白刺属植物的繁育系统与开花结实特征1)
2013-08-08李清河
李清河 徐 军
(国家林业局林木培育重点实验室(中国林业科学研究院林业研究所),北京,100091) (中国林业科学研究院沙漠林业实验中心)
高婷婷 颜 秀 辛智鸣
(河北省唐秦水文水资源勘测局) (中国林业科学研究院林业研究所) (中国林业科学研究院沙漠林业实验中心)
对植物繁育系统的了解是认识植物生活史的前提,也是其它相关研究所需的基本背景知识[1-2]。国内外部分学者对植物的花粉及其与气候关系,植物胚胎、种子萌发及人工授粉等方面进行过一些研究工作[3-7]。植物繁育系统,最早可追溯到由 Darwin利用人工杂交方法揭示了自交和异交的效果及不同花型的适应和影响[8]。自交和异交是植物繁育系统研究的一对中心问题[9-10]。也有学者指出,植物进化过程中的自交和近交衰退问题。到目前为止,仍有两种观点争论不休,但是需要看到进化是折中的。另外,开花结实是植物进化中的关键阶段。不同植物的开花与结实量存在较大差异,植物自身生物学特性、外部环境条件等导致植物的果实种子形态、数量特征的不同。
白刺属植物作为第三纪残遗植物能够延续至今,其繁育生物学特性的研究对掌握其演化维持机制具有重要的意义。然而,针对分布于我国的白刺属植物种,系统地从繁育系统上探索白刺属不同植物种的繁殖生物学特性问题,国内外却少有系统的报道。基于以往研究,我们假设:①白刺属植物的繁育系统属于混合交配系统,表现为兼性异交;②白刺属的不同植物种之间的开花结实数量与形态特征存在差异。为此,我们通过调查白刺属植物的生殖器官形态、交配方式、现时生殖值[11]等指标,统计分析白刺属植物生殖生物学数据,检验上述假设。这可为白刺属植物系统进化研究提供生殖生态学的科学依据。同时,也对进一步分析研究白刺属植物的结实规律、揭示其种群生态适应性机理和促进种群恢复有重要意义。
1 试验地概况
研究区位于内蒙古西部乌兰布和沙漠东北部地区。该地区气候属于亚洲中部温带荒漠气候,寒冷、干燥、多风,兼有大陆性和季风降雨气候特点。年平均降水量149.0 mm,主要集中在7—9月份,年平均蒸发量2 351.9 mm。唐古特白刺和大白刺主要生长在固定和半固定沙地,西伯利亚白刺主要生长在沿黄的盐碱低地,泡泡刺主要生长在狼山山前的戈壁滩地,其中唐古特白刺的分布范围最为广泛。乌兰布和沙漠地处草原化荒漠地带,植被以旱生灌木为主,白刺群落是该沙区的主要优势群落之一,其它优势种还有油蒿(Artemisia ordosica Krasch.)、白沙蒿(Artemisia sphaerocephala Krasch.)等。
2 材料和方法
2.1 试验材料
白刺属(Nitraria L.)为蒺藜科的一个古老小属,主要植物种有唐古特白刺(Nitraria tangutorum Bobr.)、西伯利亚白刺(Nitraria sibirica Pall.)、大白刺(Nitraria roborowskii Kom.)、泡泡刺 (Nitraria sphaerocarpa Maxim.),均属旱生或超旱生灌木或小灌木,在乌兰布和沙漠地区均有分布。如果春季有适宜的降水,白刺属植物每年4月末5月初返青,长出嫩叶;之后从5月至8月,该属种群开花、结果。一般唐古特白刺的盛花期在5月底,大白刺在5月中旬,泡泡刺在5月上中旬;而西伯利亚白刺最晚,约比唐古特白刺晚7~10 d。对于果期,唐古特白刺、西伯利亚白刺和大白刺一般在7月中旬,而泡泡刺的果期要提前半个月左右,但花期、果期在年际间变化较大。白刺属植物的花属于聚伞状花序顶生。唐古特白刺、西伯利亚白刺和大白刺的果实都是球形的浆果状核果,且酸甜可口;而泡泡刺的果实为密被绒毛的干膜质果。果核种子为褐色,顶端渐尖核卵形,上有分布不均、深浅不一、大小不等的孔穴,种皮角质厚[12]。
2.2 研究方法
2.2.1 调查方法
于2009年5月各种白刺的花期,对试验样地的各种白刺样株记录开花过程,如花药开裂时间与柱头可授期之间的时间间隔、柱头与花药的空间位置等。每种白刺在不同样地上随机选取5株样株,每株取5个花序,测量其开放花朵数;每物种随机取开放状态的花朵至少15朵,测量花朵直径、雄蕊群平均长度、雌蕊长度等花部形态特征,用于杂交指数(IOC)的估算。
在各种白刺的盛花期分别随机选取即将开放的花蕾(花药未开裂)各10朵,置于冰盒(0~4℃)保鲜,带回实验室备用。统计每朵花的雄蕊数目。取下花药,用1.0 mL·L-1HCl软化花药壁。然后将花粉全部移入1个具刻度的离心管,用蒸馏水定容至1 mL。在震荡器下震荡60 s后,用移液枪吸取1 μL的花粉液于载玻片上,在显微镜下观察并统计单花花粉数,每个花药重复10次。用于计算花粉-胚珠比(P/O)。
由于白刺植物单株的花、果实的数目庞大,特别是唐古特白刺、西伯利亚白刺和大白刺,记录清楚每株的生殖器官数目有一定难度。为此,我们通过选取不同大小的白刺植株作为样株,在每个样株上选取不同大小且具多级分枝的分株(西伯利亚白刺和泡泡刺是选取一个枝条作为标准枝)作为典型代表,在5月份盛花期,选定3株试验分株(标准枝),每株上选3个有代表性的生殖枝,标记其上的新生枝(花枝,它们均为顶生聚伞花序),随后在白刺的一个单位生长季的花期和果期记录每个生殖枝的单枝花数(花蕾和花)、果实数(成熟的和未成熟的)以及凋落的花数、果实数,统计即时生殖器官数和结实率。
果实、种子外部形态观察:用目测法观察果实、种子的形状、颜色,并进行描述。用精度为0.01 mm游标卡尺测量果实、种子的纵径、横径,之后计算果(种)形指数(纵径/横径)。
2.2.2 数据处理
通过对已开放的样株的花朵等花部形态特征以及果实种子形态特征的测量,用方差分析比较不同白刺种的花部形态、果实种子形态特征的差异性。另据文献记载[13-14],白刺雌蕊由3心皮合生成3室子房,每室含1枚胚珠或无,仅2枚胚珠发育,最后其形成1枚种子。据此计算花粉-胚珠比(P/O),依据Cruden的标准估测其繁育系统类型[15];同时,按照Dafni[16]的标准进行杂交指数(IOC)测算及繁育系统的评判。具体方法是:①花朵或花序直径小于1 mm记为0;1~2 mm记为1;2~6 mm记为2;大于6 mm记为3。②花药开裂时间与柱头可授期之间的时间间隔,同时或雌蕊先熟记为0;雄蕊先熟记为1。③柱头与花药的空间位置,同一高度记为0;空间分离记为1。以上3者之和为IOC值。④IOC=0时,繁育系统为闭花受精;IOC=1时,繁育系统为专性自交;IOC=2时,繁育系统为兼性自交;IOC=3时,繁育系统为自交亲和,有时需要传粉者;IOC=4时,繁育系统为部分自交亲和,异交,需要传粉者。
针对白刺属植物的开花结实的数量统计,计算理论现时生殖值(PRV_T)和实际现时生殖值(PRV_A)及结实率(Fr)等开花结实数量特征。理论现时生殖值等于花期调查时的花数、蕾数与未成熟果实数之和;实际现时生殖值等于果期调查时的成熟果实数;结实率等于实际现时生殖值占理论现时生殖值的百分比。统计分析不同白刺种之间与不同现时生殖值之间的差异性。
3 结果与分析
3.1 花部特征
不同白刺属植物相应花部特征均有显著差异,见表1。每个花序上西伯利亚白刺的花数最多,其次是唐古特白刺,大白刺和唐古特白刺相差不大,数目最小的是泡泡刺;花冠直径最大的是西伯利亚白刺,其次为泡泡刺、唐古特白刺和大白刺;在柱头高度指标上,泡泡刺的最高,其次是大白刺和唐古特白刺,最小的是西伯利亚白刺;大白刺种群中有的个体没有花丝,雄蕊是直接在花基部的,这可能是一定的环境条件造成的,雄蕊长度最长的是泡泡刺,其次是西伯利亚白刺和唐古特白刺。可见,白刺属植物的花部各特征相互独立,不成比例关系。

表1 白刺属植物花部形态特征
3.2 单花花粉量、胚珠数和花粉/胚珠比率的测定
4种白刺属植物的P/O比在400~2 000,测定结果如表2。其中唐古特白刺和泡泡刺雌雄蕊等长为同型花,P/O较小(小于500),依据Cruden的标准,其繁育系统应为兼性异交;西伯利亚白刺和大白刺雌雄蕊不等长,为异型花,它们的P/O比相对较大,繁育系统也应为兼性异交。总体看,白刺属植物的繁育系统属于兼性异交类型,异交,部分自交亲和,需传粉者。

表2 白刺属植物的花粉-胚珠比
3.3 杂交指数
依据表1所测4种白刺的花部特征,唐古特白刺与大白刺的花朵直径小于6 mm,而西伯利亚白刺和泡泡刺大于6 mm;白刺属植物虽属两性花,但开花时柱头与花药分离,直到花药散粉时柱头位置仍比花药低,因此可以认为其雌、雄器官在空间上是分离的。按照Dafni的方法对白刺属植物进行杂交指数测量,结果见表3。由表3可见,4种白刺的杂交指数(IOC)为3或4。4种白刺属植物繁育系统均存在自交亲和,需要传粉者。
3.4 开花结实的数量特征
4种白刺植物的理论现时生殖值(PRV_T)与实际现时生殖值(PRV_A)如表4所示。

表3 白刺杂交指数观测结果
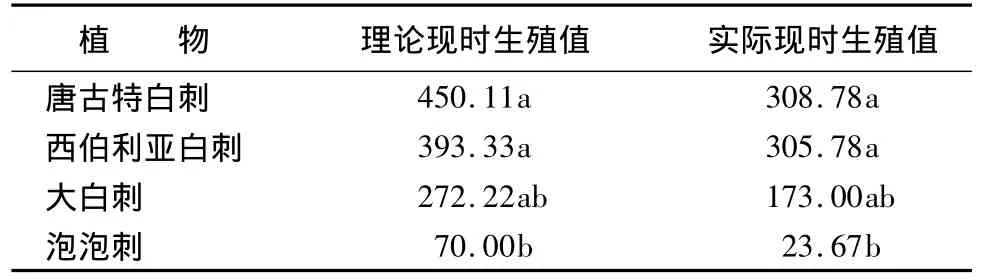
表4 4种白刺属植物单位生长季现时生殖值
不同白刺属植物种的理论现时生殖值和实际现时生殖值都有较大差距,存在显著差异,其中以泡泡刺为最小,这和其生长在戈壁生境条件下,本身植株矮小有关,其自身的生殖器官数目相对较少。同时,除泡泡刺外,其它3种白刺植物种的理论现时生殖值和实际现时生殖值间的差异均不显著。通过结实率测算,在4种白刺植物中,西伯利亚白刺的结实率(Fr)最高,达到77.78%;泡泡刺的结实率最低,只有约33.85%。
3.5 果实、种子形态特征
白刺属植物的种子形态特征详见表5。4种白刺对应果实、种子形态特征方差分析结果表明,4种白刺属植物果实、种子纵径、横径和果、种形指数差异均达到显著水平,但均未达到极显著差异。
果实与种子大小反映其营养物质积累,与子代的生长适应力相关。由表5可见,大白刺在果实形态上表现出优势,其果实的纵径与横径均大于其它白刺植物。在种子纵横径上,大白刺也较大;泡泡刺的种子纵径较大,但横径较小,从种形指数上可以看出其种子呈现长梭型。
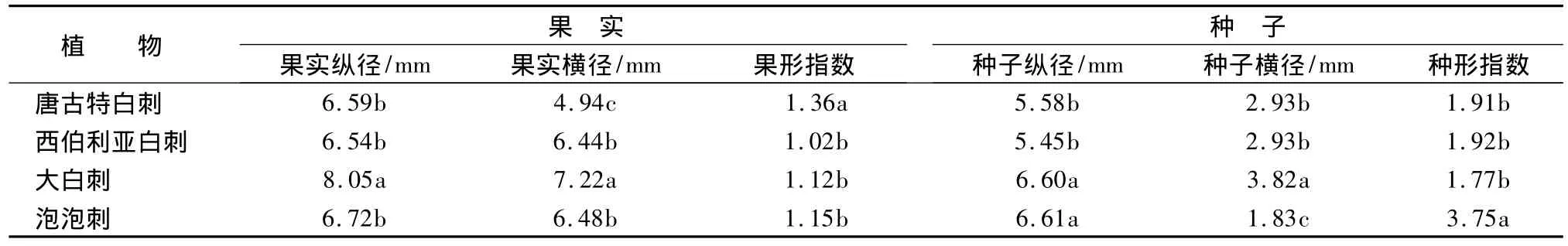
表5 4种白刺属植物果实、种子对应形态特征
4 结论与讨论
4.1 白刺属植物繁育系统
植物的繁育系统是与植物的繁殖紧密联系在一起的,是植物繁殖的核心内容之一[17],是种群有性生殖的纽带,在决定植物的进化路线和表征变异上起着重要作用[18]。4种白刺属植物的P/O和IOC值研究结果表明:白刺属植物的繁育类型为自交与异交综合特征并存的混合交配系统,这与本文的第1个假设一致。而且也与我们在野外进行套袋实验检测其繁育系统的结果一致。一般来说,自然界绝对自交或异交的植物类群很少,大多是两者兼而有之的混合交配模式。植物的繁育系统与环境因子存在着密切的联系,即与生态具有关联性[19]。沙生植物由于面临恶劣的环境条件,其繁育系统一般具有多样性,且以混合交配方式为多[20-21]。白刺等沙生植物在繁育系统上所表现出的特点,可以有效保障它们在荒漠环境等不利影响下的成功繁殖,是对荒漠环境的生态适应性表现。权衡近交和远交利弊的途径是混合的交配系统,许多学者的研究表明,混合交配系统是对植物本身和环境条件适应的一种折中的机制,可维持植物进化稳定[22-23]。它不仅能帮助植物占据新的生境、克服传粉媒介的短缺、延续种群的优良性状和提高种群的局部适应性等,还可以保证种群内的基因交流,避免“近交衰退”,增强植物对环境的适应性。
4.2 白刺属植物开花结实的数量与形态特征
不同种白刺属植物之间的理论现时生殖值和实际现时生殖值等数量特征,以及花部与果实种子的形态特征都有较大差距,这与本文的第2个假设一致。唐古特白刺和西伯利亚白刺的现时生殖值较大,这与它们的花序花数较多及其对传粉者的吸引有关。同时,白刺属植物的理论现时生殖值与实际现时生殖值之间相差不大,结实率较高,总体结实率可达到50% ~70%(泡泡刺低些,其分布区属于年降水量在100 mm以下,干燥度大于5.0的荒漠戈壁区),但是我们观察到白刺属植物在天然条件下经种子萌生的实生苗较少,这是一种借由果实产生大量种子以弥补其种子在野外条件下不易萌发的生殖补偿机制,以保证种群在自然界繁衍生息[24]。
花部形态特征是传粉昆虫重要的诱物和视觉启示,它通过影响传粉者的有效传粉从而影响植物的传粉成功[25]。白刺植株柱头较低,且花柱较粗,柱头接触面积大,能增加授粉的几率,在传粉上有一定的优势。自花传粉是被子植物进化的一种普遍趋势,是植物在恶劣的环境中(传粉者缺乏或不可预测)保证繁殖成功的一种适应机制[26]。白刺属植物花部形态为同朵花雌蕊下位,柱头和雄蕊间距较小,较易进行自花授粉,这也是其结实率高的一个原因。唐古特白刺、西伯利亚白刺和大白刺的果实都是圆形的浆果状核果,且酸甜可口;而泡泡刺的果实为干膜质果;这为其种子依靠动物采食或滚落扩散传播或借助风力滚动传播提供便利。许多沙生植物,如沙棘(Hippophae rhamnoides L.)、沙米(Agriophyllum squarrosum Moq.)等都具有类似的特性,这将有助于植物种子扩散、种群扩大。不同白刺属植物的果实种子的形态特征也存在差异,这与植物不同阶段的结实与该阶段的温度、花部形态和传粉者访问频率等有关[27-28]。花部特征差异也为果实种子的发育提供不同的基础。白刺属植物生殖器官形态上的差异,间接反映其生殖力的高低。
[1] Lincoln R,Boxshall G,Clark P.A Dictionary of Ecology,Evolution and Systematics[M].2ed.London:Cambridge University Press,1998.
[2] Bachelier J B,Endress P K.Floral structure of kirkia(Kirkiaceae)and its position in Sapindales[J].Annal of Botany,2008,102(4):539-555.
[3] Herzschuh U,Kürschner H,Ma Y Z.The surface pollen and relative pollen production of the desert vegetation of the alashan plateau,western Inner Mongolia[J].Chinese Science Bulletin,2003,48(14):1488-1493.
[4] Herzschuh U,Tarasov P,Wunnemann B,et al.Holocene vegetation and climate of the alashan plateau,NW China,reconstructed from pollen data[J].Palaeogeography Palaeoclimatology Palaeoecology,2004,211(1/2):1-17.
[5] Rodriguez-Pérez J.Breeding system,flower visitors and seedling survival of two endangered species of Helianthemum(Cistaceae)[J].Annals of Botany,2005,95(7):1229-1236.
[6] Herzschuh U,Kurschner H,Battarbee R,et al.Desert plant pollen production and a 160-year record of vegetation and climate change on the alashan plateau,NW China[J].Vegetation History and Archaeobotany,2006,15(3):181-190.
[7] Ma Y Z,Liu K B,Feng Z D,et al.A survey of modern pollen and vegetation along a south-north transect in mongolia[J].Journal of Biogeography,2008,35(8):1512-1532.
[8] Michael Proctor,Peter Yeo,Andrew Lack.The natural history of pollination[M].London:Harper Collins,1996.
[9] 何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,27(2):151-163.
[10] Takebayashi N,Morrel1 pl.Is self-fertilization an evolutionary dead end?Revisiting an old hypothesis with genetic theories and a macro-evolutionary approach[J].American Journal of Botany,2001,88(7):1143-1150.
[11] 祖元刚,张文辉,闫秀峰,等.濒危植物裂叶沙参保护生物学[M].北京:科学出版社,1999.
[12] 刘媖心.中国沙漠植物志:第2卷[M].北京:科学出版社,1987.
[13] 李师翁,马虹,屠骊珠.白刺胚乳早期发育的超微结构研究[J].云南植物研究,1992,14(2):151-156.
[14] 李师翁,屠骊珠.白刺属的胚胎学特征及其系统学意义[J].植物研究,1994,14(3):255-262.
[15] Cruden R W.Pollen-ovule ratios:a conservative indicator of breeding systems in flowering plants[J].Evolution,1977,31(1):32-46.
[16] Dafni A.Pollination ecology:a practical approach[M].Oxford:Oxford University Press,1992.
[17] 张大勇.植物生活史进化与繁殖生态学[M].北京:科学出版社,2004.
[18] Grant V.Plant speciation[M].2nd.New York:Columbia University Press,1981.
[19] 方炎明.森林植被的显花、传粉与繁育系统[J].南京林业大学学报:自然科学版,2012,36(6):1-7.
[20] 赵学杰,谭敦炎,李新蓉.两种鹤虱属短命植物的繁育系统及其对荒漠环境的适应[J].中国沙漠,2008,28(2):294-300.
[21] 潘成臣,刘林德,侯月利,等.黑河中游花棒的开花特性与繁育系统研究[J].中国沙漠,2010,30(5):1099-1103.
[22] Chang S M,Rausher M D.The role of inbreeding depression in maintaining the mixed mating system of the common morning glory,Ipomoea purpurea[J].Evolution,1999,53(5):1366-1376.
[23] Chang S M,Rausher M D.Frequency-dependent pollen discounting contributes to maintenance of a mixed-mating system in the common morning glory Ipomoea purpurea[J].American Naturalist,1998,152(5):671-683.
[24] 陈敏,刘林德,张莉,等.黑河流域中游罗布麻传粉生态学特性研究[J].西北植物学报,2011,31(4):0677-0682.
[25] 张大勇,姜新华.植物交配系统的进化资源分配对策与遗传多样性[J].植物生态学报,2001,25(2):130-141.
[26] Barrett S C H,Jesson L K,Baker A M.The evolution and function of stylar polymorphisms in flowering plants[J].Annals of Botany,2000,85(Supp 1):253-265.
[27] Parachnowitsch A L,Elle E.Variation in sex allocation and male-female trade-offs in six populations of Collinsia parviflora(Scrophulariaceae s.l.)[J].American Journal of Botany,2004,91(8):1200-1207.
[28] Galloni M,Podda L,Vivarelli D,et al.Pollen presentation,pollen-ovule ratios,and other reproductive traits in Mediterranean Legumes(Fam.Fabaceae-Subfam.Faboideae)[J].Plant Systematics and Evolution,2007,266(3/4):147-164.
