饲料糖水平对方格星虫稚虫生长、体组成和消化酶活性的影响
2013-08-22许明珠童万平董兰芳程光平黄国强
许明珠 张 琴 童万平 董兰芳 童 潼 程光平 黄国强 蒋 艳
(1.广西大学动物科学技术学院,南宁530005;2.广西壮族自治区海洋研究所,广西海洋生物技术重点实验室,北海536000)
方格星虫(Sipunculus nudus)属星虫动物门星虫科方格星虫属动物,亦称光裸方格星虫或光裸星虫,俗称“沙虫”,在我国山东、福建、广东、广西和台湾沿海地区均有分布[1],是我国名贵海产珍品,也是发展海水养殖的主要品种之一。方格星虫以肉脆味美,富含多种蛋白质、氨基酸、微量元素,并兼具一些食疗作用而闻名于世,素有海里的“冬虫夏草”美誉,在我国南部沿海已广泛养殖[2]。目前,有关方格星虫营养生理的研究已有相关报道[3-5]。
传统上,鱼粉是水产动物饲料中的主要蛋白质源,但近年来,随着水产养殖业的迅猛发展,鱼粉的需求量也越来越大,导致了鱼粉价格不断上涨,饲料成本随之升高,严重制约了水产养殖业的迅速发展。张琴等[4]在对方格星虫稚虫的研究中指出,方格星虫稚虫需要至少46%的饲料蛋白质,而大部分蛋白质需要鱼粉来供给。据“蛋白质节约效应”,在饲料中添加一定量的脂肪或糖类,部分替代用于能源的蛋白质,从而提高水生动物对蛋白质的利用率,可以起到节约蛋白质的作用[6]。糖类作为生物重要的能源物质,经消化吸收后能直接为机体的生理活动提供能量,是饲料中廉价的供能物质。在一些学者对鱼类的研究中发现,在饲料中添加可满足鱼类代谢需求且不引起不耐受的适量糖可以促进鱼类的生长[7-10]。然而,有关饲料中糖对方格星虫影响的研究尚未见相关报道。在鱼粉价格日益飙升且其资源受季节性制约严重的前提下,基于糖类作为饲料中最廉价的能量来源及其广阔的市场开发前景,本研究开展了饲料糖水平对方格星虫稚虫生长、体组成和消化酶活性影响的研究,以期为星虫类营养生理学及规模化养殖方格星虫人工配合饲料的研制提供科学依据。
1 材料与方法
1.1 试验饲料
以鱼粉和酪蛋白为蛋白质源,鱼油为脂肪源,糊化玉米淀粉为糖源,微晶纤维素为调节组分,并补充维生素预混料和矿物质预混料,配制出糊化玉米淀粉含量分别为0、6%、12%、18%、24%和30%的6种等氮等脂试验饲料(表1)。经3,5-二硝基水杨酸法测定,6种试验饲料中可消化糖水平的实测值分别为1.89%、7.35%、12.81%、18.27%、23.73%和29.19%(本试验中不可消化糖可忽略不计,饲料可消化糖水平等同于饲料糖水平)。在饲料配制前,主原料先粉碎过400目筛网,然后按配方各自称重混合均匀,再与鱼油和水充分混匀,微黏合饲料加工参照Blair等[11]介绍的方法,将之制成过150目筛的微颗粒饲料,装袋标号备用。

表1 试验饲料组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of experimental diets(DM basis) %
1.2 试验设计与饲养管理
饲养试验于2011年9月18日开始在广西海洋研究所海水增养殖试验基地进行。本试验所采用方格星虫稚虫为广西海洋研究所自主研发培育的同一批受精卵孵化的人工苗种。选用健康无病、体重相近[平均体重为(15.49±0.03)mg]的方格星虫稚虫7 200条,随机分成6组(每组3个重复,每个重复400条),分别饲喂等氮等脂的不同糖水平的人工微颗粒饲料。试验用方格星虫稚虫以重复为单位放养于水族箱(65cm×55cm×45cm)内,箱底铺一薄层(3~4cm)的细沙以供试验稚星虫栖息。试验期为8周,每天换水2次(08:30和17:30),每次换水约1/3;每次换水后投喂,采用稍过量投喂,保持底层沙表面有少量剩饵;每周彻底清理试验水槽1次。养殖所用海水经室外蓄水池沉淀,2级砂滤池过滤,进水槽前再经滤袋过滤。试验期间连续充气,采用自然光照周期,水温维持在26~30℃,盐度维持在18~22,溶解氧大于5.0mg/L。
1.3 样品采集与指标测定
1.3.1 生长指标的测定
饲养试验结束后,将试验稚星虫以重复为单位转移至底部无沙的水槽内2~3d进行吐沙,待其消化道内完全排净沙子之后,对每个水槽内的试验稚星虫进行计数并称重,用以计算各组试验稚星虫的成活率、增重率及特定生长率。
成活率(survival rate,SR,%)=100×Nt/N0;
增重率(weight gain rate,WGR,%)=100×(Wt-W0)/W0;
特定生长率(specific gain rate,SGR,%/d)=100×(lnWt-lnW0)/t。
式中:N0为初始稚星虫条数;Nt为终末稚星虫条数;W0为初始稚星虫体重;Wt为终末稚星虫体重;t为试验天数。
1.3.2 体组成的测定
每重复取100条方格星虫稚虫,鲜样用冷冻干燥机冷冻干燥之后,放入45℃的烘箱内烘8h,在干燥器中冷却至室温,之后用手术剪将其剪碎,标号装袋备用。采用 AOAC(1995)[12]的方法分析测定试验稚星虫的体组成,具体为:采用105℃恒重法测定水分含量;采用凯氏定氮法(Kjeltec 8400,Sweden)测定粗蛋白质含量;采用索氏抽提法(Soxtec 2050,Switzerland)测定粗脂肪含量;采用550℃高温灼烧法测定灰分含量。
1.3.3 消化酶活性的测定
每重复取30条方格星虫稚虫,按照质量体积比1∶1的比例加入pH=7的磷酸盐缓冲液,在冰浴上用玻璃匀浆器将其捣碎,4℃下3 000r/min离心10min,取上清液作为待测酶液。蛋白酶活性采用福林-酚法测定,脂肪酶和淀粉酶活性采用南京建成生物工程研究所生产的试剂盒测定。以牛血清蛋白作为标准,采用考马斯亮蓝法测定酶液中蛋白质含量。
1.4 数据统计与分析
试验结果采用平均值±标准误表示,采用SPSS 13.0软件进行统计分析,若差异达到显著水平(P<0.05),则进行Tukey氏多重比较。
2 结 果
2.1 饲料糖水平对方格星虫稚虫生长的影响
由表2可知,各组的成活率均在90.92%~92.92%之间,组间差异不显著(P>0.05)。当饲料糖水平为1.89%~18.27%时,增重率和特定生长率随着饲料糖水平的增加而显著上升(P<0.05),当饲料糖水平继续增加至23.73%时,增重率和特定生长率则显著下降(P<0.05),在饲料糖水平为18.27%时,二者均达到最大值。通过回归分析(图1),增重率与饲料糖水平之间存在二次曲线关系,增重率(y)与饲料糖水平(x)的回归方程为:y=-0.329 1x2+12.188x+154.24(R2=0.911 6)。根据回归方程可知,当增重率达到最大值时,饲料糖水平为18.52%。
2.2 饲料糖水平对方格星虫稚虫体组成的影响
由表3可知,饲料糖水平对虫体粗蛋白质和粗脂肪含量有显著影响(P<0.05),而对虫体水分和灰分含量无显著影响(P>0.05)。虫体粗蛋白质含量随着饲料糖水平的增加呈先升后降的趋势,饲料糖水平为23.73%时虫体粗蛋白质含量最高,达69.26%,并显著高于3个低糖水平(1.89%、7.35%和12.81%)组(P<0.05)。虫体粗脂肪含量随着饲料糖水平的增加呈持续升高的趋势,最高糖水平(29.19%)组虫体粗脂肪含量最高,为2.79%,显著高于3个低糖水平(1.89%、7.35%和12.81%)组(P<0.05)。
2.3 饲料糖水平对方格星虫稚虫消化酶活性的影响
由表4可知,随着饲料糖水平的增加,虫体蛋白酶和淀粉酶活性均呈先升后降的趋势。饲料糖水平为23.73%的组虫体蛋白酶活性最高,其次是饲料糖水平为29.19%的组,且这2组的虫体蛋白酶活性均显著高于其他糖水平组(P<0.05)。饲料糖水平为18.27%的组的虫体淀粉酶活性最高,显著高于其他糖水平组(P<0.05)。饲料糖水平对方格星虫稚虫虫体脂肪酶活性没有显著影响(P>0.05)。
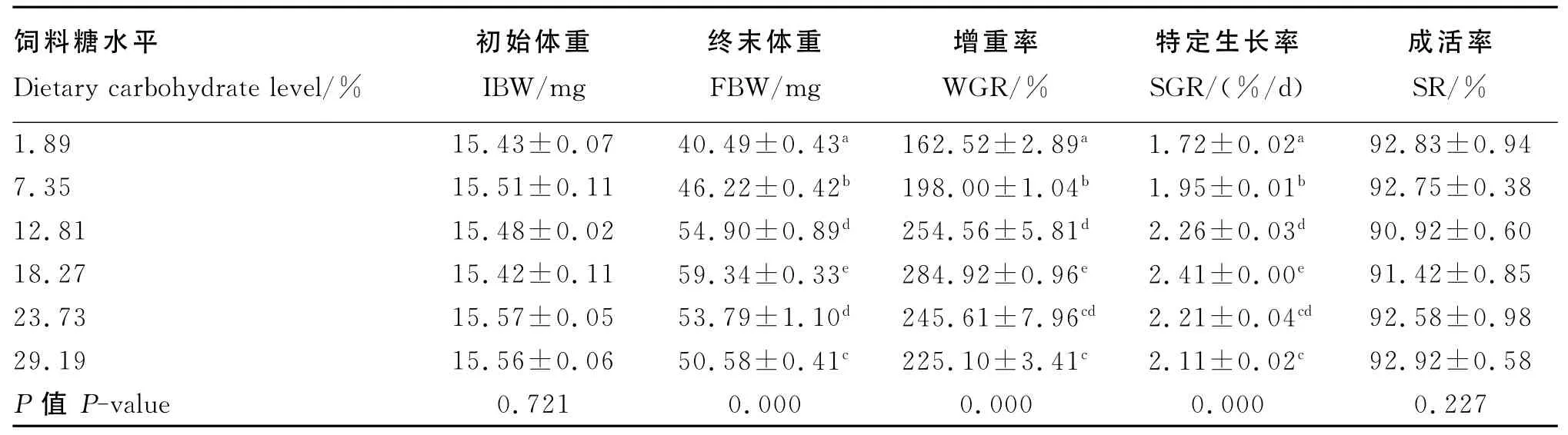
表2 饲料糖水平对方格星虫稚虫生长的影响Table 2 Effects of dietary carbohydrate level on growth of juvenile peanut worm
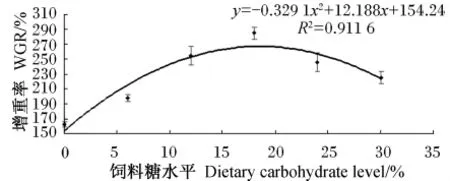
图1 方格星虫稚虫的增重率与饲料糖水平的回归分析Fig.1 Regression analysis between WGR of juvenile peanut worm and dietary carbohydrate level
3 讨 论
3.1 饲料糖水平对方格星虫稚虫生长的影响
李葳等[13]报道,随着饲料糖水平的增加,点篮子鱼(Siganus guttatus)的摄食率和特定生长率呈先增加后降低的趋势,点篮子鱼的饲料最适糖水平为 20.83%。 蔡 春 芳 等[14]在 对 异 4 育 银 鲫(Carassius auratus gibelio)的研究中发现,随着饲料糖水平的增加,异育银鲫的特定生长率呈先增加后降低的趋势。这一现象在其他学者对南方鲇(Silurus meridionalis)[15]、宝 石 鲈 (Scorturm barco)[16]和厚唇若棘(Hephaestus fuliginosus)[17]上的研究中也同样出现。本研究中,饲料糖水平较低时,增重率随着饲料糖水平的增加而显著升高,当饲料糖水平达到18.27%时,增重率和特定生长率都达到最大值,之后饲料糖水平再增加增重率和特定生长率反而缓慢地下降,这一结果与上述学者在其他水产动物上的研究得到的结论是一致的。方格星虫稚虫增重率和特定生长率能随饲料糖水平在一定范围内增加而上升,证明其对糖的消化吸收能力可随着饲料糖水平的增加而增加,但这种能力有限。有关饲料糖水平对鱼类生长影响的研究中,既有提高饲料糖水平没有提高鱼类生长[18-20]的报道,又有一些草食性鱼类,如鲤鱼(Cyprinus carpio)[21]能耐受35%~40%的淀粉、南亚野鲮(Labeo rohita)[22]能耐受35%~40%的糊精的报道,还有大菱鲆(Scophthalmus maximus L.)幼鱼[23]和65g大菱鲆[24]饲料糖的适宜水平分别为4.0%和16.2%的报道。不同结论的得出可能与糖的种类、添加量及水产动物的种类和规格等有关[6]。鱼类对饲料主要能源物质中蛋白质的利用率和需求最高,其次是脂肪,最低的是糖[25]。然而,马爱军等[23]研究大菱鲆时发现,饲料中主要能源物质对大菱鲆体增重和饲料转化率的影响顺序为蛋白质>糖>脂肪。据张琴等[3-4]关于饲料蛋白质和脂肪水平对方格星虫稚虫生长性能的影响,并结合本试验的研究结果来看,对方格星虫稚虫增重率影响最大的是饲料蛋白质水平,其次是饲料糖水平,影响最小的是饲料脂肪水平。因此,如果投喂糖水平过低的饲料,饲料中其他营养成分则不能得到充分利用,这可能也是本研究中过低的饲料糖水平限制了方格星虫稚虫生长的原因。本研究中,当饲料糖水平过高时,方格星虫稚虫的生长表现出一定程度的下降,可能的原因:一是饲料糖水平过高,虫体虽然吸收了,但是消化吸收进入血液循环之后却不能进一步有效利用。二是饲料中降低的糖水平由纤维素填充,而纤维素对水产动物饲料糖的利用也有影响。Shiau[25]发现纤维素使奥尼罗非鱼(Oreochromis niloticus×O.aureus)增重率显著降低;Morita等[26]发现羧甲基纤维素提高了真鲷(Chrysophrys major)对糊精的利用。三是饲料中糖超适宜水平的添加可能导致方格星虫稚虫的摄食率下降,进一步导致其生长速率下降。在对鱼类的研究中发现,饲料糖水平超过一定范围时,可能会增加鱼体糖原含量、血糖水平及葡萄糖的周转率,而这些因素特别是血糖水平持续升高和高的葡萄糖周转率可能抑制摄食[27]。付世建等[15]对南方鲇的有关研究表明,饲料糖的添加可能导致摄食率的下降,而饲料糖吸收速率越快,对摄食的抑制作用越大。另外,Shiau[25]和 Dixon等[28]也发现饲料糖水平超过一定范围,鱼体会出现一些不良症状,如血糖过高、肝糖原积累、免疫力变化、生长停滞等。本研究中,饲料糖水平对方格星虫稚虫生长影响的具体机制还有待进一步的研究。

表3 饲料糖水平对方格星虫稚虫体组成的影响Table 3 Effects of dietary carbohydrate level on body composition of juvenile peanut worm %
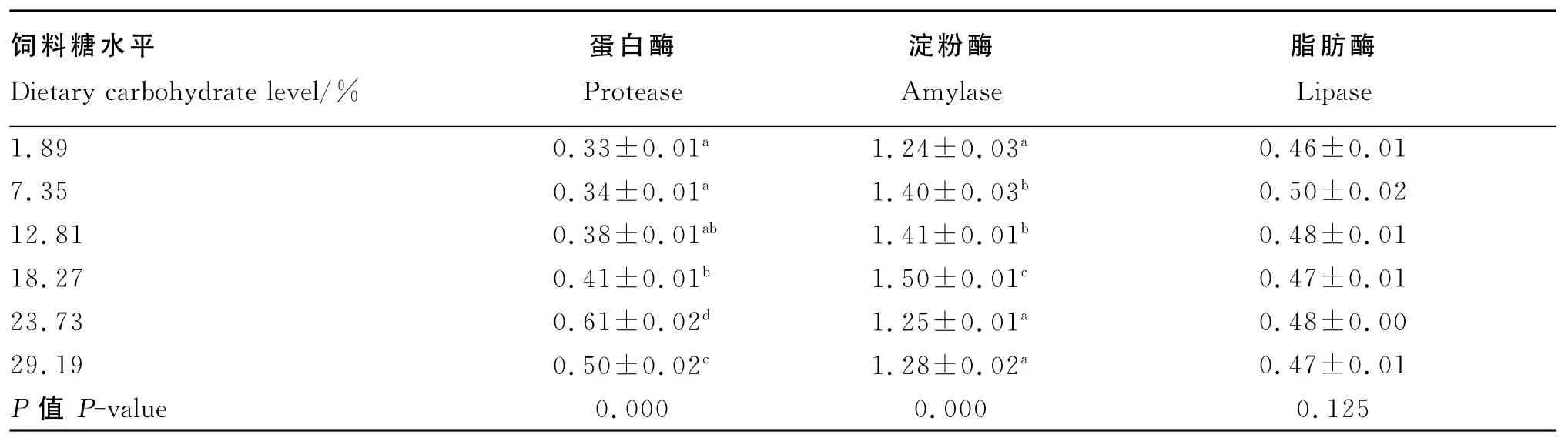
表4 饲料糖水平对方格星虫稚虫消化酶活性的影响Table 4 Effects of dietary carbohydrate level on digestive enzyme activities of juvenile peanut worm U/mg prot
3.2 饲料糖水平对方格星虫稚虫体组成的影响
有研究表明,饲料糖水平对点篮子鱼全鱼水分含量的影响不显著[13],这与本研究中饲料糖水平对方格星虫稚虫虫体水分含量无显著影响是一致的。蔡春芳等[14]研究发现,超过适宜糖水平(36%)的高糖水平(50%)饲料对异育银鲫鱼体灰分含量没有显著影响,本研究也得到了相似的结论。造成方格星虫稚虫虫体灰分含量与饲料糖水平的变化无显著关联的原因可能在于本试验中各试验饲料中鱼粉用量相同,鱼粉中的矿物质在体内沉积能导致灰分的增加[4]。
蔡春芳等[14]研究发现,饲料糖水平的增加使异育银鲫肌肉中粗蛋白质含量有增加的趋势。Hemre等[8]研究发现,高糖饲料喂养的鲫鱼(Carassius auratus)全鱼和肝胰脏中粗蛋白质的含量比低糖饲料喂养的鲫鱼要高。Enes等[20]对金头鲷(Sparus aurata)和 Zhang 等[29]对 方 斑 东 风 螺(Babylonia areolate)的研究中也发现,饲料糖水平可以提高体蛋白质含量。本研究中,摄食含高糖(23.73%)饲料8周后,方格星虫稚虫虫体粗蛋白质含量显著增加,这与上述学者的研究结果是相似的。鱼类饲料中糖具有节约蛋白质的作用,当蛋白质与糖一起被摄入鱼体时,在鱼体内储留的氮比单独摄入蛋白质时要多[30]。因此,方格星虫稚虫虫体粗蛋白质含量显著增加的原因可能是因为饲料中适宜水平的糖不但能代替部分作为能量消耗的蛋白质,还可以为非必需氨基酸提供碳源,并抑制氨基酸分解酶的活性,从而提高蛋白质的利用率[6]。
吴凡等[31]在对奥尼罗非鱼幼鱼的研究中发现,饲料糖水平增加能显著提高其全鱼脂肪含量。郭冉等[32]对南美白对虾(Penaeus vannamei)投喂不同糖水平的饲料后发现高糖组的体脂肪含量相对较高。本研究中,方格星虫稚虫虫体粗脂肪含量也随着饲料糖水平的增加而升高,高糖水平(18.27%、23.73%和29.19%)组的虫体 粗脂肪含量显著高于低糖水平(1.89%、7.35% 和12.81%)组,这与其他学者在虹鳟(Oncorhynchus mykiss)[33]、鳙 鱼 (Aristichthys nobilis)[34]、金 头鲷[20]、凡纳滨对虾(Litopenaeus vianname)[35]以及点篮子鱼[13]上的研究结果一致。能源物质能在体内相互转化,多余的糖在体内代谢的途径可以是转氨基成为蛋白质,但一般情况下是转化为脂肪储存起来,所以饲料中大量糖被虫体吸收后,除去作为能源物质消耗掉一部分,剩下的很有可能在体内通过代谢途径转化为脂肪储存起来[13]。
另外,由于糖、蛋白质、脂肪、灰分、水分含量相互之间的牵制,如饲料糖水平对灰分的影响很有可能是影响水分及有机质含量的反映,以及能源物质在体内的相互转化[8],因此,尚不能从本研究中得出关于饲料糖在方格星虫稚虫体内代谢去向的明确结论。
3.3 饲料糖水平对方格星虫稚虫消化酶活性的影响
消化酶是指由消化系统和消化腺分泌的,起营养消化作用的酶类。饲料成分对鱼类消化酶活性影响的报道有很多,研究结果虽不尽相同,但一般的变化规律为饲料中某种成分增加,消化该成分的消化酶活性增加,而其他种类的消化酶活性也会发生相应的变化[36],即消化酶活性的大小变化与食物组成的变化有一定关系[37]。黎军胜等[38]研究饲料成分与环境温度对奥尼罗非鱼消化酶活性的影响时发现,饲料成分对鱼类消化酶活性的影响具有复杂性和效应的多样性。
方格星虫稚虫在摄食含高糖(23.73%和29.19%)的饲料8周后,其虫体蛋白酶活性显著提高,这表明稚星虫能逐渐适应饲料糖,在一些鱼类上的研究也得到了相似的结论[39-41]。作为酶作用的底物,饲料中各养分的含量往往会引起消化酶的“适应性分泌”,Flores等[42]报道了提高猪饲粮中糖类的含量能提高其消化道中淀粉酶的活性。因此,在饲料中添加适宜糖水平时,淀粉酶活性的增加可能原因是方格星虫稚虫需要更多的淀粉酶来消化饲料中的糖。消化酶之间具有一定的协同作用[43],蛋白酶活性的增加可能原因是饲料中糖类已使方格星虫稚虫所需能量得到满足,节省了作为能量消耗的一部分蛋白质,进而可能导致消化系统分泌更多的蛋白酶来分解蛋白质,促使饲料蛋白质高效率地转化为体蛋白质。影响方格星虫稚虫消化酶活性变化的因素有很多,饲料糖水平对其影响的具体机理还有待进一步研究。
4 结 论
① 饲料糖水平过低会造成方格星虫稚虫生长缓慢,糖水平过高则不仅造成其生长缓慢,还会出现体脂沉积。
② 以增重率为评价指标,方格星虫稚虫饲料中糖的适宜水平为18.57%。
[1] 李凤鲁,孔庆兰,史贵田,等.中国沿海方格星虫属(星虫动物门)的研究[J].青岛海洋大学学报,1990,20(1):93-99.
[2] 张桂和,李理,赵谋明,等.方格星虫营养成分分析及抗疲劳作用研究[J].营养学报,2008,30(3):318-320.
[3] 张琴,童万平,董兰芳,等.饲料中脂肪水平对方格星虫稚虫生长性能、体组成及消化酶活性的影响[J].渔业科学进展,2011,32(6):99-106.
[4] 张琴,童万平,董兰芳,等.饲料蛋白水平对方格星虫稚虫生长和体组成的影响[J].渔业科学进展,2012,33(1):86-92.
[5] 张琴,童潼,童万平,等.饲料蛋白水平对方格星虫稚虫日增重和消化酶活性的影响[J].渔业现代化,2012,39(2):41-46.
[6] 李孟均,陈春娜.饲料中的糖对水产动物的影响[J].北京水产,2008(2):57-59.
[7] KIM J D,KAUSHIK S J.Contribution of digestible energy from carbohydrates and estimation of protein/energy requirements for growth of rainbow trout(Oncorhynchus mykiss)[J].Aquaculture,1992,106:161-169.
[8] HEMRE G I,SANDNES K,LIE Ø,et al.Carbohydrate nutrition in Atlantic salmon,Salmo salar L.,growth and feed utilization[J].Aquaculture Research,1995,26:149-154.
[9] ERFANULLAH,JAFRI A K.Growth rate,feed conversion,and body composition of Catla catla,Labeo rohita,and Cirrhinus mrigala fry fed diets of various carbohydrate-to-lipid ratios[J].Journal of the World Aquaculture Society,1998,29:84-91.
[10] ERFANULLAH,JAFRI A K.Growth,feed conversion,body composition and nutrient retention efficiencies in fingerling catfish,Heteropneustes fossilis(Bloch),fed different sources of dietary carbohydrate[J].Aquaculture Research,1999,30:43-49.
[11] BLAIR T,CASTELL J,NEIL S,et al.Evaluation of microdiets versus live feeds on growth,survival and fatty acid composition of larval haddock (Melanogrammus aeglefinus)[J].Aquaculture,2003,225:451-461.
[12] AOAC.Official methods of analysis of official analytical chemists international[S].16th ed.Arlington,V A:Association of Official Analytical Chemists,1995.
[13] 李葳,侯俊利,章龙珍,等.饲料糖水平对点篮子鱼生长性能的影响[J].海洋渔业,2012,34(1):64-70.
[14] 蔡春芳,王道尊.异育银鲫对糖利用性的研究:饲料糖含量对生长、消化吸收的影响[J].苏州大学学报:自然科学版,1999,15(3):87-90.
[15] 付世建,谢小军.饲料碳水化合物水平对南方鲇生长的影响[J].水生生物学报,2005,29(4):393-398.
[16] 宋理平.宝石鲈营养需求的研究[D].博士学位论文.济南:山东师范大学,2009.
[17] 宋理平,韩勃,王爱英,等.碳水化合物水平对厚唇弱棘鯻生长和血液指标的影响[J].大连水产学院学报,2010,25(4):293-297.
[18] HEMRE I,SHIAU S Y,DENGD F,et al.Utilization of hydrolysed potato starch by juvenile Atlantic salmon Salmo salar L.,when using a restricted feeding regime[J].Aquaculture Research,2000,31:207-212.
[19] ENES P,PANSREAT S,KAUSHIK S,et al.Effect of normal and waxy maize starch on growth,food utilization and hepatic glucose metabolism in European sea bass(Dicentrarchus labrax)juveniles[J].Comparative Biochemistry and Physiology,2006,143A:89-96.
[20] ENES P,PANSERAT S,KAUSHIK S,et al.Growth performance and metabolic utilization of diets with native and waxy maize starch by gilthead sea bream(Sparus aurata)juveniles[J].Aquaculture,2008,274:101-108.
[21] WILSON R P.Utilization of dietary carbohydrate by fish[J].Aquaculture,1994,124:67-80.
[22] SATPATHY B B,RAY A K.Effect of dietary protein and carbohydrate levels on growth,nutrient utilization and body composition in fingerling rohu,Labeo rohita(Hamilton)[J].Journal of Applied Ichthyology,2009,25(6):728-733.
[23] 马爱军,陈四清,雷霁霖,等.饲料中主要能量物质对大菱鲆幼鱼生长的影响[J].海洋与湖沼,2001,32(5):527-533.
[24] REGOST C,ARZEL J,KAUSHIK S J.Partial or total replacement of fish meal by corn gluten meal in diet for turbot(Psetta maxima)[J].Aquaculture,1999,180:99-117.
[25] SHIAU S Y.Utilization of carbohydrates in warmwater fish-with particular reference to tilapia,Orechromis niloticus×O.aureus[J].Aquaculture,1997,151:79-96.
[26] MORITA K,FUREICHI M,YONE Y.Effect of carboxymethylcellulose supplemented to dextrin-containing diets on the growth and feed efficiency of red sea bream (Chrysophrys major)[J].Bulletin of the Japanese Society of Scientific Fisheries,1982,48:1617-1620.
[27] HEMRE G I,HANSEN T.Utilisation of different dietary starch sources and tolerance to glucose loading in Atlantic salmon (Salmo salar),during parr-smolt transformation[J].Aquaculture,1998,161:145-157.
[28] DIXON D G,HILTON J W.Influence of available dietary carbohydrate content on tolerance of waterborne copper by rainbow trout,Salmo gairdneri Richardson[J].Fish Biology,1981,19(5):509-518.
[29] ZHANG L L,ZHOU Q C,CHEN Y Q.Effect of diet-ary carbohydrate level on growth performance of juvenile spotted babylon(Babylonia areolata Link 1807)[J].Aquaculture,2009,295:238-242.
[30] COWEY C B,ADRON J W,BROWN D A,et al.Studies on the nutrition of marine flatfish.The metabolism of glucose by plaice(Pleuronectes platessa)and the effect of dietary energy source on protein utilization in plaice[J].British Journal of Nutrition,1975,33:219-231.
[31] 吴凡,文华,蒋明,等.饲料碳水化合物水平对奥尼罗非鱼幼鱼生长、体成分和血清生化指标的影响[J].华南农业大学学报,2011,32(4):91-95.
[32] 郭冉,刘永坚,田丽霞,等.不同糖源对南美白对虾Penaeus vannamei生长、成活率和虾体组成的影响[J].中山大学学报:自然科学版,2005,44(3):90-92.
[33] KAUSHIK S J,MEDALE F,FAUCONNEAU B,et al.Effect of digestible carbohydrates on protein/energy utilization and on glucose metabolism in rainbow trout(Salmo gairdneri R.)[J].Aquaculture,1989,79:63-74.
[34] SANTIAGO C B,REYES O S.Optimum dietary protein level for growth of bighead carp(Aristichthys nobilis)fry in a static water system[J].Aquaculture,1991,93:155-165.
[35] 胡毅,谭北平,麦康森,等.不同碳水化合物水平饲料对凡纳滨对虾生长及部分生理生化指标的影响[J].水生生物学报,2009,33(2):289-295.
[36] GANGADHARA B N,EESHA M C,VARGHESE T J,et al.Effect of varying protein and lipid levels on the growth of rohu,Labeo rohita[J].Asian Fisheries Society,1997,10(2):139-147.
[37] 李军,吴天星.鱼类消化酶的影响因素[J].水利渔业,2006,26(6):30-32.
[38] 黎军胜,李建林,吴婷婷.饲料成分与环境温度对奥尼罗非鱼消化酶活性的影响[J].中国水产科学,2004,11(6):585-588.
[39] ROSAS C,CUZON G,GAXIOLA G,et al.Influence of dietary carbohydrate on the metabolism of juvenile Litopenaeus stylirostris[J].Experimental Marine Biology and Ecology,2000,249:181-198.
[40] MOHAPATRA M,SAHU N P,CHAUDHARI A.Utilization of gelatinized carbohydrate in diets of Labeo rohitafry[J].Aquaculture Nutrition,2003,9:189-196.
[41] 高梅,罗毅平,曹振东.饲料碳水化合物水平对南方鲇幼鱼消化酶活性的影响[J].西南师范大学学报,2006,31(2):119-123.
[42] FLORES C A,BRANNON P M,BUSTAMANTE S A,et al.Effect of diet on intestinal and pancreatic enzyme activities in the pig[J].Pediatric Gastroenterology Nutrition,1988,7:914-921.
[43] 张丽丽,周歧存,程怡秋,等.不同糖源对方斑东风螺生长、饲料利用和消化酶活性的影响[J].广东海洋大学学报,2009,29(4):14-18.
