太湖草、藻型湖区沉积物-水界面厚度及环境效应研究
2013-09-07王永平朱广伟洪大林秦伯强南京水利科学研究院水文水资源与水利工程科学国家重点实验室江苏南京009中国科学院南京地理与湖泊所湖泊与环境国家重点实验室江苏南京0008
王永平 ,朱广伟 ,洪大林 ,秦伯强 * (.南京水利科学研究院,水文水资源与水利工程科学国家重点实验室,江苏 南京 009;.中国科学院南京地理与湖泊所,湖泊与环境国家重点实验室,江苏 南京 0008)
沉积物-水界面是天然水体在物理、化学和生物特征等方面差异性最显著和负责水体和沉积物之间物质输送和交换的重要边界环境.沉积物-水界面的生物地球化学过程就是指发生在界面附近的物理和化学反应,包括氧化和还原、溶解和沉淀、吸附和解吸、迁移和转化、扩散和埋藏、细菌生化反应及生物扰动等作用[1].因而沉积物-水界面是水环境地球化学循环和生物系统耦合的重要方面,是控制和调节水体和沉积物之间物质输送和交换的重要途径,也被认为是影响浅水湖泊内源释放的重要因素.
目前,许多学者围绕着浅水湖泊中沉积物-水界面的物质(包括营养盐、重金属以及浮游藻类等)释放通量[2-6]、释放通量与界面环境条件的关系[7]、营养盐在水-沉积物界面上迁移的过程[8]、沉积物再悬浮对磷释放的影响[9-10]、环境因子(如:水中溶解氧、pH值、氧化还原电位、温度、生物及水体的扰动等)对沉积物中营养盐释放影响等[6,11-13]等开展了大量的研究.但是,大部分研究都将沉积物-水界面当成“黑匣子”,仅研究其功能,而对于沉积物-水界面的厚度及其结构的研究甚少,且所得结论差距较大.吴丰昌[14]等认为,沉积物-水界面是指新近沉降的沉积物(15cm左右)与水的界面,并且存在扩散亚层――沉积物最表面水体组成的边界层,是沉积物表面粘滞流层的底层,对界面的物质交换和早期成岩作用的方式和强度等有重要的价值,但只能通过间接估计方法得出其厚度.高光等在对太湖沉积物中微生物的研究中发现,在3~5和10cm 左右,无论是微生物的种类还是数量,都出现了峰值(待发表数据).王雨春等[2]和杨龙元等[15]研究认为,营养盐的分解及释放通常集中在沉积物-水界面0~2cm的薄层内.Morris等[16]模拟了沉积物剖面上有机质的矿化,认为有机质矿化作用主要发生在表层2cm厚的沉积物中,2cm以下沉积物中有机质的矿化速度小的几乎可以忽略.而最近一些研究发现:具有极高生物地球化学活性的沉积物-水界面通常在mm范围内[17].
因此,本试验拟通过对太湖沉积物-水界面的结构的调查比较,以期为了解沉积物-水界面厚度和进一步开展沉积物-水界面微环境结构的研究打下基础.
1 材料与方法
2010年9月中旬,分别在太湖贡湖湾与梅梁湾交叉口(藻型湖区)和胥口湾(草型湖区)采样.每个样点用自制的分层采水器采集泥面以上5,20,35cm处的水样以及水面以下20cm处水样,一部分水样现场过0.45um GF/F膜,滤后水放入车载冰箱冷藏,剩余水样带回实验室.同时用柱状采样器采集界面清晰的泥柱 3根,密封后运回实验室,用溶氧微电极(PreSens, German)测定沉积物表层溶解氧(DO)剖面后,再从上往下按 1,2,2,5,5cm间隔取5层泥样.
滤后水用 Skalar流动分析仪(荷兰)测定硝氮、亚硝氮、氨氮和磷酸根,用1020型TOC仪(O.I. Corporation, American)测定溶解性有机碳(DOC)浓度,原水参照《湖泊富营养化调查规范》[18]测定 TN、TP、TDN、TDP、Chl-a、总悬浮物浓度(SS总)和有机悬浮物浓度(SS有机)[19].
泥样采用烘干法测定其含水量与孔隙率[20],重铬酸钾-硫酸消化-凯氏定氮法测定 TN,高氯酸-硫酸酸溶-钼锑抗比色法测定TP,重铬酸钾-硫酸(油浴)氧化-硫酸亚铁滴定法测定总有机碳(TOC).用于粒径分析的样品用稀盐酸、双氧水处理,分别去除碳酸盐和有机质后,用Mastersizer-2000型激光粒径仪(Malvern,English)进行分析,样品分析误差小于5%.
2 结果与分析
2.1 草、藻型湖区各水层指标差异
草、藻型湖区各水层指标如图1所示.草型湖区中SS总、SS有机、Chl-a、TN、TDN、TP和TDP等指标显著低于藻型湖区.其中,除Chl-a和TDP浓度在各层间没有显著差异外,草型湖区水层中 SS总、SS有机、TN、TDN和TP都大致呈现出越往下浓度越高的趋势.在藻型湖区水层中,由于风浪扰动频繁导致水柱混合,各水层间的SS总、SS有机、TN、TDN和TP都没有显著差异.其中, Chl-a在表层水体中显著高于其他各层,主要原因是蓝藻上浮并在水表面富集.
草、藻型湖区间以及各水层间的DOC、磷酸根和氨氮浓度的规律性不是很明显,差异也不显著.其中,仅有草型湖区水柱的磷酸根浓度同样受沉积物静态内源释放的影响,基本呈现出越往下浓度越高的趋势.另外,硝态氮和亚硝态氮在藻型湖区水 柱中含有微量,而在草型湖区中基本检测不出.
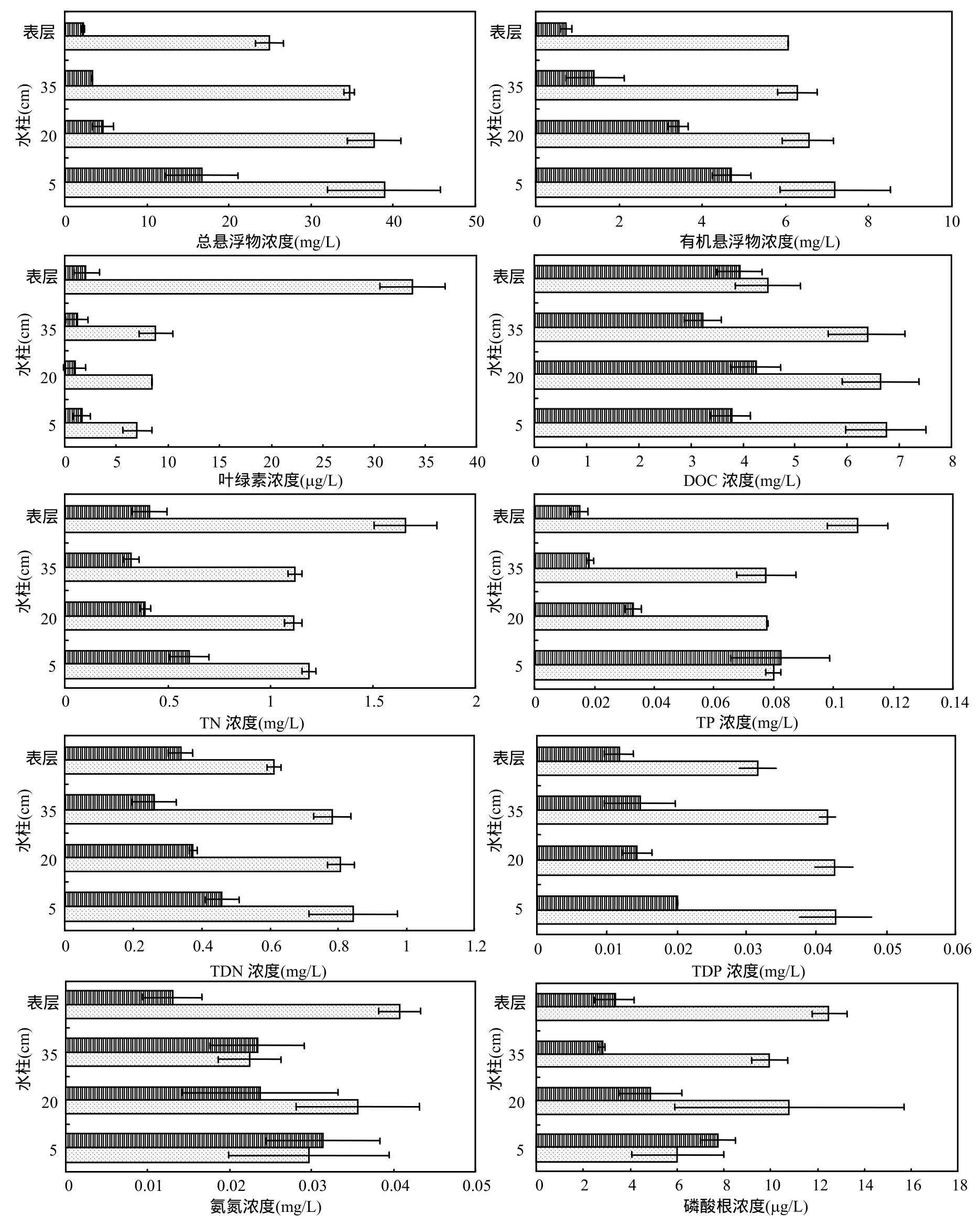
图1 太湖草、藻型湖区水层各项指标差异Fig.1 Parameters profile in water columns of phytoplankton-dominant and macrophyte-dominant regions
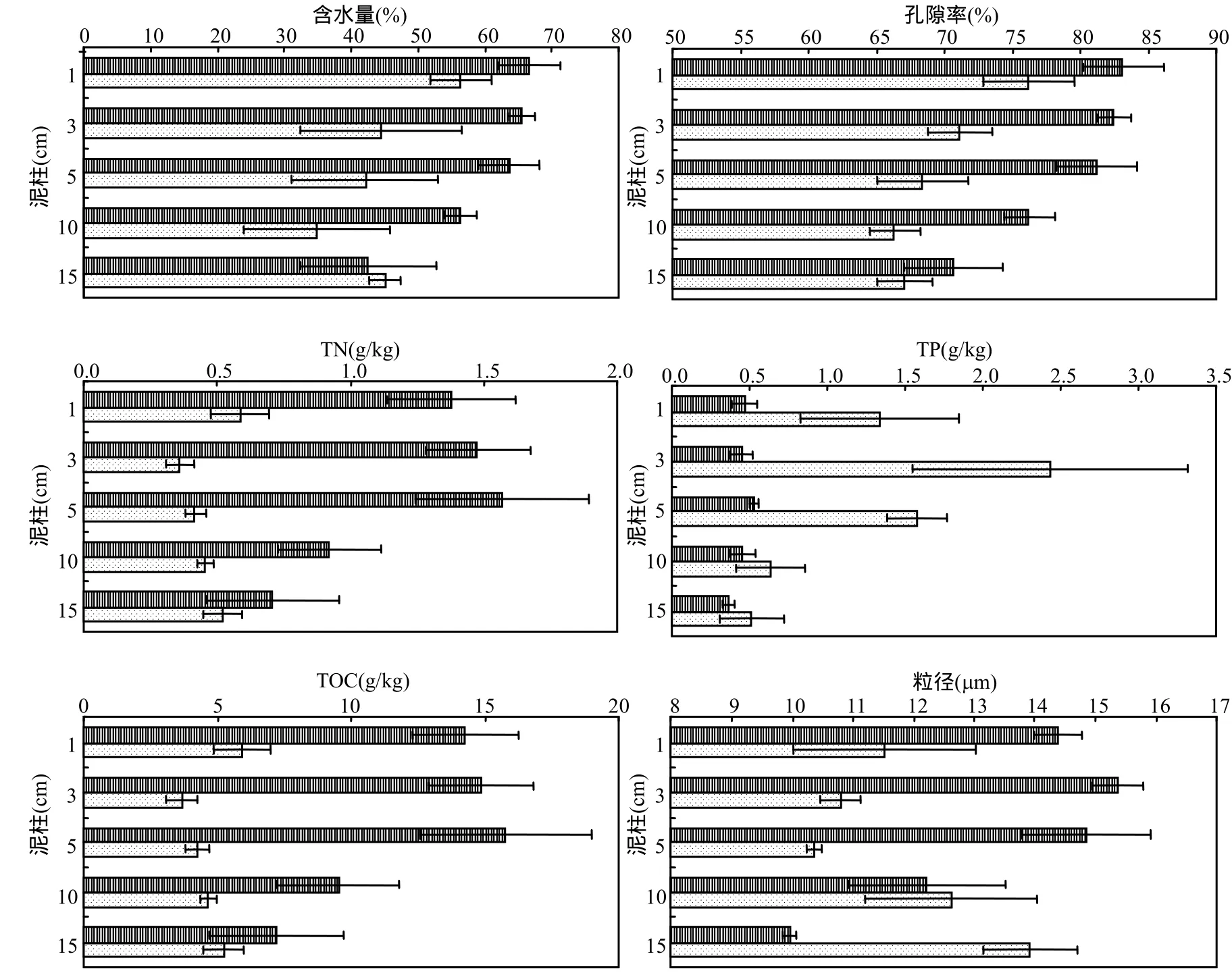
图2 太湖草、藻型湖区沉积物层各项指标差异Fig.2 Parameters profile in sediment cores of phytoplankton-dominant and macrophyte-dominant regions
2.2 草、藻型湖区沉积物层各指标变化

图3 太湖草、藻型湖区表层沉积物溶解氧含量Fig.3 Oxygen profiles in sediment cores in phytoplankton-dominant and macrophytedominant regions
草、藻型湖区沉积物各层指标如图 2所示.草、藻型湖区沉积物的含水量与孔隙率都大致呈逐渐递减的趋势,草型稍高于藻型.草型湖区沉积物的TN和TOC显著高于藻型,且都在3~5cm处出现增高或降低的拐点.而沉积物的 TP则表现为藻型湖区显著高于草型湖区,但也在3~5cm处出现了拐点.两类湖区间的沉积物中值粒径没有显著差异,各层均值都在 9~16μm 之间.同样可以观察到,中值粒径在3~5cm处发生增大或变小的改变.
图3是由溶氧微电极测得的草、藻型湖区沉积物表层DO浓度剖面,可以看到,草型湖区沉积物表面的溶解氧高于藻型,但随后迅速下降,在泥下1mm处即检测不到溶解氧.而在藻型湖区的沉积物中,溶解氧在泥下0.5~1mm处出现迅速下降的趋势,直到泥下2.5mm处才趋于0,溶氧层厚度明显高于草型.
3 讨论
由于受沉积物分层技术和检测技术的分辨率的限制,对于沉积物-水界面的厚度一直都众说纷纭.本试验结果显示,草、藻型湖区沉积物的TN、TP、TOC和粒径等指标都在3~5cm处出现拐点,意味着 3~5cm 处上下沉积物层的特异性.而沉积物表面溶解氧深度的试验结果表明,DO浓度在沉积物表面以下0.5mm即出现拐点.并且,草型湖区表层1mm和藻型湖区表层2.5mm沉积物由于含氧,与其他各层沉积物也存在着明显的差异性.有观点认为,界面发生的许多生物地球化学反应实际上是一个由有氧/无氧条件变化调节的氧化还原边界层的转化控制的.很多化学物质氧化还原态的改变,也将改变表层沉积物的吸收和释放特征,如有机质的氧化会释放N和P,氧化态的铁和锰将对P等产生吸附等.因此,虽然受不同指标和不同研究手段的影响,沉积物-水界面中沉积物厚度的界定出现了较大的分歧.但是,与沉积物-水界面微环境研究密切相关以及对沉积物营养盐的内源释放影响最大的,很有可能是与溶解氧有关的mm级沉积物表层.
另外,对沉积物—水界面的环境效应研究发现:在水动力扰动频繁的藻型湖区,至少在沉积物表面以上 35cm 内,水体各项指标都没有出现显著差异;而在相对较为静止的草型湖区,水柱中大部分指标都呈现出越往下浓度越高的现象,可见这些指标主要受沉积物静态内源释放的影响.另一方面,藻型湖区由于蓝藻水华的存在,能显著改变水体中pH值并刺激沉积物中溶解性营养盐的释放[21-22]或者降低表层沉积物对磷的滞留能力[23].而水生植物的根际微环境由于根系的泌氧和分泌其他氧化性物质的能力而处于氧化环境,能促进金属离子从还原态转化为氧化态,从而极显著地增加沉积物对磷的吸附,减少磷的解析[24-25].并且,水生植物能通过根系吸收沉积物中生物有效磷,有效减少沉积物间隙水中的磷向上覆水的扩散量,还通过茎叶拦截、吸附水中的颗粒物质并通过颗粒物质吸附水中的溶解性磷,有效的降低上覆水中磷及颗粒物的负荷[26].也有研究表明,沉水植物在存在及生长过程中,沉积物上部5cm沉积物间隙水中氨氮含量逐步降低,表现出沉积物-水界面氮的释放通量与沉积物生物量存在负相关性[27].即使根系不发达的黑藻,也能显著降低上覆水中总磷、溶解性活性磷和溶解性总磷的浓度[28].这些研究结果与本试验结果中草型湖区水柱的 SS总、SS有机、Chl-a、TN、TDN、TP和TDP等指标显著低于藻型湖区一致.因此,不同的生境条件对水-沉积物界面的水层厚度也有明显的影响.
4 结论
4.1 草型湖区中 SS总、SS有机、Chl-a、TN、TDN、TP和 TDP等指标显著低于藻型湖区.草型湖区水柱中SS总、SS有机、TN、TDN和TP等指标都呈现出越往下浓度越高的趋势,而藻型湖区各水层间差异不明显.
4.2 草型湖区沉积物的TN和TOC显著高于藻型,TP则显著低于藻型;两个湖区沉积物的TN、TP、TOC和粒径都在3~5cm处出现拐点.
4.3 草型湖区沉积物溶解氧层厚度(<1mm)小于藻型湖区(<2.5mm).
[1]Berner R A. Early diagenesis: a theoretical approach [M].Princeton: Princeton University Press, 1980.
[2]杨龙元,蔡启铭,秦伯强.太湖梅梁湾沉积物-水界面氮迂移特征初步研究 [J]. 湖泊科学, 1998,(10):41-47.
[3]Havens K E, Schelske C L. The importance of considering biological processes when setting total maximum daily loads(TMDL) for phosphorus in shallow lakes and reservoirs [J].Environmental Pollution, 2001,113:1-9.
[4]Petticrew E L, Arocena J M. Evaluation of iron-phosphate as a source of internal lake phosphorus loadings [J]. The Science of the Total Environment, 2001,266:87-93.
[5]Kisand A, Nõges P. Sediment phosphorus release in phytoplankton dominated versus macrophyte dominated shallow lakes: importance of oxygen conditions [J]. Hydrobiologia, 2003,506~509:129-133.
[6]范成新,张 路,秦伯强,等.太湖沉积物-水界面生源要素迁移机制及定量化-Ⅰ. 铵态氮释放速率的空间差异及源-汇通量 [J].湖泊科学, 2004,16(1):10-20.
[7]罗潋葱,秦伯强,朱广伟,等.动力扰动下太湖梅梁湾水-沉积物界面的营养盐释放通量 [J]. 中国科学 D 辑, 2005,35(增刊 II):166-172.
[8]Qin B Q, Zhu G W. The nutrient forms, cycling and exchange flux in the sediment and overlying water system in lakes from the middle and lower reaches of Yangtze River [J]. Science in China:Series D Earth Sciences, 2006,49(Supp. I):1-13.
[9]朱广伟,秦伯强,张 路,等.太湖底泥悬浮中营养盐释放的波浪水槽试验研究 [J]. 湖泊科学, 2005,17(1):61-68.
[10]范成新,张 路,秦伯强,等.风浪作用下太湖悬浮态颗粒物中磷的动态释放估算 [J]. 中国科学D辑, 2003,33(8):760-768.
[11]张 路,朱广伟,罗潋葱,等.风浪作用下太湖梅梁湾水体磷负荷变化与水体氧化还原特征关系 [J]. 中国科学(D辑), 2005,35(增刊II):138-144.
[12]姜 霞,钟立香,王书航,等.巢湖水华暴发期水-沉积物界面溶解性氮形态的变化 [J]. 中国环境科学, 2009,29(11):1158-1163.
[13]张 雷,商景阁,王兆德,等.河蚬扰动沉积物界面效应及其在水中代谢速率 [J]. 中国环境科学, 2011,31(6):1001-1006.
[14]吴丰昌,万国江,蔡玉荣.沉积物-水界面的生物地球化学作用[J]. 地球科学进展, 1996,11(2):191-197.
[15]王雨春,万国江,尹澄清,等.红枫湖、百花湖沉积物全氮、可交换态氮和固定铵赋存特征 [J]. 湖泊科学, 2002,(14):301-309.
[16]Morris J T, Bowden W B. A mechanistic, numerical model of sedimentation, mineralization, and decomposition for marsh sediments [J]. Soil Sci Soc America J, 1986,50(1):96-105.
[17]Viollier E, Rabouille C, Apitz S E, et al. Benthic biogeochemistry:state of the art technologies and guidelines for the future of in situ survey [J]. Journal of Experimental Marine Biology and Ecology,2003,285-286:5-31.
[18]金相灿,屠清瑛.湖泊富营养化调查规范 [M]. 2版,北京:中国环境科学出版社, 1992.
[19]张运林,秦伯强,陈伟民,等.太湖水体中悬浮物研究 [J]. 长江流域资源与环境, 2004,13(3):266-271.
[20]Urban U R, Dinke C, Wehrli B. Solute transfer across the sediment surface of a eutrophic lake: I. Porewater profiles from dialysis samplers [J]. Aquatic Science, 1997,59:1-25.
[21]Xie L Q, Xie P, Tang H J. Enhancement of dissolved phosphorus release from sediment to lake water by microcystis blooms--an enclosure expriment in a hyper-eutrophic, subtropical Chinese lake [J]. Environ Pollut, 2003, 122:391-399.
[22]Denis L, Grenz C, Alliot E, Rodier M. Temporal variability in dissolved inorganic nitrogen flux at the sediment-water interface and related annual budget on a continental shelf (NW Mediterranean) [J]. Oceanoogical Acta, 2001,24:85-97.
[23]Lijklemal L. Interaction of orthophosphate with iron and aluminum hydroxides [J]. Environmental Science and Technology,1980,14:537-541.
[24]张西科,张福锁,毛达如.水稻根麦铁氧化物胶膜对水稻吸收磷的影响 [J]. 植物营养与肥料学报, 1997,3(4):295-299.
[25]刘文菊,朱永官.湿地植物根表的铁锰氧化物膜 [J]. 生态学报,2005,25(2):358-363.
[26]马 凯,蔡庆华,谢志才,等.沉水植物分布格局对湖泊水环境N、P的因子影响 [J]. 水生生物学报, 2003,27(3):231-237.
[27]包先明,陈开宁,范成新.沉水植物生长对沉积物间隙水中的氮磷分布及界面释放的影响 [J]. 湖泊科学, 2006,18(5):515-522.
[28]王圣瑞,金相灿,赵海超,等.沉水植物黑藻对上覆水中各形态磷浓度的影响 [J]. 地球化学, 2006,35(2):179-186.
