干旱和坡向互作对栓皮栎和侧柏生长的影响
2013-09-07冯锦霞王双霞贾长荣万贤崇
王 林,冯锦霞,王双霞,贾长荣,万贤崇,*
(1.中国林业科学研究院新技术所,北京 100091;2.河南省济源市气象局,济源 454650;3.济源市国有大沟河林场,济源 454650)
有许多地区,虽然从年平均降雨或水分平衡来评价都不属于干旱地区,但雨量的不均也会对植物造成严重的危害[1],季节性缺少降雨对树木所造成的干旱胁迫程度与当地的蒸散强度和土壤储水能力有很大的关系,然而,这方面的研究尚不多见。不同的地形特征会影响到环境条件、树种组成和森林生产力。坡向是一个重要的地形影响因素[2],不同坡向由于接受的太阳辐射量不同,在很大程度上影响到温度、空气湿度和土壤水分状况[3]。阳坡因为太阳辐射量更大,与阴坡相比会表现出林内气温高、空气湿度小、土壤水分含量低。林内微气候条件的差异会影响到地表特性和树木的生长速率。阳坡有充足的光照,在水分充足的条件下,阳坡更适合植物生长,尤其是先锋树种和阳性植物,而在干旱地区或干旱时期,阳坡的蒸散失水更严重,土壤干旱加剧,不利于植物生长。不同坡向导致的这些环境条件的差异会造成不同树木在不同坡向生长的不同[2,4]。华北石质山区,土壤瘠薄,生态脆弱,研究这一地区降雨和坡向的交互作用对该地区的生态保护和森林培育具有理论意义。对不同坡向植物的生长状况和水分状况进行研究对不同地形条件下造林树种的选择具有重要的指导意义。
树木年轮学被广泛的用来研究树木与环境之间的长期关系[5-9]。树木年轮宽度和气象条件,尤其是降雨和温度,有着密切的关系[1-2,10]。叶片碳稳定同位素组成(δ13C值)能反映当前时间尺度内气孔导度、叶片内外二氧化碳浓度状况,可以作为水分利用效率(WUE)的指标以及比较不同树木应对干旱的反应[11-12]。植物在轻度和中度干旱胁迫下,WUE往往与受到干旱胁迫的程度呈正相关[13-14]。树木年轮学和碳稳定同位素方法结合能够反映树木生长状况、水分状况,以及它们应对干旱的响应策略[15-16],是分析水分状态和树木生长关系的有效途径[17-18]。
栓皮栎(Quercus variabilis)、侧柏(Platycladus orientalis)是我国的华北石质山区广泛应用的造林树种,这两个树种在我国华北地区都有较强的适应性。但近年人们发现阴坡栓皮栎生长状况明显好于阳坡,反应在树木的大小和森林密度;侧柏也有同样的趋势,但差别没有栓皮栎那么明显。这两个树种有着不同的抗旱策略[19],这可能是造成差别响应的原因。造成不同坡向生长状况差异的主要原因是水分因素,两种树木在两坡向之间的差别与两种树木的抗旱能力和抗旱策略相关。本研究通过对不同坡向栓皮栎和侧柏的年轮宽度和叶片δ13C值进行测定,并结合降雨量、林内微气象和土壤水分状况比较分析两个树种对水分条件的差别响应。目的是探索在华北石质山区水分条件对栓皮栎和侧柏生长发育的影响,分析土壤对这两个树种不同森林密度的承载能力。本研究的假说是:(1)不同坡向不同的蒸散强度造成阳坡和阴坡林内微气象及土壤水分状况的差异,(2)由于土壤水分承载能力的限制,旱季降水量是决定树木生长的主要因子,(3)不同的耐旱能力及应对干旱的策略导致这两个树种在生长上对坡向的差别响应。
1 材料与方法
1.1 试验地概况
试验地位于河南省济源市境内的黄河小浪底森林生态系统定位研究站(35°01'N,112°28'E),地处黄河中游,紧连太行山,属暖温带大陆性季风气候。属低山丘陵区,山体石质,土壤瘠薄。试验地中心区域海拔为410 m左右,土壤类型为棕壤和石灰岩风化母质淋溶性褐土。年平均气温12.4—14.3℃,全年日照时数为2367.7 h,年日照率为54%。年平均降水量641.7 mm,由于受季风气候的影响,雨量分布不均匀。6—9月平均降水量为438.0 mm,占全年的68.3%,冬季及生长季前期常出现严重的季节性干旱[20-21]。
试验地选用生长在丘陵上的栓皮栎和侧柏纯林,坡度为15—25°,于2011年分别在栓皮栎林和侧柏林阴坡和阳坡设置3个20 m×20 m的样地,样地位于坡中部,土层厚度和森林密度见(表1,表2),栓皮栎年龄在30—35a,侧柏树龄为30a左右,林下有少量扁担木(Greeia biloba)荆条(Vitex negundo)等灌木。每个样地随机测定8—10棵树的胸径和树高。

表1 土层厚度Table1 The thickness of soil/cm

表2 植株密度Table2 The plant density of plots
1.3 木芯采集和年轮宽度及其面积测定
在每个样地中选择平均木4株,每株在胸径处(1.3 m)取东西方向和南北方向木芯各1根,用硅胶将木芯固定在木槽中,在实验室中晾干,用不同径级的砂纸按从粗到细的顺序逐级打磨,直到表面光滑平整,年轮间界限清楚,利用LINTAB年轮分析仪测量年轮的宽度(精确到0.01 mm)。将得到的年轮宽度序列用COFECHA软件交叉定年[22],每个单株4个方向相同年份的年轮宽度取平均值。由于本研究的目的主要是比较两个树种在不同生长环境下的生长状况,所以未对年轮宽度进行标准化处理。
每一个单株的两个木芯共4个方向的年轮宽度取平均值作为当年年轮宽度,自树芯最内一年起至当年的总年轮宽度作为半径计算树干截面积,某年的树干截面积减去前一年树干截面积即为某年的树干面积的年增长量。
1.4 土壤含水量测定
土壤含水量用便携式土壤水分测定仪(TDR,Time-Domain Reflectometry)(TRME-T3,MKO公司,德国)测定,测定时间分别为2011年5月底(旱季)、8月初(雨季)。测量深度为20 cm左右。
1.5 林内微气象观测
在2012年6月上旬晴天在两个林分的阴阳坡用手持式温湿度检测仪检测林分内的温度和湿度,6:00—20:00期间每两个小时监测1次,探头放置高度为1.4 m。用温度和湿度计算饱和蒸汽压亏缺,公式如下:
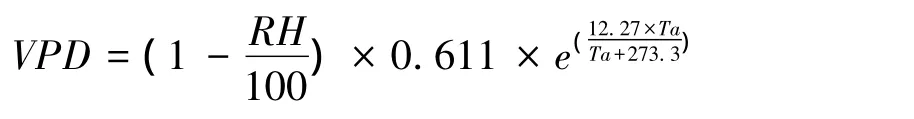
式中,VPD为饱和蒸汽压亏缺(kPa);RH为相对湿度(%),Ta为空气温度(℃)。
由于2个不同林分在相同坡向的林内微气象几乎没有差别,取2个林分相同坡向数据的平均值作为坡向林内微气候数据。
1.6 叶片碳同位素测定
于2011年5月底(旱季),采集树冠中上部向阳面、完全展开的叶片用于碳稳定同位素测定。采后叶片立刻带回实验室洗净、烘干,用粉碎机粉碎,过80目筛。碳稳定同位素测定在中国林业科学院稳定同位素实验室进行,用Flash EA1112元素分析仪-Conflo III-Delta Plus同位素质谱仪Isotope Ratio Mass Spectrometer(DELTA V Advantage Thermo Fisher Scientific,Inc.,USA),分析精度±0.1‰。碳稳定同位素组成采用以下公式计算 δ13C =((13C/12Csample)/(13C/12Cstandard)-1)×1000。13C/12Cstandard是国际标准物(PDB,Pee Dee Belnite)[23]。
1.7 统计分析
用Excel软件进行方差分析、t-检验和相关分析。以P<0.05作为显著性的标准。
2 结果与分析
2.1 不同坡向两个树种的大小和年轮宽度
阳坡栓皮栎的胸径和树高均显著低于阴坡(P<0.05),阳坡栓皮栎胸径是阴坡的86.2%,树高是阴坡的95.8%。侧柏在两种坡向之间也有显著差别,阳坡侧柏胸径是阴坡的92.6%(P=0.032),树高是阴坡的95.0%(P=0.036)(图 1)。
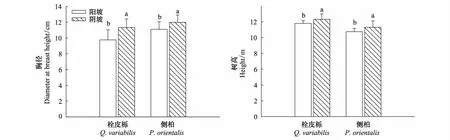
图1 栓皮栎、侧柏的树高和胸径Fig.1 Height and diameter at breast height of Q.variabilis and P.orientalis
栓皮栎阴坡和阳坡平均年轮宽度分别为1.41 mm和1.16 mm,阴坡阳坡侧柏的年轮宽度分别为1.57 mm和1.46 mm(图2)。运用成对双样本t检验显示阳坡栓皮栎和侧柏均显著(P<0.05)低于阴坡,阳坡栓皮栎比阴坡低11.6%,阳坡侧柏比阴坡低7.5%。栓皮栎和侧柏年轮宽度都表现出随着树龄的增加年轮宽度减小的趋势,但变化模式不同(图2)。栓皮栎年轮宽度变异幅度很大,在1993年以后年轮宽度急剧减小,而侧柏年轮宽度随时间的变化相对平缓(图2)。每年增加的截面积和年轮宽度变化结构不同(图3)。栓皮栎在1985年以前年截面积增长量直线上升,之后一直保持相对平稳的生长趋势,阴坡上栓皮栎明显高于阳坡,尤其是最近8—9a(图3)。侧柏在2006年以前,一直保持平稳上升的增长趋势,在有些年份,阴坡的略高于阳坡(图3)。2006年以后两个树种截面积在所有坡向都出现下降。运用成对双样本t检验显示阳坡栓皮栎和侧柏年增加截面积均显著(P<0.05)低于阴坡,分别低34.4%和13.8%。
2.2 叶片碳稳定同位素
栓皮栎和侧柏都表现出阳坡叶片δ13C值显著高于阴坡(P<0.05)。栓皮栎在两种坡向之间的δ13C值差别为1.17,侧柏为0.56,栓皮栎两坡向之间的δ13C值差别大于侧柏(图4)。
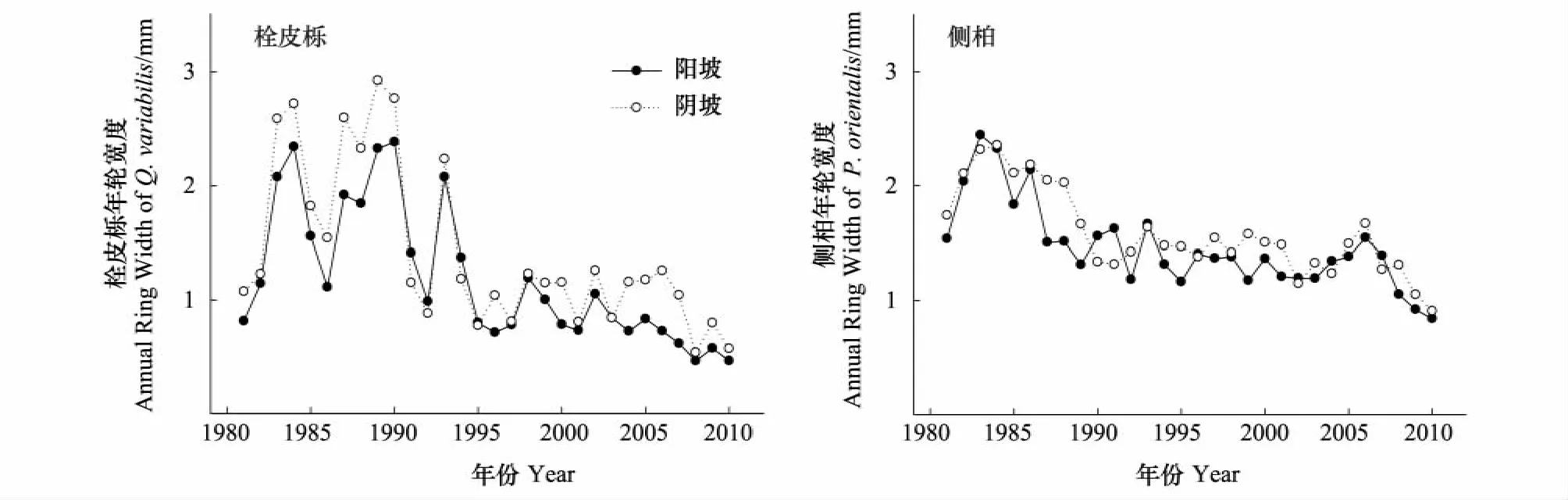
图2 不同坡向栓皮栎与侧柏年轮宽度Fig.2 The annual ring width of Q.variabilis and P.orientalis on different aspects
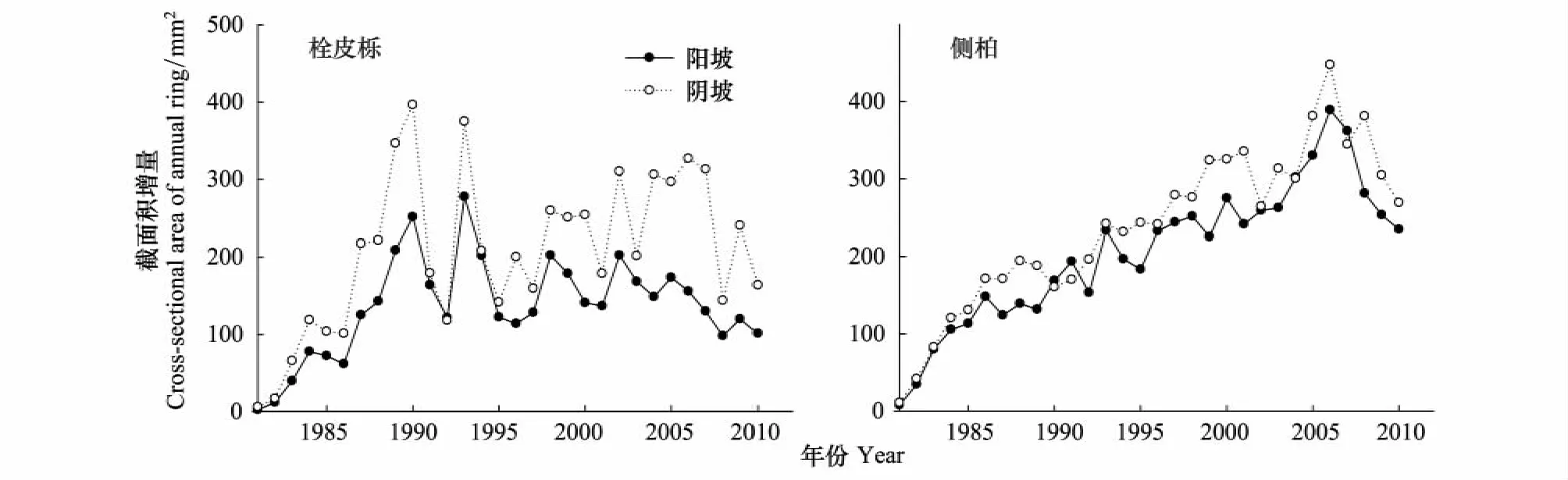
图3 不同坡向栓皮栎与侧柏年轮截面积Fig.3 The cross-sectional area of annual ring of Q.variabilis and P.orientalis on different aspects
2.3 气象数据和林内微气象
1980—2010年温度和降水数据见(图5),观测点距离试验地20 km左右。
在10:00—18:00期间,阳坡林内气温明显高于阴坡,湿度低于阴坡,饱和蒸汽压亏缺高于阴坡。两坡向之间最大温差、最大空气湿度差和最大饱和蒸汽压亏缺差值都出现在14:00,此时阳坡温度比阴坡高1.5℃,相对湿度比阴坡低1.6%。饱和蒸汽压亏缺比阴坡高4.1 kPa,阴坡温度在凌晨和傍晚略高于阳坡(图6)。
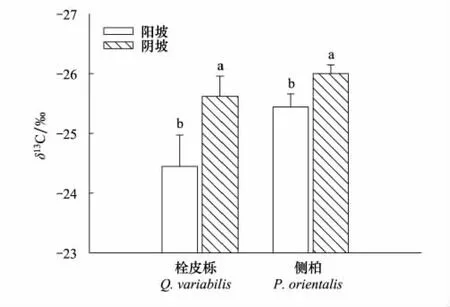
图4 不同坡向的栓皮栎、侧柏叶片δ13 C值Fig.4 The leaf δ13 C of Q.variabilis and P.orientalis on different aspects
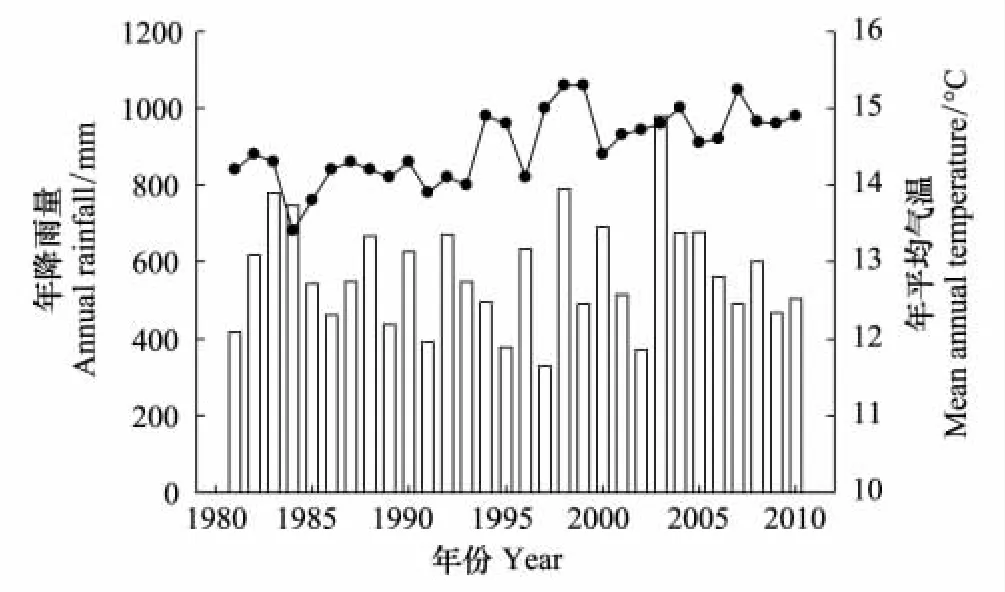
图5 1981—2010年的年降水量和平均气温Fig.5 Annual rainfall and mean annual temperature from 1981 to 2010
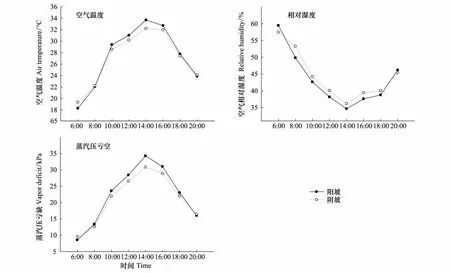
图6 不同坡向林地在6月上旬晴天的空气温度、相对湿度、蒸汽压亏空的日变化Fig.6 The diurnal variation of air temperature,relative humidity,and vapor deficit on sunny days in early June
2.4 土壤含水量
两个林分不同坡向雨季土壤水分含量都大约是旱季的2倍。在旱季,阳坡土壤含水量显著(P<0.05)低于阴坡,在雨季阳坡土壤含水量也略低于阴坡,但差异不显著(P=0.08)。对两个树种的林地进行比较,在旱季,侧柏林土壤含水量低于栓皮栎林(P<0.05),而在雨季两者之间没有差别(P=0.3)(图7)。
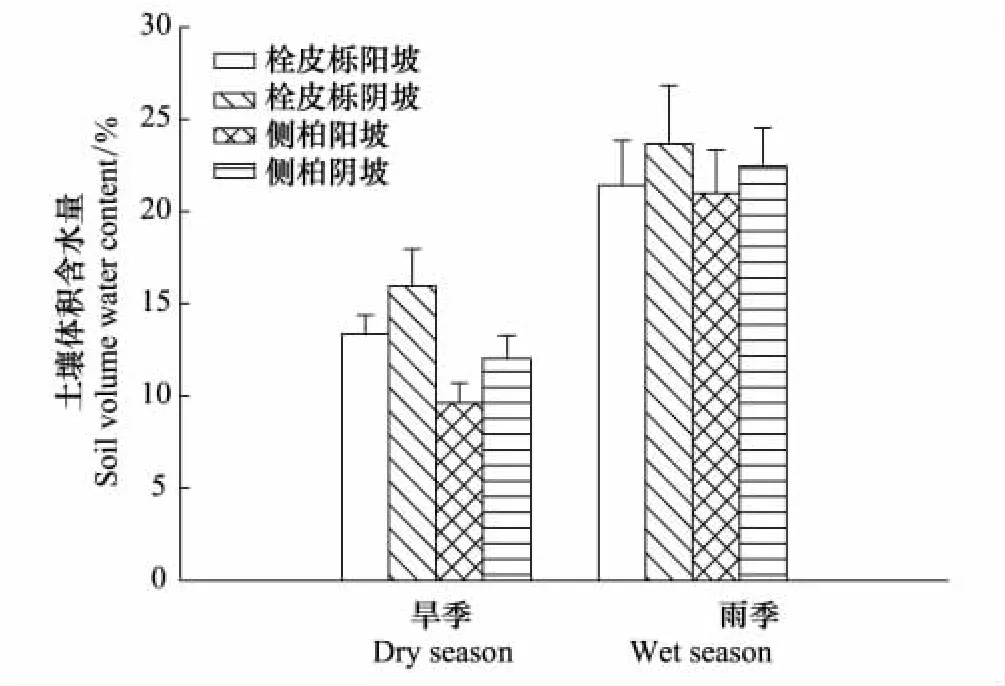
图7 栓皮栎、侧柏林在旱季和雨季土壤含水量Fig.7 Soil volume water content of Q.variabilis and P.orientalis in dry season and wet season
2.5 降雨量和生长的关系
本文分别用全年降雨量、雨季降雨量或旱季降雨量和树干截面积年增长量、年轮宽度进行了相关分析,结果显示只有旱季降雨量和年增长面积呈现差异相关性(表3)。栓皮栎的年增长面积和旱季降雨量呈显著正相关,尤其是阳坡的栓皮栎,其相关系数达0.7。而侧柏年增长面积和旱季降雨量没有显著关系,尽管阴坡的年增长面积侧柏与旱季降雨量呈现一定的正相关趋势。无论是全年、雨季还是旱季降雨量和年轮宽度都没有相关性,全年或雨季降雨量和树干截面积年增长也没有显著相关。

表3 旱季(11月—5月)降雨量与树干截面积年增长量的相关性Table3 The correlations between the cross-sectional area of annual ring and precipitation from November to May
3 讨论
华北丘陵低山的坡向不足以改变降雨,水分平衡主要可能受蒸散的影响。另外,在季节性干旱的森林生态系统,土壤厚度也是影响森林生长力的重要因素,因为土壤厚度影响根系生长量、土壤蓄水能力[24-25]。两坡向之间林内微气象的差异主要与阳坡接收太阳辐射量差异有关,阳坡的净太阳辐射明显超过阴坡[26]。导致阳坡的温度较高,相应的饱和蒸汽压亏缺更大,尤其在气孔张开的白天。这些现象和以前的报道是一致的[2,27]。饱和蒸汽压亏缺直接影响到蒸腾速率和光合速率[28],所以坡向所导致的微气候差别会对土壤水分和树木生长造成不同的影响。在干旱季节,阳坡的土壤含水量显著低于阴坡。另外,还注意到,在旱季阳坡侧柏林的土壤含水量低于栓皮栎林。在这个地区,旱季跨过整个冬季,常绿树种在当温度适宜时保持着蒸腾,所以对水分的消耗较落叶林多,后者在发叶之前,蒸腾量很小。另外,侧柏林密度比栓皮栎大,也是加大耗水的一个因素。
δ13C值和植物的水分利用效率成正比。当外界CO2浓度和叶温没有显著差异时,主要取决于胞间CO2浓度,换句话说也就是气孔导度和叶肉导度,以及羧化酶对12C的选择性[23,29],这些指标都反映了树木受干旱胁迫的程度。另外,适度干旱条件会提高植物的水分利用效率[30]。这表明阳坡栓皮栎和侧柏生长受抑主要是由于较严重的水分亏欠。值得注意的是,这两个树种的δ13C值,即水分利用效率,和生产力没有正的相关关系,亦即和它们的抗干旱能力没有显著的关系,这种现象曾有过报道[29,31],他们认为在有些情况δ13C值只反应受干旱胁迫的程度和植物的生产力没有关系。还有,无论从直径生长还是从δ13C值所反映的受干旱胁迫程度,坡向对栓皮栎的影响都显著大于侧柏。这可能和侧柏与栓皮栎本身的特性相关,先前我们用气体交换仪测定发现在正常供水和干旱条件下栓皮栎的瞬时水分利用效率都高于侧柏[19]。侧柏有更大的有效根长密度[32-33],更大的有效根长密度预示着有更强的根系吸水能力。
试验地区土壤脊薄,土壤的储水能力有限,根系接触到地下水的可能性很小,植物的生长主要取决于降水,所以季节性干旱的影响很突出。从相关分析来看,旱季的降雨量对栓皮栎直径生长起到决定性的作用(表3),但侧柏的直径生长不完全取决于旱季的降雨。我们先前的研究发现侧柏的气孔对干旱胁迫的敏感性较高[19],通过关闭气孔来减少水分损失、缓解干旱胁迫,但同时也导致了光合速率下降[34]。可以推测侧柏由于气孔关闭减少了光合积累,导致其在短期的干旱条件下生长缓慢,但通过关闭气孔可以应对严重的干旱。侧柏气孔相对较高的敏感性使其在雨季短暂的缺少降雨的情况下也可能引起气孔关闭,减少光合积累。而在旱季,侧柏具有较强的耐旱能力,其需水较少,在很干旱时仍然可以保持一定水平的光合作用[19],侧柏的这些特性可以解释为什么旱季的降雨量和它的直径生长没有密切的相关性。另外,植物也可能通过各器官再分配来适应外界环境。与此不同,栓皮栎是落叶树种,冬季的干旱应该对它的影响较小,本文结果可见,旱季中生长季早期的干旱对栓皮栎的增粗生长有很严重的影响,这和增粗生长主要受当年的环境条件调节的学说是一致的[35]。在华北石质山区,树木的生长主要受干旱的制约。但从全年的降雨量来看,该地区并不属于干旱或半干旱地区。所以全年的降雨量和树干截面积年增长并没有显著相关。另外,雨季的雨量充沛,有时还过剩,所以不是制约生长的因子。
随着树干直径不断增大,同样宽度的年轮增长,意味着更大的树干截面积或生物量的增长,在本试验地区,水肥条件所限栓皮栎和侧柏都不能保持均速生长,我们发现年轮宽度随树龄有下降的趋势,尤其是年龄较大时这种现象更明显。所以年轮宽度和降雨量没有呈现显著相关性。
4 结论
石质山区土壤脊薄,其承载能力主要取决于降雨量和降雨的均匀度。在华北石质丘陵地带,阳坡具有更大蒸散作用,其与旱季交互作用导致土壤干旱,抑制了栓皮栎和侧柏的生长。这两种树种相比较侧柏具有更强的抗旱能力。旱季的降雨量制约了栓皮栎的生长,而侧柏直径生长和旱季降雨量没有直接的关系。
[1] Villalba R,Grau H R,Boninsegna J A,Jacoby G C,Ripalta A.Tree-ring evidence for long-term precipitation changes in subtropical South America.International Journal of Climatology,1998,18(13):1463-1478.
[2] Fekedulegn D,Hicks R R,Colbert J.Influence of topographic aspect,precipitation and drought on radial growth of four major tree species in an Appalachian watershed.Forest Ecology and Management,2003,177(1/3):409-425.
[3] Rosenberg N J,Blad B L,Verma SB.Microclimate:The Biological Environment.New York:John Wiley& sons Inc,1983.
[4] Abrams M D,Ruffner C M,Morgan T A.Note:tree-ring responses to drought across species and contrasting sites in the ridge and valley of central Pennsylvania.Forest Science,1998,44(4):550-558.
[5] Cullen L E,Palmer JG,Duncan R P,Stewart GH.Climate change and tree-ring relationships of Nothofagusmenziesii tree-line forests.Canadian Journal of Forest Research,2001,31(11):1981-1991.
[6] Lo Y H,Blanco J A,Seely B,Welham C,Kimmins J P.Relationships between climate and tree radial growth in interior British Columbia,Canada.Forest Ecology and Management,2010,259(5):932-942.
[7] Peterson D W,Peterson D L.Mountain hemlock growth responds to climatic variability at annual and decadal time scales.Ecology,2001,82(12):3330-3345.
[8] Zhang W T,Jiang Y,Dong M Y,Kang M Y,Yang H C.Relationship between the radial growth of Picea meyeri and climate alongelevations of the Luyashan Mountain in North-Central China.Forest Ecology and Management,2012,265:142-149.
[9] Zhang Y X,Wilmking M,Gou X H.Changing relationships between tree growth and climate in Northwest China.Plant Ecology,2009,201(1):39-50.
[10] Liang E Y,Shao X M,Kong Z C,Lin JX.The extreme drought in the 1920s and its effect on tree growth deduced from tree ring analysis:a case study in North China.Annals of Forest Science,2003,60(2):145-162.
[11] Pate J S.Carbon isotope discrimination and plant water-use efficiency//Unkovich M,Pate J,eds.Stable Isotope Techniques in the Study of Biological Processes and Functioning of Ecosystems.Boston:Kluwer,2001:19-36.
[12] Brienen R JW,Wanek W,Hietz P.Stable carbon isotopes in tree rings indicate improved water use efficiency and drought responses of a tropical dry forest tree species.Trees-Structure and Function,2011,25(1):103-113.
[13] Anyia A O,Slaski J J,Nyachiro J M,Archambault D J,Juskiw P.Relationship of carbon isotope discrimination to water use efficiency and productivity of barley under field and greenhouse conditions.Journal of Agronomy and Crop Science,2007,193(5):313-323.
[14] Cui N B,Du T S,Kang S Z,Li F H,Hu X T,Wang M X,Li Z J.Relationship between stable carbon isotope discrimination and water use efficiency under regulated deficit irrigation of pear-jujube tree.Agricultural Water Management,2009,96(11):1615-1622.
[15] Adams H D,Kolb T E.Drought responses of conifers in ecotone forests of northern Arizona:tree ring growth and leafδ13C.Oecologia,2004,140(2):217-225.
[16] Williams A P,Michaelsen J,Leavitt SW,Still C J.Using tree rings to predict the response of tree growth to climate change in the continental United States during the twenty-first century.Earth Interactions,2010,14(19):1-20.
[17] Ferrio JP,Florit A,Vega A,Serrano L,Voltas J.δ13Cand tree-ring width reflect different drought responses in Quercusilex and Pinus halepensis.Oecologia,2003,137(4):512-518.
[18] Vaganov E A,Schulze E D,Skomarkova M V,Knohl A,Brand W A,Roscher C.Intra-annual variability of anatomical structure andδ13Cvalues within tree rings of spruce and pine in alpine,temperate and boreal Europe.Oecologia,2009,161(4):729-745.
[19] Jin X,Xu J,Bai K D,Feng J X,Zhang J S,Wan X C.Comparison of drought strategies of three co-existing woody plants by their hydraulic structures.Journal of Beijing Forestry University,2011,33(6):136-141.
[20] Sun SJ,Meng P,Zhang J S,Wan X C.Variation in soil water uptake and its effect on plant water status in Juglans regia L.during dry and wet seasons.Tree Physiology,2011,31(12):1378-1389.
[21] Zhang J S,Men P,Wang H S,Gao J,Ren Q F,Jia CR,Ren Y F.Soil respiration of Robinia pseudoacacia plantation in the Rocky Mountainous area of North China.Scientia Silvae Sinicae,2008,44(2):8-14.
[22] Holmes R L.Computer-assisted quality control in tree-ring dating and measurement.Tree-Ring Bulletin,1983,43(1):69-78.
[23] Farquhar G D,Ehleringer J R,Hubick K T.Carbon isotope discrimination and photosynthesis.Annual Review of Plant Physiology and Plant Molecular Biology,1989,40(1):503-537.
[24] Poff R J.Effects of silvicultural practices and wildfire on productivity of forest soils//Sierra Nevada Ecosystem Project:Final Report to Congress,vol.II:Assessments and Scientific Basis for Management Options.Davis:University of California,Centers for Water and Wildlands Resources,1996:477-495.
[25] Buttle JM,Dillon P J,Eerkes G R.Hydrologic coupling of slopes,riparian zones and streams:an example from the Canadian Shield.Journal of Hydrology,2004,287(1/4):161-177.
[26] Lee R,Sypolt C R.Toward a biophysical evaluation of forest site potential.Forest Science,1974,20(2):145-154.
[27] Liang E Y,Shao X M,Eckstein D,Huang L,Liu X H.Topography-and species-dependent growth responses of Sabina przewalskii and Picea crassifolia to climate on the northeast Tibetan Plateau.Forest Ecology and Management,2006,236(2/3):268-277.
[28] Kramer P J,Kozlowski T T.Physiology of Trees.New York:McGraw-Hill,1960.
[29] Monclus R,Dreyer E,Villar M,Delmotte F M,Delay D,Petit J M,Barbaroux C,Le Thiec D,Brechet C,Brignolas F.Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides×Populus nigra.New Phytologist,2006,169(4):765-777.
[30] Rouhi V,Samson R,Lemeur R,Van Damme P.Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery.Environmental and Experimental Botany,2007,59(2):117-129.
[31] Condon A G,Richards R A,Rebetzke G J,Farquhar G D.Breeding for high water-use efficiency.Journal of Experimental Botany,2004,55(407):2447-2460
[32] Xiang SQ,Zhao X H.Root systems of main afforestation tree species in Beijing.Journal of Beijing Forestry University,1981,(2):19-32.
[33] Wang JX,Wang DH,Liu GQ.Distribution characteristics of effective root density in the planted Robinia pseudoacacia and Platycladus orientalis forest site.Acta Botanica Boreali-Occidentalia Sinica,2004,24(13):2208-2214.
[34] McDowell N G,Pockman WT,Allen CD,Breshears DD,Cobb N,Kolb T,Plaut J,Sperry J,West A,Williams DG,Yepez EA.Mechanisms of plant survival and mortality during drought:why do some plants survive while others succumb to drought?New Phytologist,2008,178(4):719-739.
[35] Kozlowski T T,Pallardy SG.Growth Control in Woody Plants.San Diego:Academic Press,1997.
参考文献:
[19] 靳欣,徐洁,白坤栋,冯锦霞,张劲松,万贤崇.从水力结构比较3种共存木本植物的抗旱策略.北京林业大学学报,2011,33(6):136-141.
[21] 张劲松,孟平,王鹤松,高峻,任庆福,贾长荣,任迎丰.华北石质山区刺槐人工林的土壤呼吸.林业科学,2008,44(2):8-14.
[32] 向师庆,赵相华.北京主要造林树种的根系研究.北京林业大学学报,1981,(2):19-32.
[33] 王进鑫,王迪海,刘广全.刺槐和侧柏人工林有效根系密度分布规律研究.西北植物学报,2004,24(13):2208-2214.
