(2):605-613.
[20] Wang SP,Zhang ZQ,Zhang JJ,Zhu JZ,Guo JT,Tang Y.modeling the effects of topography on the spatial distribution of secondary species in a small watershed in the Gully region on the Loess Plateau,China.Acta Ecologica Sinica,2010,30(22):6102-6112.
[21] Xie J Y,Chen L Z.Species diversity characteristics of deciduous forests in the warm temperate zone of north china.Acta Ecologica Sinica,1994,14(4):337-344.
[22] Zhu Y,Kang M Y,Liu Q R,Su Y,Jiang Y.Comparison of forest structure and floristic composition of coniferous forest in Helan Mountain.Geographical Research,2007,26(2):305-313.
[23] Fang JY.Exploring altitudinal patterns of plant diversity of China's mountains.Biodiversity Science,2004,12(1):1-4.
[24] Rahbek C.The role of spatial scale and the perception of large-scale species-richness patterns.Ecology Letters,2005,8(2):224-239.
[25] Tang Z Y,Fang J Y.A review on the elevational patterns of plant species diversity.Biodiversity Science,2004,12(1):20-28.
[26] Hao Z Q,Yu D Y,Yang X M,Ding Z H.α diversity of communities and their variety along altitude gradient on northern slope of Changbai Mountain.Chinese Journal of Applied Ecology,2002,13(7):786-789.
[27] Dolezal J,Srutek M.Altitudinal changes in composition and structure of mountain-temperate vegetation:a case study from the Western Carpathians.Plant Ecology,2002,158(2):202-221.
[28] Chen J,Guo Y L,Lu X L,Ding S Y,Su S,Guo J J,Li Q X.Species diversity of herbaceous communities in the Yiluo River Basin.Acta Ecologica Sinica,2012,32(10):3021-3030.
[29] Ojeda F,Marañón T,Arroyo J.Plant diversity patterns in the Aljibe Mountains(S.Spain):a comprehensive account.Biodiversity and Conservation,2000,9(9):1323-1343.
[30] Ma K P,Huang JH,Yu SL,Chen L Z.Plant community diversity in Dongling Mountain,Beijing,China Ⅱ.Species richness,evenness and species diversities.Acta Ecologica Sinica,1995,15(3):268-277.
[31] Condit R,Pitman N,Leigh E G Jr,Chave J,Terborgh J,Foster R B,Núñez P,Aguilar S,Valencia R,Villa G,Muller-Landau H C,Losos E,Hubbell SP.Beta-diversity in tropical forest trees.Science,2002,295(5555):666-669.
[32] Normand S,Vormisto J,Svenning J C,Grández C,Balslev H.Geographical and environmental controls of palm beta diversity in paleo-riverine terrace forests in Amazonian Peru.Plant Ecology,2006,186(2):161-176.
[33] Bridgewater S,Ratter J A,Ribeiro J F.Biogeographic patterns,β-diversity and dominance in the Cerrado biome of Brazil.Biodiversity and Conservation,2004,13(12):2295-2318.
[34] Hao Z Q,Yu D Y,Wu G,Deng H B,Jiang P,Wang QL.Analysis onβdiversity of plant communities on northern slope of Changbai Mountain.Acta Ecologica Sinica,2001,21(12):2018-2022.
[35] Vázquez G J A,Givnish T J.Altitudinal gradients in tropical forest composition,structure,and diversity in the Sierra de Manantlan.Journal of Ecology,1998,86(6):999-1020.
[36] Odland A,Birks H J B.The altitudinal gradient of vascular plant richness in Aurland,western Norway.Ecography,1999,22(5):548-566.
[37] Lu X L,Hu N,Ding SY,Fan Y L,Liao B H,Zhai Y J,Liu J.The pattern of plant species diversity of Funiu Mountain Nature Reserve.Acta Ecologica Sinica,2010,30(21):5790-5798.
[38] Whittaker R J,Willis K J,Field R.Scale and species richness:towards a general,hierarchical theory of species diversity.Journal of Biogeography,2001,28(4):453-470.
[39] Abrams P A.Monotonic or unimodal diversity-productivity gradients:what does competition theory predict?Ecology,1995,76(7):2019-2027.
参考文献:
[1] 赵振勇,王让会,尹传华,张慧芝,王雷.天山南麓山前平原植物群落物种多样性及空间分异研究.西北植物学报,2007,27(4):784-790.
[6] 刘增力,郑成洋,方精云.河北小五台山北坡植物物种多样性的垂直梯度变化.生物多样性,2004,12(1):137-145.
[7] 赵淑清,方精云,宗占江,朱彪,沈海花.长白山北坡植物群落组成、结构及物种多样性的垂直分布.生物多样性,2004,12(1):164-173.
[8] 贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征.生态学报,1997,17(1):91-99.
[9] 刘兴良,史作民,杨冬生,刘世荣,杨玉坡,马钦彦.山地植物群落生物多样性与生物生产力海拔梯度变化研究进展.世界林业研究,2005,18(4):27-34.
[10] 张晓玲,景慎好.五鹿山国家级自然保护区生物多样性研究与发展对策.山西农业科学,2011,39(7):696-698.
[11] 毕润成,魏学智,段长锁,尉文龙,崔本义.山西省五鹿山自然保护区科学考察报告.北京:中国科学技术出版社,2004:1-249.
[12] 毛芬芳,徐宝珊.五鹿山地区主要植被类型和分布.山西师大学报:自然科学版,1988,2(1):61-67.
[13] 魏学智,苏俊霞,刘维仲,王玉燕,张丽萍,毕润成,李晓峰,黄磊,李俊峰,崔本一.五鹿山种子植物名录.山西师大学报:自然科学版,2003,17(3):72-79.
[14] 崔保山,赵欣胜,杨志峰,唐娜,谭学界.黄河三角洲芦苇种群特征对水深环境梯度的响应.生态学报,2006,26(5):1533-1541.
[16] 沈泽昊,赵俊.基于植物-地形关系的物种丰富度空间格局预测——GAMs途径的一种应用.生态学报,2007,27(3):953-963.
[17] 朱源,康慕谊,江源,刘全儒.贺兰山木本植物群落物种多样性的海拔格局.植物生态学报,2008,32(3):574-581.
[18] 徐远杰,陈亚宁,李卫红,付爱红,马晓东,桂东伟,陈亚鹏.伊犁河谷山地植物群落物种多样性分布格局及环境解释.植物生态学报,2010,34(10):1142-1154.
[19] 任学敏,杨改河,王得祥,秦晓威,刘振学,赵双喜,白宇.环境因子对巴山冷杉-糙皮桦混交林物种分布及多样性的影响.生态学报,2012,32(2):605-613.
[20] 王盛平,张志强,张建军,朱金兆,郭军庭,唐寅.黄土残塬沟壑区流域次生植被物种分布的地形响应.生态学报,2010,30(22):6102-6112.
[21] 谢晋阳,陈灵芝.暖温带落叶阔叶林的物种多样性特征.生态学报,1994,14(4):337-344.
[22] 朱源,康慕谊,刘全儒,苏云,江源.贺兰山针叶林结构特征与种类组成的比较.地理研究,2007,26(2):305-313.
[23] 方精云.探索中国山地植物多样性的分布规律.生物多样性,2004,12(1):1-4.
[25] 唐志尧,方精云.植物物种多样性的垂直分布格局.生物多样性,2004,12(1):20-28.
[26] 郝占庆,于德永,杨晓明,丁之慧.长白山北坡植物群落α多样性及其随海拔梯度的变化.应用生态学报,2002,13(7):785-789.
[28] 陈杰,郭屹立,卢训令,丁圣彦,苏思,郭静静,李乾玺.伊洛河流域草本植物群落物种多样性.生态学报,2012,32(10):3021-3030.
[30] 马克平,黄建辉,于顺利,陈灵芝.北京东灵山地区植物群落多样性的研究 Ⅱ.丰富度、均匀度和物种多样性指数.生态学报,1995,15(3):268-277.
[34] 郝占庆,于德永,吴钢,邓红兵,姜萍,王庆礼.长白山北坡植物群落β多样性分析.生态学报,2001,21(12):2018-2022.
[37] 卢训令,胡楠,丁圣彦,范玉龙,廖秉华,翟元杰,柳静.伏牛山自然保护区物种多样性分布格局.生态学报,2010,30(21):5790-5798.
