我国南亚热带几种人工林生态系统碳氮储量
2013-08-02王卫霞史作民刘世荣卢立华明安刚于浩龙
王卫霞,史作民,*,罗 达,刘世荣,卢立华,明安刚,于浩龙
(1.中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态环境重点实验室,北京 100091;2.中国林业科学研究院热带林业实验中心,凭祥 532600)
20世纪80年代,在我国热带和亚热带地区发起了大规模的造林运动[1],对退化荒坡进行生态恢复,在此过程中,大量的针叶树种作为先锋树种被用于营造人工林[2-3]。然而,这些主要以马尾松(Pinus massoniana)和杉木(Cunninghamia lanceolata)为主的单一物种的针叶林容易加速土壤酸化,降低生物多样性和土壤肥力,并且易受病虫害的攻击[4]。为了减少这些针叶人工纯林所带来的不利影响,许多不同功能型的树种(如速生树种和乡土珍贵树种,固氮树种和非固氮树种等)被用于改建人工林经营模式。而且越来越多的研究表明,不同树种对于土壤性质的影响是不同的,尤其是固氮与非固氮树种之间以及乡土树种与外来树种之间[5-7]。
在我国广西凭祥地区,许多乡土阔叶树种,如格木(Erythrophleum fordii)、红椎(Castanopsis hystrix)、西南桦(Betula alnoides)、米老排(Mytilaria laosensis)等用于人工林的改建,这些乡土阔叶树种有利于增加森林生态系统的生物多样性和生态系统服务功能[8-9]。除此之外,一些外来速生树种,如桉树(Eucalyptus)和相思(Acacia)等也用于森林恢复。但对于这些不同功能型树种用于改建人工林后,它们所在生态系统各组分碳氮储存及分配格局的研究还比较缺乏[10-12]。而碳和氮是构成森林生态系统组分、维持养分循环和影响森林生态功能发挥的两个重要元素,其循环过程及其相互作用,对生态系统的生产力、固碳潜力以及稳定性都具有关键的影响作用[13]。本文以南亚热带不同功能型人工林生态系统为对象,研究其碳氮储量及其分配格局,以揭示碳氮储存、分配格局与不同功能型树种之间的相互关系,旨在为该区人工林营建中的树种选择、人工林生态系统生产力和固碳潜力、以及人工碳汇林的经营管理研究提供科学参考。
1 研究地点与方法
1.1 试验地概况
研究地点位于广西西南边陲的凭祥市境内中国林业科学研究院热带林业实验中心白云实验林场(22°06'N,106°46'E)。该地区位于南亚热带季风气候区域内的西南部,属湿润半湿润气候。境内日照充足,降雨充沛,干湿季节分明;年均气温为20.5—21.7℃,平均月最低温度12.1℃,平均月最高温度26.3℃;年平均降水1400 mm,主要发生在每年4—9月份;≥10℃活动积温6000—7600℃。主要地貌类型以低山丘陵为主,地带性土壤以红壤为主,主要由花岗岩风化形成[14]。
选取格木、红锥和马尾松3种人工林为研究对象。格木人工林林冠较开阔,叶量较低,部分阳光可直达林下层,林下草本灌木较多,主要以杜茎山(Maesa japonica)、玉叶金花(Mussaenda pubuscens)、五节芒(Miscanthus floridulus)等为优势,盖度约70%;红椎人工林林冠密闭,郁闭度很大,极少有直射光到达林下,林下极少有灌草植物(盖度约<10%),多以红椎更新幼苗为主,但落叶量很大,地表有很厚的未分解半分解落叶层;马尾松林林冠不密闭,林下有致密的草本植物,主要以蔓生莠竹(Microstegium vagans)、铁芒箕(Dicranopteris dichotoma)为优势,盖度约90%。3种人工林均是在马尾松人工林皆伐炼山后、经块状整地营建的人工纯林。林分基本情况见表1。

表1 实验林地概况Table 1 Status of experimental plantations
1.2 研究方法
1.2.1 标准地建立与乔木生物量测算
在3种人工林中按坡位上、中、下分别随机设置5个20 m ×20 m的标准地。2011年10月(树木生长末期),对各标准地内的林木进行每木检尺,起测径级为5 cm,测定并计算林分的平均树高、平均胸径和密度,其中格木林和马尾松林根据每木检尺结果,按平均胸径和平均树高选取标准木10株进行生物量测定。样木伐倒后,地上部分按分层切割法分器官测定生物量,地下部分则采用全根挖掘法测定生物量[12,14]。红椎林则是根据刘恩等[10]所建立的生物量预测模型直接估算林分叶、枝、干和根系等器官的生物量。
1.2.2 林下植被生物量、凋落物量的测定
在每个标准地内按梅花形布点设置2 m×2 m、1 m×1 m小样方各5块,采用“样方收获法”测定生物量[15]。灌木层(2 m×2 m)和草本层(1 m×1 m)植物按地上部分和地下部分分别称其鲜重,凋落物(1 m×1 m)则取其混合物称鲜重,选取样品,在80℃烘箱中烘干至恒重,计算其生物量。
1.2.3 样品的采集
2011年10月在对标准地林分生物量测定的同时,分别采集乔木层不同器官(叶、枝、干和根系)、灌木层、草本层和凋落物层样品,经烘干、粉碎、过筛后,进行碳氮含量的测定。
在各林分的每个标准地内,按梅花形布点设置5个土壤采样点,沿土壤剖面按0—10 cm、10—30 cm、30—50 cm、50—75 cm、75—100 cm分层采集土壤样品,把相同标准地内同一层次土壤样品按质量比例混匀后带回实验室自然风干,粉碎过筛后进行碳氮含量的测定;同时在采集土壤剖面样品时用100 cm3环刀取样,105℃下烘干至恒重,测定土壤容重。
1.2.4 碳氮含量的测定
植物和土壤样品中碳含量采用重铬酸钾-水合加热法测定,全氮含量采用凯氏法测定[16]。
1.2.5 碳氮储量的测算
植被部分(包括乔木层、灌木层、草本层和凋落物层)碳氮储量采用每部分生物量与其碳、氮含量之积进行计算[17]。
100 cm土层碳氮储量S(g/cm2)采用以容重BD(g/cm3)、C或N含量C(%)及土层厚度T(cm)进行计算,其计算公式为[18]:

人工林生态系统碳氮储量则是根据乔木层、灌木层、草本层、凋落物层和土壤层碳氮储量之和进行计算。
1.2.6 数据处理
所有数据采用SPSS18.0和Sigmaplot10.0进行处理分析,不同人工林生态系统各结构层次碳氮含量、储量差异显著性采用一元方差及LSD多重比较进行分析,所有数据均呈正态分布。
2 结果与分析
2.1 生态系统不同结构层次的碳氮含量
2.1.1 乔木层各器官碳氮含量
由表2可以看出,在不同器官中,C含量在48.58%—56.61%之间,且其分布与器官年龄(本研究主要指不同器官,如幼嫩器官叶、枝;老化器官干、根等)的关系不明显;N含量变幅为0.16%—3.62%,树叶中氮含量最高,且固氮树种格木不同器官的氮含量均显著高于红椎和马尾松(P<0.05),且幼嫩器官中的氮含量显著大于老化器官(P<0.05),说明氮含量与树木器官年龄和树种生理特性的关系较为明显[19]。从C/N的变化情况来看,老化器官中的C/N比值大于幼嫩器官。各林分乔木层不同器官中的C/N比值的大小为:马尾松>红椎 >格木,反映出非固氮树种的C/N比值大于固氮树种,针叶林的C/N比值大于阔叶林的一般特征。
2.1.2 林下植被和凋落物层碳氮含量
不同人工林林下植被其组成物种不同,其有机物合成和积累能力也不同,因而碳氮平均含量也存在一定的差异。从表3可以看出,不同林分下灌木层的碳氮平均含量在43.22%—49.51%、0.58%—2.02%之间;草本层在35.94%—48.80%、0.64%—2.02%之间;凋落物层在46.19%—50.91%、0.97%—1.96%之间,其中灌木层和草本层的地上部分碳氮含量均较地下部分高,格木林林下植被层氮含量较其它两种林分林下植被层高。总体而言,不同林分林下植被层碳氮含量与各自乔木层碳氮含量的分配规律基本一致。

表2 不同林分乔木层各器官碳氮含量Table 2 The concentration of C and N in different organs of tree layers under different plantations(mean±SE,n=5)

表3 林下植被、地表现存凋落物碳氮含量Table 3 The concentration of C and N in understory vegetation and ground litter(mean±SE,n=5)
总的来看,3种人工林生态系统植被层各层次碳含量有较明显的层次规律。同一林分中各层次平均碳含量均是乔木层 >灌木层 >草本层。表现出随植物个体高度或组织木质化程度的降低,其碳素含量相应减少的趋势[20]。而氮素含量则与其不一致,说明氮含量与器官年龄的关系较为明显(P<0.05)。
2.1.3 土壤层碳氮含量
由表4可以看出,在格木、红椎和马尾松林分中,0—10 cm表层土有机碳及全氮含量显著高于下层土层含量(P<0.05),但林分之间除表土层有机碳的含量差异不显著外,其它层次均表现为差异显著(P<0.05);全氮含量则表现为固氮树种与非固氮树种人工林之间差异显著(P<0.05)。无论有机碳还是全氮均随土壤深度的增加相应降低。对3种林分土壤全氮和有机碳含量进行相关分析表明,3种林分土壤全氮和有机碳含量之间存在显著的线性正相关(相关系数均为r2>0.95)。
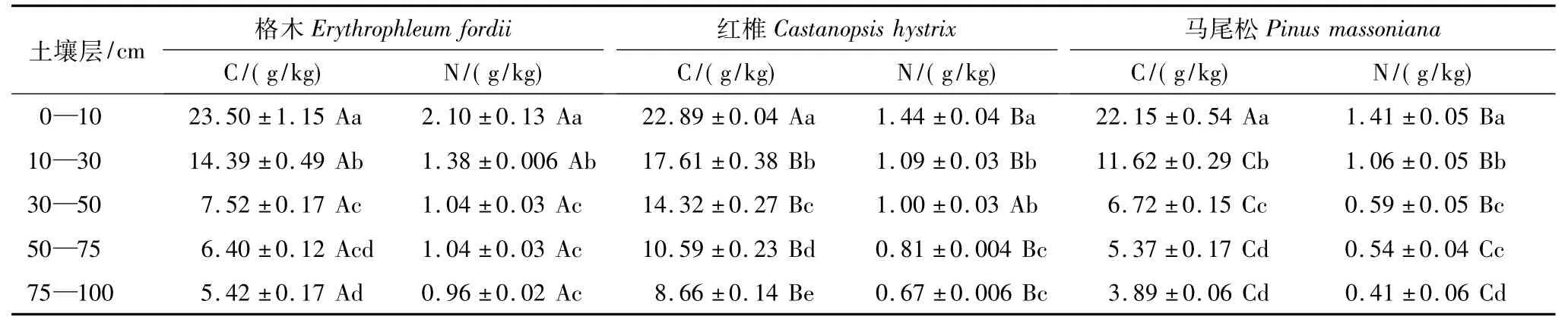
表4 人工林不同土壤层碳氮含量Table 4 The concentration of C and N in different soil layers of the plantations(mean±SE,n=5)
不同林分土壤全氮含量大小依次为格木 >红椎 >马尾松,而不同林分土壤有机碳的含量则与全氮含量不一致,除表层(0—10 cm)土壤外,其它层次不同林分土壤有机碳的含量均表现为红椎 >格木 >马尾松,且红椎林土壤碳含量显著高于格木林和马尾松林(P<0.05)。
2.2 生态系统碳氮储量及分配格局
2.2.1 植被和凋落物层碳氮储量及分配格局
从图1、图2可以看出,乡土固氮树种格木人工林生态系统植被层碳储量要高于乡土阔叶树种红椎林和针叶树种马尾松林。且3种人工林生态系统植被层碳氮储量均表现为差异显著(P<0.05)。其中,格木人工林生态系统植被层碳储量为102.30 t/hm2,红椎人工林生态系统植被层碳储量比格木人工林减少了4.33%,而马尾松减少的幅度则较大,达18.33%。植被层氮储量表现与碳储量基本一致,格木人工林生态系统植被层氮储量最高,为1.34 t/hm2,红椎次之,为0.72 t/hm2,马尾松仅为0.45 t/hm2。
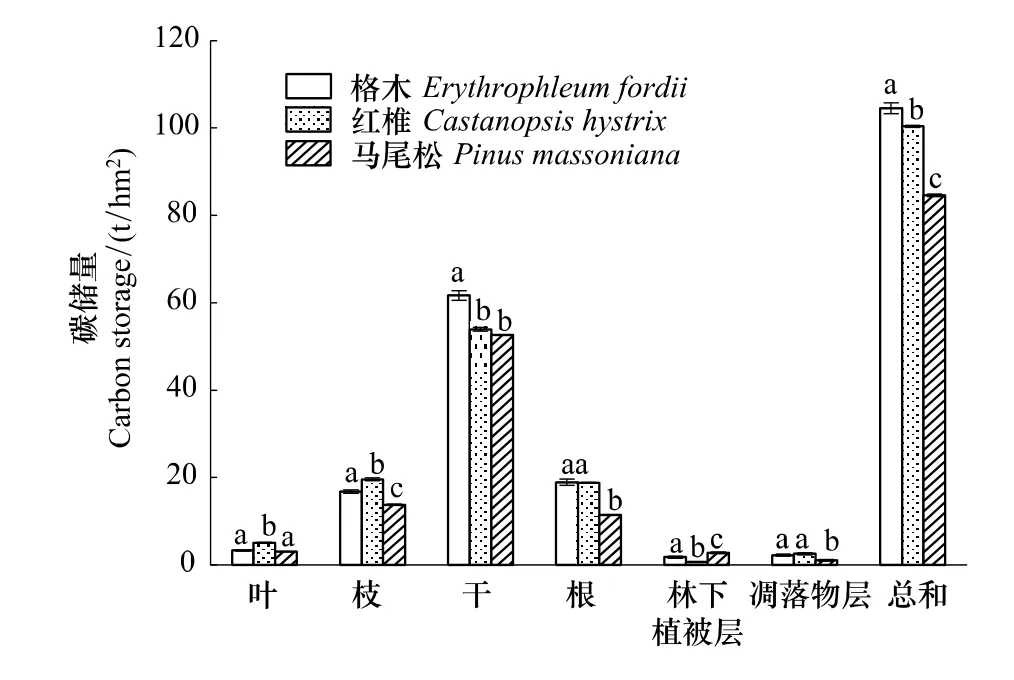
图1 不同林分植被层各组分碳储量Fig.1 C storage of different components in different plantations

图2 不同林分植被层各组分氮储量Fig.2 N storage of different components in different plantations
从各林分植被碳储量的分配格局来看,不同林分各结构层次碳储量的差异程度有所不同。乔木层作为森林生态系统重要组成部分,其碳储量占整个植被碳储量的95%以上,而树干又是乔木层碳储量的主体,其碳储量占乔木层碳储量的55.44%—65.06%;从各林分植被氮储量的分配格局来看,植被氮储量同样主要集中在乔木层,占整个植被氮储量的84%以上,在乔木层各个器官中,氮储量的分配比例并不像碳储量分配的那么集中,相对来说比较分散,但仍以树干氮储量为主体,在各林分植被氮储量的29.28%—39.25%之间波动,占乔木层氮储量的比例处于29.69%—41.09%之间。
林下灌草以及凋落物对森林土壤的理化性质和微生物活性具有重要的调节作用[21]。因此,森林的林下植被和凋落物现存量的变化对土壤碳氮储量具有一定的影响。从图1、图2可以看出,格木林、红椎林和马尾松林林下植被层和凋落物层碳储量分别为1.74、0.62、2.76 t/hm2和2.18、2.52、1.02 t/hm2。而氮储量则分别为0.064、0.013、0.073 t/hm2和0.093、0.054、0.020 t/hm2。由此可见,红椎林下主要以凋落物层碳氮储量为主,马尾松林下主要以灌草碳氮储量为主,格木林下灌草和凋落物碳氮储量相当。这种差异主要是由不同树种的生理特性以及各自林下的环境特征所引起的。
从3种林分植被层碳氮储量的分配格局来看,3种林分林下灌草和凋落物层碳储量占林分植被层碳储量的比例较小,分别为0.06%—3.30%和1.23%—2.58%;而氮储量占林分植被层氮储量的比例则相对较大,分别为1.83%—16.29%和4.47%—7.54%。
2.2.2 土壤层碳氮储量及分配格局
从图3和图4可以看出,红椎林的土壤碳储量要高于格木林和马尾松林,而格木林土壤氮储量高于红椎林和马尾松林。不同林分土壤总碳氮储量存在显著差异(P<0.05)。格木、红椎、马尾松3种人工林土壤碳储量分别为 131.73、167.45 t/hm2和 115.99 t/hm2,氮储量则分别为 16.48、11.61 t/hm2和 10.11 t/hm2。

图3 土壤有机碳储量Fig.3 Organic C storage of soil in different soil depths
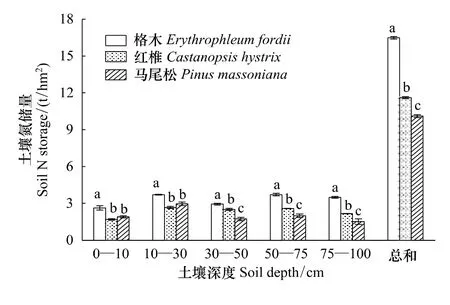
图4 土壤氮储量Fig.4 N storage of soil in different soil depths
从林地土壤碳氮储量的分配格局来看,3种不同林分土壤碳氮储量的分配格局基本一致。林地表层(0—10 cm)土壤碳氮储量分别占土壤总碳氮储量的15.97%—25.55%和14.51%—18.91%,10—30 cm土层碳氮储量则分别占土壤总碳氮储量的25.76%—29.39%和22.53%—29.31%,由此可以看出,0—30 cm土层碳氮是土壤碳氮储量的主体,占土壤碳氮总储量的41.73%—53.66%和37.48%—48.21%。
2.2.3 生态系统碳氮储量及其分配格局
由表5可见,乡土阔叶树种红椎林生态系统总碳储量要高于格木林和马尾松林,具体表现为:红椎林(267.84 t/hm2)>格木林(236.22 t/hm2)>马尾松林(200.57 t/hm2)。生态系统氮储量则以乡土固氮树种格木林生态系统最高,其次是乡土阔叶树种红椎林,针叶树种马尾松林最小。
3种人工林生态系统碳氮储量的分配格局基本一致,红锥和格木人工林生态系统均是土壤层 >乔木层>凋落物层 >林下植被层,马尾松人工林生态系统是土壤层 >乔木层>林下植被层 >凋落物层。其中土壤碳库占整个人工林生态系统总碳库的比例分别为55.77%、62.52%和57.83%,分别是乔木层的1.31倍、1.72倍和1.44倍;而土壤层氮储量占绝对优势,一般是总量的92.00%—95.53%,其次是乔木层(3.59%—7.12%)。由此可见,3种人工林生态系统碳氮储量主要集中在土壤层。

表5 人工林生态系统碳氮储量及其分配格局Table 5 The C and N storage and distribution pattern in different plantation ecosystems
3 讨论与结论
在人工林生态系统中,不同林分乔木层各器官碳的分布与器官年龄的关系并不明显,说明作为构建植物器官的必要元素的碳具有相对稳定性。而不同器官组织氮含量的差异则与器官年龄关系较为密切,表现为幼嫩器官中的氮含量大于老化器官,老化器官中的C/N比值大于幼嫩器官,这与刘增文等[19]和项文化等[22]的研究结果基本一致,另外谌小勇和潘维俦[23]对会同县18年生杉木人工林生态系统中氮素的动态特征的研究也表明枝叶的氮含量与枝叶的龄级成反比,说明氮具有流动性,并且氮素对于幼嫩器官的生长具有重要作用。
土壤有机碳、氮含量主要受动植物残体、凋落物、植物根系、降雨和微生物分解作用以及树种等的影响,并且在一定程度上,土壤氮素的水平也会影响土壤中有机碳的含量[24]。本研究中树种不同,其土壤有机碳和全氮含量具有显著差异,表土层有机碳和全氮含量均是乡土固氮树种格木林表现为最高,且高于广东鹤山外来固氮树种马占相思林(Acacia mangium)[25],乡土阔叶树种红椎林土壤有机碳含量则高于鹤山乡土林木荷林(Schima superba)[25]。说明乡土阔叶树种在本研究区是最具有潜力的土壤改良树种。
格木林、红椎林和马尾松林生态系统碳氮储量的分配格局均是土壤层 >乔木层 >凋落物层。这种分配格局与黄宇等[26]、Finér等[17]的研究结果基本一致。另据阮宏华等人[27]对亚热带苏南地区不同森林类型地上部分与地下部分碳储量之比的研究发现,40年生栎林为1∶1.1,27年生杉木林为1∶1.2;而黄宇等[26]对22年生人工林的研究结果为:杉木纯林1∶1.53,杉木-火力楠(Michelia macclurei)混交林1∶1.55,火力楠纯林1∶2.0。本研究中几种人工林生态系统地上部分与地下部分碳储量之比格木林为1∶1.76,红椎林为1∶2.28,马尾松林为1∶1.74。显然广西亚热带地区这几种人工林类型地下部分碳储量与地上部分碳储量之比相对较高。说明这几种人工林生态系统地下部分碳储存能力较高。同时,红椎林生态系统碳储量高于我国森林生态系统平均碳储量258.83 t/hm2[28],说明红椎林生态系统固碳能力较高,尤以地下部分土壤层的高固碳能力为主。
森林土壤是森林生态系统的一个极重要的碳氮存储库,在平衡温室气体中发挥重要的作用[29]。本研究中,格木林土壤层碳储量高于广东鹤山20年生马占相思林土壤层碳储量114.62 t/hm2[25],红椎林土壤层碳储量略低于广西27年生红椎林土壤层碳储量[10]。3种人工林土壤层碳储量均低于我国森林土壤平均碳密度193.55 t/hm2[28],其主要原因在于受亚热带良好水热条件的影响,亚热带土壤呼吸速率较大,加之植被对土壤中养分的大量吸收,造成土壤碳素积累较少[28]。而两种阔叶林土壤层氮储量则高于我国亚热带常绿阔叶林区域土壤层氮密度10.17 t/hm2,其中红椎林和马尾松林土壤层氮储量则略低于我国热带雨林季雨林区域土壤层氮密度13.64 t/hm2[30]。因此说明森林生态系统土壤层碳氮储量受土壤质地、树种和植被类型、气候带以及林龄等多个因素的影响[17-18]。
根据方晰等人[31]对湖南会同22年生的杉木林地土壤碳储量的研究,杉木林地0—30 cm土层碳储量占土壤总碳储量的53.52%,Batjes[32]对全球各类土壤C贮量的研究认为,0—30 cm土层碳储量占土壤总碳储量的比例平均为49%。另根据黄宇等人[26]对不同人工林土壤氮储量的研究,林地土壤0—40 cm土层氮储量占0—100 cm的比例平均为55.50%。本区格木林、红椎林和马尾松林土壤碳氮储量分配格局的研究结果表明,不同林分0—30 cm土层碳储量分别占土壤总碳储量的51.73%、41.73%和53.66%,而0—30 cm土层氮储量分别占土壤总氮储量的38.48%、37.48%和48.21%,与其它地区的研究结果基本一致。另外从结果中还可以看出马尾松林0—30 cm土层碳氮储量占土壤总碳氮储量的比例较高,说明格木林和红椎林深层土壤(30—100 cm)比马尾松林深层土壤具有更高的碳储存能力和养分供应能力,这也说明乡土阔叶树种更有利于土壤肥力的保持。
[1]Li Z A,Peng S L,Rae D J,Zhou G Y.Litter decomposition and nitrogen mineralization of soils in subtropical plantation forests of southern China,with special attention to comparisons between legumes and non-legumes.Plant and Soil,2001,229(1):105-116.
[2]Peng S L,Hou Y P,Chen B M.Vegetation restoration and its effects on carbon balance in Guangdong Province,China.Restoration Ecology,2009,17(4):487-494.
[3]Ren H,Shen W J,Lu H F,Wen X Y,Jian S G.Degraded ecosystems in China:status,causes,and restoration efforts.Landscape and Ecological Engineering,2007,3(1):1-13.
[4]Yu Z Y,Peng S L.Ecological studies on vegetation rehabilitation of tropical and subtropical degraded ecosystems.Guangzhou:Guangdong Scienceand Technology Press,1996.
[5]Lal R.Forest soils and carbon sequestration.Forest Ecology and Management,2005,220(1/3):242-258.
[6]Binkley D,Giardina C,Bashkin M A.Soil phosphorus pools and supply under the influence of Eucalyptus saligna and nitrogen-fixing Albizia facaltaria.Forest Ecology and Management,2000,128(3):241-247.
[7]Russell A E,Raich J W,Valverde-Barrantes O J,Fisher R F.Tree species effects on soil properties in experimental plantations in tropical moist forest.Soil Science Society of America Journal,2007,71(4):1389-1397.
[8]Carnevale N J,Montagnini F.Facilitating regeneration of secondary forests with the use of mixed and pure plantations of indigenous tree species.Forest Ecology and Management,2002,163:217-227.
[9]Liang R L.Current situation of Guangxi indigenous broadleaf species resource and their development counter-measures.Guangxi Forestry Science,2007,36(1):5-9.
[10]Liu E,Wang H,Liu S R.Characteristics of carbon storage and sequestration in different age beech(Castanopsis hystrix)plantations in south subtropical area of China.Chinese Journal of Applied Ecology,2012,23(2):335-340.
[11]Ming A G,Jia H Y,Tao Y,Lu L H,Su J M,Shi Z M.Biomass and its allocation in a 28-year-old Mytilaria laosensis plantation in southwest Guangxi.Chinese Journal of Ecology,2012,31(5):1050-1056.
[12]He B,Liu Y H,Yu H G,Qin W M,Wei S H,Cai S W.Carbon density and storage of Acacia mangium plantation ecosystem in Nanning,Guangxi.Scientla Silvae Sinicae,2009,45(2):6-11.
[13]Reich P B,Hobbie S E,Lee T,Ellsworth D S,West J B,Tilman D,Knops J M H,Naeem S,Trost J.Nitrogen limitation constrains sustainability of ecosystem response to CO2.Nature,2006,440(7086):922-925.
[14]Kang B,Liu S R,Zhang G J,Chang J G,Wen Y G,Ma J M,Hao W F.Carbon accumulation and distribution in Pinus massoniana and Cunninghamia lanceolata mixed forest ecosystem in Daqingshan,Guangxi of China.Acta Ecologica Sinica,2006,26(5):1320-1329.
[15]Li H T,Wang S N,Gao L P,Yu G R.The carbon storage of the subtropical forest vegetation in central Jiangxi Province.Acta Ecologica Sinica,2007,27(2):693-704.
[16]Liu G S.Observation and standard analysis method of Chinese Ecosystem Research Network:Physiochemical analysis and profile description of soil.Beijing:China Standard Press,1996.
[17]Finér L,Mannerkoski H,Piirainen S,Michael S.Carbon and nitrogen pools in an old-growth,Norway spruce mixed forest in eastern Finland and changes associated with clear-cutting.Forest Ecology and Management,2003,174(1/3):51-63.
[18]Luo T S,Chen B F,Chen Y F,Yang Y C,Yang X S,Li D J.Variation of the soil carbon and nitrogen for initial stage after the felling in tropical montane rainforest of Bawangling,Hainan Island.Forest Research,2000,13(2):123-128.
[19]Liu Z W,Wang N J,Li Y S,Lü Y L.Nutritional principle of forest ecosystem stability.Journal of Northwest Sci-tech University of Agriculture and Forestry(Natural Science),2007,34(12):129-134.
[20]Li M H,Yu M J,Chen Q C,Chang J,Pan X D.Dynamics of carbon in the evergreen broadleaved forest dominated by Cyclobalanopsis glauca in south-east China.Acta Ecologica Sinica,1996,16(6):645-651.
[21]Yang Y S,Guo J F,Lin P,He Z M,Xie J S,Chen G S.Carbon and nutrient pools of forest floor in native forest and monoculture plantations in subtropical China.Acta Ecologica Sinica,2004,24(2):359-367.
[22]Xiang W H,Tian D L.Nutrient cycling in pinus massoniana stands of different age classes.Acta Phytoecologica Sinica,2002,26(1):89-95.
[23]Chen X Y,Pan W C.Dynamic properties of nitrogen in the Chinese fir plantation ecosystem.Acta Ecologica Sinica,1989,9(3):201-206.
[24]Su Y Z,Zhao H L.Advances in researches on soil organic carbon storages,affecting factors and its environmental effects.Journal of Desert Research,2002,22(3):220-228.
[25]Li Y L,Hu C Z,Zhang Y,Wen J Z.Studies of the carbon storages of soils under the different kinds of plantations.Journal of Fujian Forestry Science and Technology,2004,31(4):4-7.
[26]Huang Y,Feng Z W,Wang S L,Feng Z Z,Zhang H X,Xu Y R.C and N stocks under three plantation forest ecosystem of Chinese-fir,Michelia macclurei and their mixture.Acta Ecologica Sinica,2005,25(12):3146-3154.
[27]Ruan H H,Jiang Z L,Gao S M.Study on carbon cycling of major forest types in South Jiangsu:content and distribution.Chinese Journal of Ecology,1997,16(6):17-21.
[28]Zhou Y R,Yu Z L,Zhao S D.Carbon storage and budget of major Chinese forest types.Acta Phytoecologica Sinica,2000,24(5):518-522.
[29]Thompson J A,Kolka R K.Soil carbon storage estimation in a forested watershed using quantitative soil-landscape modeling.Soil Science Society of America Journal,2005,69(4):1086-1093.
[30]Zhang C N,Yan X D,Yang J H.Estimation of nitrogen reserves in forest soil of China.Journal of Southwest Agricultural University,2004,26(5):572-575.
[31]Fang X,Tian D L,Xiang W H.Effects of different management patterns on soil carbon storage of the deforested lands in Chinese fir plantation.Journal of Central South Forestry University,2004,24(1):1-5.
[32]Batjes N H.Total carbon and nitrogen in the soils of the world.European Journal of Soil Science,1996,47(2):151-163.
参考文献:
[4]余作岳,彭少麟.热带亚热带退化生态系统植被恢复生态学研究.广州:广东科技出版社,1996.
[9]梁瑞龙.广西乡土阔叶树种资源现状及其发展对策.广西林业科学,2007,36(1):5-9.
[10]刘恩,王晖,刘世荣.南亚热带不同林龄红锥人工林碳贮量与碳固定特征.应用生态学报,2012,23(2):335-340.
[11]明安刚,贾宏炎,陶怡,卢立华,苏建苗,史作民.桂西南28年生米老排人工林生物量及其分配特征.生态学杂志,2012,31(5):1050-1056.
[12]何斌,刘运华,余浩光,秦武明,韦善华,蔡树威.南宁马占相思人工林生态系统碳素密度与贮量.林业科学,2009,45(2):6-11.
[14]康冰,刘世荣,张广军,常建国,温远光,马姜明,郝文芳.广西大青山南亚热带马尾松,杉木混交林生态系统碳素积累和分配特征.生态学报,2006,26(5):1320-1329.
[15]李海涛,王珊娜,高鲁鹏,于贵瑞.赣中亚热带森林植被碳储量.生态学报,2007,27(2):693-704.
[16]刘光崧主编.中国生态系统研究网络观测与分析标准方法——土壤理化分析与剖面描述.北京:中国标准出版社,1996.
[18]骆土寿,陈步峰,陈永富,杨彦臣,杨秀森,李大江.海南岛霸王岭热带山地雨林采伐经营初期土壤碳氮储量.林业科学研究,2000,13(2):123-128.
[19]刘增文,王乃江,李雅素,吕月玲.森林生态系统稳定性的养分原理.西北农林科技大学学报:自然科学版,2007,34(12):129-134.
[20]李铭红,于明坚,陈启瑺,常杰,潘晓东.青冈常绿阔叶林的碳素动态.生态学报,1996,16(6):645-651.
[21]杨玉盛,郭剑芬,林鹏,何宗明,谢锦升,陈光水.格氏栲天然林与人工林枯枝落叶层碳库及养分库.生态学报,2004,24(2):359-367.
[22]项文化,田大伦.不同年龄阶段马尾松人工林养分循环的研究.植物生态学报,2002,26(1):89-95.
[23]谌小勇,潘维俦.杉木人工林生态系统中氮素的动态特征.生态学报,1989,9(3):201-206.
[24]苏永中,赵哈林.土壤有机碳储量,影响因素及其环境效应的研究进展.中国沙漠,2002,22(3):220-228.
[25]李跃林,胡成志,张云,文锦柱.几种人工林土壤碳储量研究.福建林业科技,2004,31(4):4-7.
[26]黄宇,冯宗炜,汪思龙,冯兆忠,张红星,徐永荣.杉木,火力楠纯林及其混交林生态系统C,N贮量.生态学报,2005,25(12):3146-3154.
[27]阮宏华,姜志林,高苏铭.苏南丘陵主要森林类型碳循环研究——含量与分布规律.生态学杂志,1997,16(6):17-21.
[28]周玉荣,于振良,赵士洞.我国主要森林生态系统碳贮量和碳平衡.植物生态学报,2000,24(5):518-522.
[30]张春娜,延晓冬,杨剑虹.中国森林土壤氮储量估算.西南农业大学学报,2004,26(5):572-575.
[31]方晰,田大伦,项文化.不同经营方式对杉木林采伐迹地土壤C储量的影响.中南林学院学报,2004,24(1):1-5.
