内蒙古荒漠草原主要草食动物食性及其营养生态位
2013-08-02刘贵河王国杰汪诗平韩建国宛新荣郝树广
刘贵河,王国杰,汪诗平,* ,韩建国 ,宛新荣,郝树广
(1.河北北方学院动物科技学院,张家口 075000;2.中国科学院青藏高原研究所高寒生态学和生物多样性实验室,北京 100101;3.中国农业大学动物科技学院草业科学系,北京 100193;4.中国科学院动物研究所,北京 100080)
近年来,我国90%的可利用草地处于不同程度退化之中[1],尤其是内蒙古天然草原退化极为严重[2-3]。多数学者认为超载过牧是草地退化的根本原因[4-5]。还有学者认为,草原鼠类频繁地采食优良牧草、挖洞损伤牧草等也是引发草地退化的原因[6-8],尤其在重牧区影响更为严重[9]。此外,蝗虫也是天然草原最主要的食草动物之一,其群落结构和种群数量的变化影响着草原群落结构和功能的发挥,其危害程度取决于牧草的生育期和群落结构[10-11]。总之,食草动物频繁的啃食、践踏和损伤不仅改变了植物群落的结构和种类组成,也影响并决定着群落的演替方向[6,12-13];相反,植被群落的变化也影响着食草动物群落与种群结构的改变[14-18],二者互为因果又相辅相成,这种关系形成了不同放牧强度下草原特有的生态系统。在该系统中,食草动物对食物资源相互竞争,出现生态位重叠与分化现象[19]。有关食草动物食性及生态位研究多集中在同一类物种内[20-24],对不同类食草动物种间生态位变化的研究相对较少[25-26]。而天然草原同时承受着上述多种食草动物的干扰,它们的食性及生态位变化共同决定着草原的演替方向,但这方面的研究未见报道。本研究选择内蒙古荒漠草原小针茅(Stipa klemenzii)群落为研究对象,运用饱和链烷技术[27-31]研究不同放牧强度下,绵羊(Ovis aries)、优势鼠种达乌尔黄鼠(Spermophilus dauricus)、优势蝗虫亚洲小车蝗(Oedaleus asiaticus)食性变化及其营养生态位关系,以揭示天然草原放牧退化演替规律,为植被恢复及草原合理利用提供理论依据。
1 材料与方法
1.1 试验区概况
试验样地选择在内蒙古荒漠草原苏尼特右旗北部小针茅群落中,地处北纬42°46'36″,东经112°40'25″,海拔1100 m,年降雨量平均为200 mm,未退化植物群落种类组成以小针茅、多根葱(Allium bidentatum)和无芒隐子草(Cleistogenes songorica)为主,草群高度为13—17 cm,植被总盖度为35%左右,土壤为棕钙土。试验于2003年7—8月份用链烷技术估测群落中自由放牧绵羊、优势鼠种、优势蝗虫的食性。
1.2 试验设计
1.2.1 自由放牧绵羊食性研究
对天然草原自由放牧的家畜而言,放牧梯度等级的确定最好是根据离开水源的距离来确定[32-33],该试验的放牧强度是以离开村庄或围栏距离的远近来选择不同放牧压力的试验小区,无牧区选择在围封10a的围栏内,中等放牧区和重度放牧区分别选择在距村庄2.0 km和1.0 km的区域,因为当地牧民的家畜晚上都圈于圈中。
1.2.2 达乌尔黄鼠食性研究
采用4号木板夹,在不同牧压区域进行鼠类取样,夹距5 m,行距50 m,共3行,每行100夹,共放置300夹,连续测定3日,在每日的上午和傍晚查夹并取鼠,每40夹的面积为1 hm2,3日连续夹捕数目可视为样地鼠密度。捕获的鼠经熏蒸灭蚤(疫)后,统计鼠的种类和数量,以计算鼠的密度及优势鼠种比例(表1)。试验区主要鼠种有达乌尔黄鼠、黑线毛足鼠(Phodopus sungorus)、五趾跳鼠(Allactaga sibirica)、长爪沙鼠(Meriones unguiculatus)、黑线仓鼠(Cricetulus Barabensis)、短耳仓鼠(Allocricetulus eversmanni)、五趾心颅跳鼠(Cardiocranius paradoxus)等,不同放牧区达乌尔黄鼠数量最多,为优势种(表1)。然后在实验室解剖达乌尔黄鼠并取其胃内含物。

表1 不同放牧强度鼠种的密度及其组成Table 1 The density of mice and their composition under different grazing intensities
1.2.3 亚洲小车蝗食性研究
运用扫网的方法测定草原蝗虫的种类和密度,在不同牧压试验区内随机取20个点,每点向外辐射扫10网,共计200网,统计蝗虫种类与数量,计算蝗虫密度,确定优势蝗虫种并在笼内关养1d,收集其粪便。试验区不同牧压下优势蝗虫均为亚洲小车蝗(表2)。
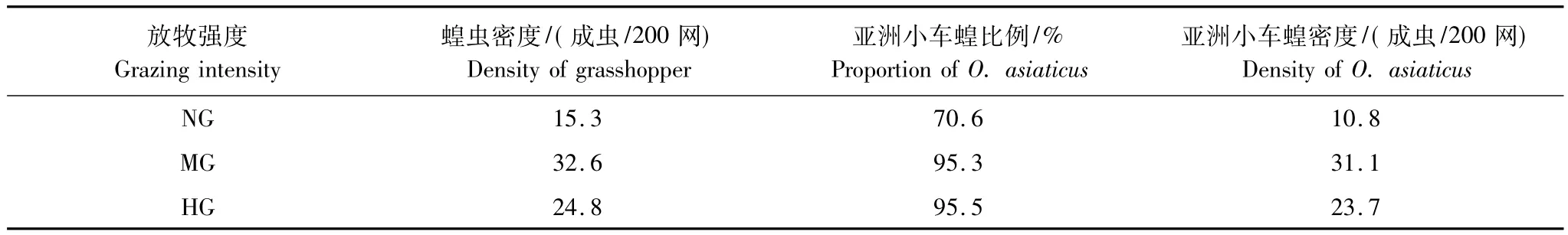
表2 不同放牧强度蝗虫的密度及其优势种比例Table 2 The density of grasshopper and proportion of their dominant species under different grazing intensities
1.3 样品采集和处理
1.3.1 植物样品的采集与处理
在不同放牧压力下的每个试验小区内,沿2条100 m的样线间隔10 m做一个样方(1.0 m×1.0 m),共计20个样方。样方内植物齐地面刈割,分种称量鲜重后,在65℃烘箱中烘干至恒重,称量干物质量。取相对生物量大于0.5%的每种牧草约20—200 g过1 mm筛粉碎,分析其链烷含量。
1.3.2 羊粪样品的采集与处理
同期在相应试验区域随机选择10只绵羊,带上集粪袋,连续3d收集新鲜羊粪并置于65℃烘箱中烘干,然后将每只绵羊的粪便分别混合均匀,粉碎过1 mm筛,分析其链烷含量。
1.3.3 鼠胃内含物的取样与处理
对在试验区内所取的达乌尔黄鼠全部进行解剖(无牧区10只;中度放牧区6只;重度放牧区6只),取胃内含物,并按放牧梯度混合胃内含物,在65℃烘箱中烘干,粉碎过1 mm筛,分析其链烷含量。
1.3.4 蝗虫粪便的取样与处理
通过扫网,取亚洲小车蝗成熟个体约150头(M∶F=1∶1),放置在笼中(1.0 m ×0.5 m ×0.5 m),不加任何食物,每隔6小时收集一次粪便,共收集4次,粪便样品在65℃烘箱中烘干,分析其链烷模式。
1.4 样品分析
植物与粪便样品链烷模式分析方法参见刘贵河等[31]方法进行分析。
1.5 数据处理和分析
1.5.1 采食牧草比例计算
对放牧绵羊、优势鼠种和优势蝗虫的牧草采食比例采用最小平方优化程序Eatwhat软件进行计算[34]。
1.5.2 选择性指数计算
选择性指数(SI)又称偏嗜性指数,是指食物中某种牧草占的重量百分率与群落中该种植物占的重量百
1.5.3 营养生态位计算
食草动物营养生态位宽度采用Shaanon-Wiener信息指数计算,生态位重叠指数采用Hurlbert(1978)重叠指数计算,计算公式分别为,式中Bj为生态位宽度;Oik生态位重叠指数;Pij为i食草动物利用j资源的比例;Pkj为k食草动物利用j资源的比例;Cj为第j个资源状态的相对多度[37]。
1.5.4 数据分析
试验数据采用SPSS11.5软件GLM模型进行二因素及三因素方差分析,应用LSD与T方法进行多重比较,所有数据用Excel计算并形成图表。
2 结果与分析
2.1 群落中植物的种类组成
通过植被样方数据可知,不同放牧强度下小针茅群落中共有14—20种植物,但其中的10种植物地上生物量占群落总生物量的99%以上,它们是食草动物的主要食物来源。其中,优势种小针茅随放牧强度的增加而下降,猪毛菜(Salsola collina)随放牧强度增加而增加,变为优势种(表3),说明小针茅群落随牧压的增大演替为猪毛菜群落。有关放牧强度对植被更详细的影响参见王国杰等[38],这里不再赘述。
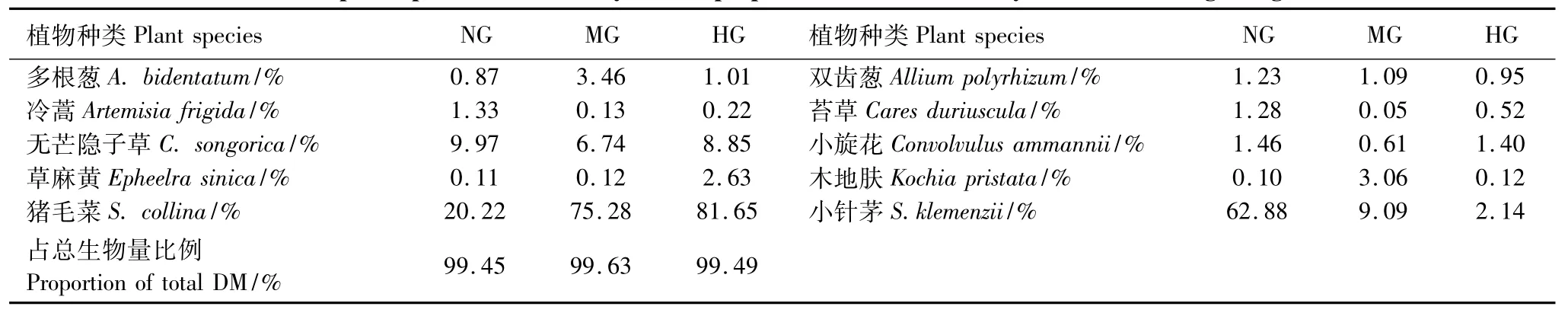
表3 不同放牧强度主要植物种及其干物质比例Table 3 Main plant species and their dry matter proportion in the community under different grazing intensities
2.2 群落中主要植物种链烷模式
小针茅群落中10种主要植物的链烷模式见表4,在这些植物中,奇数碳链的浓度高于相邻偶数碳链浓度,且优势链烷C27、C29、C31和C33模式存在种间差异,链烷浓度总和也存在种间差异。

表4 小针茅群落中主要植物种链烷浓度/(mg/kg干物质)Table 4 n-alkane concentrations of main plant species in S.klemenzii community
2.3 不同放牧强度下3种食草动物的食性
试验区内虽然有10种主要植物种,但由于不同放牧强度下群落组成发生了显著变化(表3),所以导致了不同放牧强度下食草动物采食的牧草种类和数量存在一定的差异(表5)。绵羊主要采食其中的6—8种植物,其中猪毛菜占的比例最大,且在重牧区绵羊增加了对冷蒿(Artemisia frigida)和苔草(Cares duriuscula)的采食;达乌尔黄鼠主要采食其中的5—7种植物,主要以猪毛菜和多根葱为主;亚洲小车蝗主要采食5种植物,其中无芒隐子草、小针茅和猪毛菜采食比例较大,然后是双齿葱(Allium polyrhizum)和多根葱。通过对放牧强度、食草动物和牧草种类3因子进行方差分析,发现每种因子都存在显著的差异,而且两两之间和三者之间都存在极显著的互作(表6)。进一步进行多重比较发现,无牧区野生食草动物采食的牧草比例显著低于放牧区,但中度放牧区与重度放牧区食草动物采食牧草比例差异不显著(表7);绵羊采食牧草比例显著低于达乌尔黄鼠和亚洲小车蝗,但达乌尔黄鼠和亚洲小车蝗的采食比例差异不显著(表8)。不同放牧强度不同食草动物对8种牧草采食的比例除苔草和木地肤(Kochia pristata)外,所有牧草采食比例差异均显著(表9)。
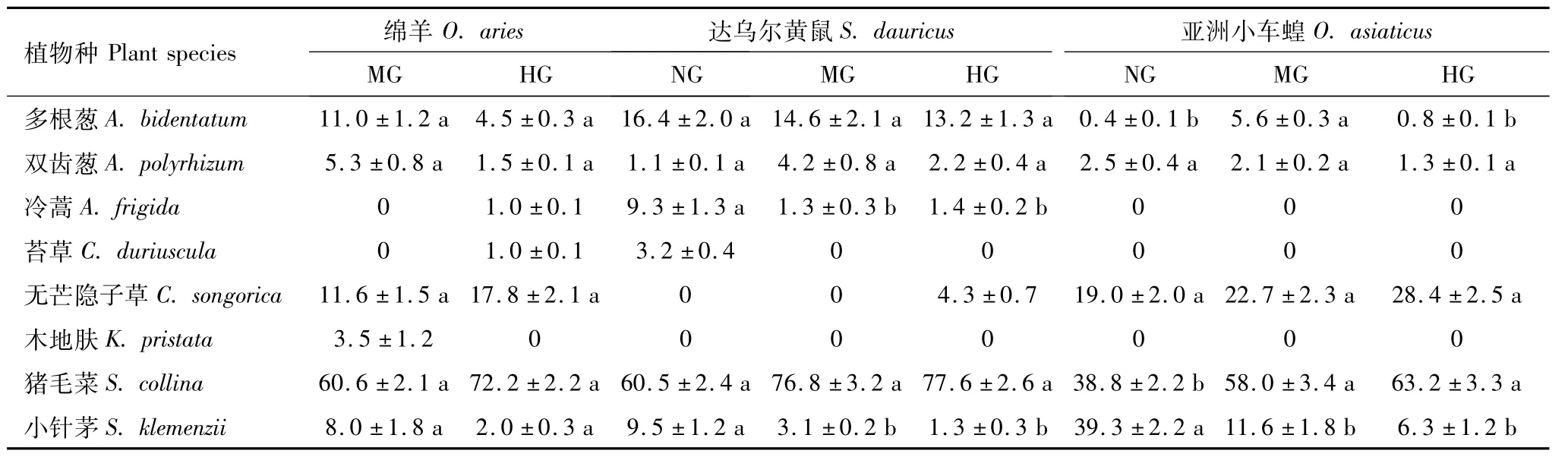
表5 不同放牧强度3种食草动物食性组成及多重比较(平均值±标准误)/%Table 5 Estimated diet composition(Mean±S.E%)and their multiple comparisons of three herbivores using the n-alkane technique under different grazing intensities

表6 牧草采食比例三因子方差分析表Table 6 Three factor analysis of variance of diet proportion

表7 不同放牧强度牧草采食比例多重比较Table 7 Multiple comparisons of diet proportion under different grazing intensities

表8 不同食草动物牧草采食比例多重比较Table 8 Multiple comparisons of diet proportion in different herbivores
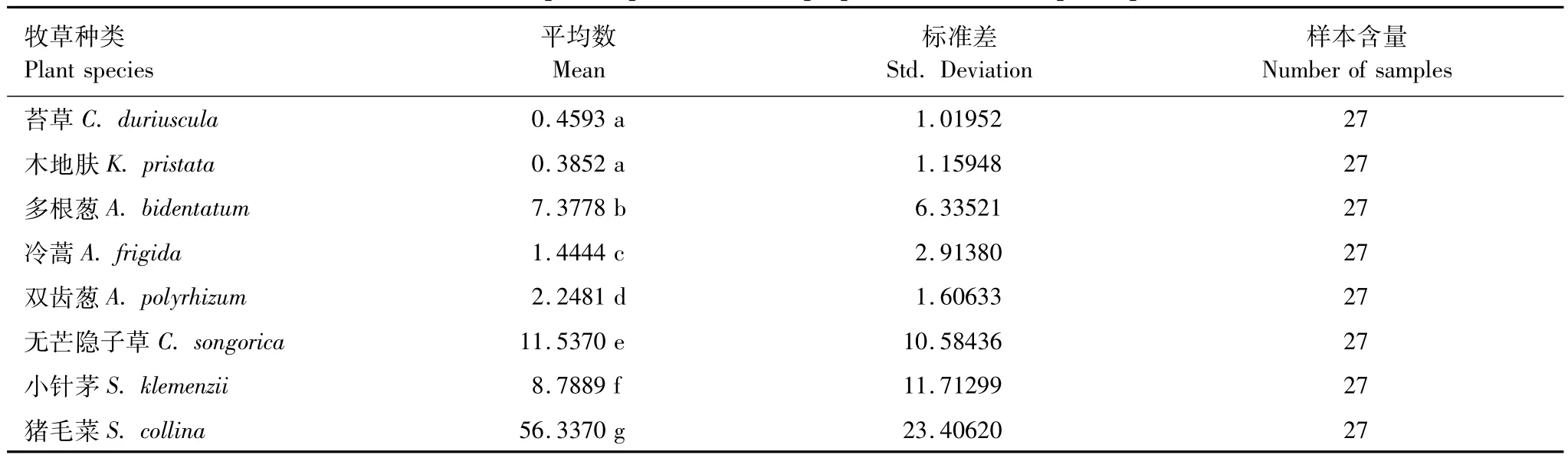
表9 不同牧草种类采食比例多重比较表Table 9 Multiple comparisons of diet proportion in different plant species
2.4 不同放牧强度3种食草动物对牧草的选择性指数
选择性指数大于1.0说明该种植物被优先采食,在不同牧压下,绵羊对牧草的选择性指数不同(表10),但优先采食多根葱、无芒隐子草和双齿葱,其中多根葱和无芒隐子草随牧压的增大选择性指数上升,双齿葱随牧压的增大选择性指数下降,并且在中度放牧区和重牧区,绵羊分别对木地肤、冷蒿和苔草具有很高的选择性,虽然绵羊的选择性采食表现为随牧压变化而变化,但主要取决于群落中可食牧草数量及易接近程度;而达乌尔黄鼠对食物的选择性指数除小针茅和无芒隐子草外均很高(表10),尤其对多根葱的选择性指数最高,随牧压的增大,达乌尔黄鼠对猪毛菜的选择性指数下降,对小针茅的选择性指数增加,也同样说明该鼠种具有很强的选择性采食行为,也同样受牧压的影响较大;随着牧压的增大,亚洲小车蝗对双齿葱和猪毛菜的选择性指数下降,而对无芒隐子草和小针茅选择性指数增加,多根葱在适牧条件下,选择性指数最高(表10),说明亚洲小车蝗对食物的选择性采食受牧压影响也较大。
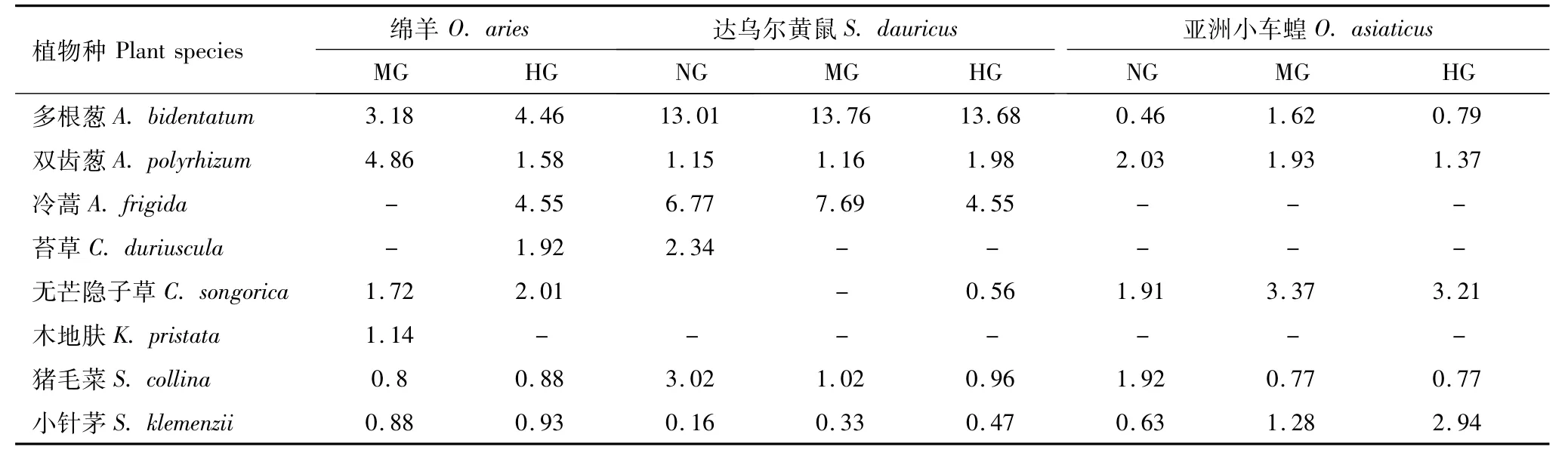
表10 不同放牧强度3种食草动物对牧草的选择性指数(SI)Table 10 Selectivity index(SI)of three herbivores of primary plant species under different grazing intensities
2.5 不同放牧强度3种食草动物营养生态位宽度
3种食草动物在不同放牧压力下,其营养生态位宽度是不一样的(表11),随牧压的增大,绵羊和亚洲小车蝗的营养生态位变窄,而达乌尔黄鼠的生态位在中度放牧区最低,但总的变化趋势也随牧压的增大而变窄。

表11 不同放牧强度3种食草动物营养生态位宽度Table 11 Coefficient of trophic niche breadth of three herbivores under different grazing intensities
2.6 3种食草动物营养生态位重叠度
生态位重叠程度反映了物种间对资源利用的相似程度,同时也反映它们之间潜在的竞争程度。3种食草动物在小针茅群落中实际营养生态位重叠程度较高(表12),其中绵羊和达乌尔黄鼠生态位重叠程度最高,依次为绵羊和亚洲小车蝗、达乌尔黄鼠和亚洲小车蝗生态位的重叠,说明3种食草动物在小针茅群落中对牧草的利用程度相似,竞争较为激烈。结合3种食草动物对牧草的选择性指数(表10)分析,三者对双齿葱和多根葱的竞争最为激烈,其次是绵羊与达乌尔黄鼠对冷蒿的竞争,以及绵羊与亚洲小车蝗对无芒隐子草的竞争,因为猪毛菜在群落中比例较高,竞争表现不明显。
3 讨论
(1)链烷技术理论上可以评价食草动物对15种牧草的采食[28],但是当食草动物采食的牧草种类较多或牧草中链烷含量较低的情况下会增大估测误差[39-40],普遍结论认为,链烷技术可以准确评价食草动物对2—10种牧草的采食比例[29,41]。据植被样方统计,无牧区有20种植物,中度放牧区有14种植物,重度放牧区有17种植物,根据牧草生物量及其出现频率,人为剔除了无牧区中生物量低于0.05%的几种植物或出现频率极少的植物,比如米口袋(Gueldenstaedtia verna)占群落生物量的0.03%、地苕瓜(Cynanchum thesioides)占0.02%、扁蓄豆(Pocockia ruthenia)占0.0005%、乳白花黄芪(Astragalus galactites)占 0.002%、平车前(Plantago depressa)占0.04%、洽草(Koeleria cristata)占0.03%、雾冰黎(Bassia dasyphylla)占0.04%等,类似的植物还有鸦葱(Scorzonera austriaca)、细叶鸢尾(Iris tenuifolin)、天门冬(Asparagus cochinchinensis),而这些植物在放牧区域几乎没有或偶见。因此本研究选择生物量占99%以上的10种代表性植物进行试验,力求结果更为真实。尽管食草动物可能采食了10种牧草以外的其他牧草种类,但由于其在群落中的数量很少,对绵羊和蝗虫的食性影响较小,加之链烷技术本身的缺陷[40],本研究只对其中的10种牧草进行估测,建议以后结合其他方法对食草动物食性进一步展开量化研究。

表12 3种食草动物营养生态位重叠度Table 12 Overlap of trophic niche among three herbivores in S.klemenzii community
(2)放牧家畜食性的变化受很多因素影响,比如,牧草的适口性、物候期、形态学特征及气候、地形等因素[42]。当放牧地牧草资源(地上生物量)丰富时,放牧家畜能够选择喜食的牧草种,反之,家畜就被迫采食先前不喜食的植物种,尤其在重牧区[12,36,43]。本试验结果表明,在重牧区,绵羊对猪毛菜的采食比例增加,这与群落中猪毛菜数量增多有关。当食物资源缺乏或喜食的牧草种类较少时,绵羊通过增加放牧时间来寻找适口性好的牧草或难于接近的牧草[44-45],试验结果显示,在重牧区,绵羊增加了对冷蒿与苔草的采食,而这些牧草在群落中数量极少,此结果也同样说明了绵羊具有很强的选择性采食行为。
(3)放牧不仅改变了草地的植物种类组成[5],进而也改变了鼠的种群结构[7,46],本研究随着牧压的变化鼠的种类及其密度也随之变化(表1),达乌尔黄鼠在无牧区和重度放牧区的密度虽然低于50%,但在群落中其种群密度相对较高,尽管达乌尔黄鼠不足以代表草地啮齿动物类群,但一定程度上也能反映出主要鼠类与放牧家畜之间的营养生态位关系,笔者未能研究全部鼠种的食性,建议在以后研究中逐步完善。啮齿动物食性较为复杂,因为多数啮齿动物除了采食牧草茎叶外,还采食植物根系、牧草种子和无脊椎动物[17,47],所以在分析啮齿动物采食牧草种类时,只能进行粗略的估测,不能确定相对准确的采食比例。但达乌尔黄鼠是以植物性食物为主的鼠种[48],受实验条件和方法的限制,本研究只能粗略估测出达乌尔黄鼠采食了5—6种牧草,主要以杂类草和禾草为主,此结果与王桂明等[21]的研究结果类似,尽管群落中牧草种类和数量随牧压改变发生了巨大的变化,但达乌尔黄鼠仍喜食多根葱、双齿葱和冷蒿,达乌尔黄鼠的这种选择性采食可能与鼠种有关[49],也可能与植物生长季节有关[50]。本研究在重度放牧区黄鼠增加了对无芒隐子草的采食,这与群落中无芒隐子草的生物量无关,可能与群落中喜食牧草种类减少有关。
(4)前人总结了很多的方法来评价蝗虫食性,但大多数方法只能对蝗虫采食的牧草种类进行归类[51],虽然嗉囊内含物显微分析技术被广泛用于评价蝗虫的食性[52],但在实际应用中很难被推广[53],本研究应用链烷技术估测的亚洲小车蝗食性主要以禾草和杂类草为主,此结果与前人的研究结果一致[11]。影响蝗虫食性的因素很多,如气候干旱、高温[54]、植被类型、盖度和草地退化程度[16,20]。本研究发现,亚洲小车蝗的食性随放牧强度的变化差异不显著,说明亚洲小车蝗对食物的选择性较强,受群落多样性的影响较小。
(5)生态位宽度是指一个种群在一个群落中所利用的各种不同资源的总和[37]。当食物资源充足时,食草动物利用最适于其生存和最方便利用的食物资源,导致营养生态位变窄[50],当食物资源短缺时,它们则尽量发挥其资源利用潜力,促使生态位变宽[21]。本试验3种食草动物的生态位随牧压的增大而变窄,对绵羊而言,这不是食物资源充足的结果,而是食物资源唯一性的结果,对鼠和蝗虫而言,食物资源变的丰富。放牧不仅改变了小针茅群落的植物种类组成,而且会加剧鼠虫害的发生,同时也说明小针茅群落已经不适合放牧利用[55]。
(6)不同种群生态位重叠意味着对食物资源存在竞争,尤其在食物资源不足时[56-57]。本研究达乌尔黄鼠和亚洲小车蝗的生态位与绵羊的生态位重叠程度较大,说明它们的食谱相近,对食物资源存在激烈的竞争。造成食草动物生态位重叠的原因有很多,其中动物本身对植物的选择性采食和食物资源的可利用性是主要因素[58]。本研究发现,3种食草动物共同喜食的牧草种类很多,如多根葱、双齿葱和冷蒿等。此外,种群密度也是影响生态位重叠的一个重要因素[59],当群落中一个种群的密度增加,就会加大与其他种群生态位的重叠程度,同时也会影响其他与之生态位重叠的种群数量,它们之间是负相关关系[35]。本研究发现,在围栏内,鼠种的密度最大,在中度放牧区鼠种的密度最低,随着放牧强度的增加,鼠种的密度又增大,说明适度放牧会降低群落中鼠的密度,过度放牧又会加大群落中鼠的密度,而蝗虫密度随牧压的变化与鼠的密度变化相反,说明放牧会引发鼠、虫密度的消长[6],同时也说明过度放牧是引发鼠、虫害发生的直接原因。因此,控制鼠、虫的种群密度,防止鼠、虫害的发生对保证放牧家畜的食物安全也具有十分重要的现实意义。
[1]Chen Z Z,Wang S P.Typical Grassland Ecosystem in China.Beijing:Science Publishing House,2000:125-169.
[2]Xu Z X,Li Y Q.Influence of grassland degeneration on soil erosion.Journal of Arid Land Resources and Environment,2003,17(1):65-68.
[3]Li B.The rangeland degradation in North China and its preventive strategy.Scientia Agricultura Sinica,1997,30(6):1-9.
[4]Sun H Q,Zhou H,Wang P.Progress on grassland degenerated succession.Grassland of China,1999,(l):51-56.
[5]Wang S P,Wang Y F,Chen Z Z.Grazing System Management.Beijing:Science Publishing House,2003:150-290.
[6]Wan X R,Liu W,Wang G H,Zhong W Q.Food consumption and feeding characters of Phodopus roborovskii on Hunshandake sandy land of Inner Mongolia.Chinese Journal of Ecology,2007,26(2):223-227.
[7]Li X L,Zhang J,Zhang G S.Studies on the relationship between grazing succession of grassland and harmful rats density in alpine region.Pratacultural Science,1996,13(1):44-50.
[8]Huang C D.Effect of plateau xokoron grassland vegetatiln in cold and high meadow.Pratacultural Science,2004,21(10):58-62.
[9]Jiang X L.Relationship of population quantities of plateau Pika with vegetation homogeneity.Acta Pratacultural Sinica,1998,7(1):60-64.
[10]Kang L.Grasshopper-plant interactions under different grazing intensities in Inner Mongolia.Acta Ecologica Sinica,1995,15(1):1-11.
[11]He D H,Zheng Z M.Trophic niche and interspecific food competitive model of grasshoppers in desert grassland.Chinese Journal of Applied Ecology,1997,8(6):605-611.
[12]Wang S P,Li Y H,Wang Y F,Chen Z Z.Influence of different stocking rates on plant diversity of Artemisia frigida Community in Inner Mongolia Steppe.Acta Botanica Sinica,2001,43(l):89-96.
[13]Li Y H.The divergence and convergence of Aneurolepidium chinense steppe and Stipa grandis steppe under the grazing influence in Xilin river valley,Inner Mongolia.Chinese Journal of Plant Ecology,1988,12(3):189-196.
[14]Wang S P.Relationships between body gains and stocking rates of grazing sheep on typical Inner Mongolian grassland.Acta Prataculturae Sinica,2000,9(2):10-16.
[15]Kang L,Chen Y L.Dynamics of grasshopper communities under different grazing intensities in Inner Mongolian steppes.Entomologia Sinica,1995,2(3):265-281.
[16]Qiu X H,Li H C.Effect of livestock exclusion on abundance of grasshoppers on Aneurolepidium chinense and Stipa grandis steppes.Chinese Journal of Applied Ecology,1997,8(4):403-406.
[17]Liu W,Zhou L,Wang X.Responses of plant and rodents to different grazing intensity.Acta Ecologica Sinica,1999,19(3):376-382.
[18]Jiang T A,Wang S Q,Xue Z D.Correlation between vegetation coverage and Zoker population quantity.Bulletin of Soil and Water Conservation,2005,25(5):24-27.
[19]Fu H P,Wu X D,Yang Z L.Niche characteristics of rodents by diverse disturbance in Alashan Desert,Inner Mongolia.Acta Ecologica Sinica,2005,25(10):2637-2643.
[20]Kang L,Chen Y L.Trophic niche of grasshoppers within steppe ecosystem in Inner Mongolia.Acta Entomologica Sinica,1994,37(2):178-189.
[21]Wang G M,Zhou Q Q,Zhong W Q.Trophic niches of four species of common small mammals in Inner Mongolia grassland and their relationships.Acta Ecologica Sinica,1996,16(1):71-76.
[22]Mysterud A.Diet overlap among ruminants in Fennoscandia.Oecologia,2000,124(1):130-137.
[23]Fu H P,Ma C M,Ai D,Huang Y,Wu X D.The population niche of principal rodents in the Alashan desert region of Inner Mongolia.Journal of Inner Mongolia Agricultural University:Natural Science Edition,2003,24(4):22-25.
[24]Liu B W,Jiang Z G.Dietary overlap between przewalski's gazelle and domestic sheep in the Qinghai lake region and implications for rangeland management.Journal of Wildlife Management,2004,68(2):241-246.
[25]Melladoa M,Olverab A,Querob A,Mendoza G.Dietary overlap between prairie dog(Cynomys mexicanus)and beef cattle in a desert rangeland of northern Mexico.Journal of Arid Environments,2005,62(3):449-458.
[26]Steen H,Atle M,Gunnar A.Sheep grazing and rodent populations:evidence of negative interactions from a landscape scale experiment.Oecologia,2005,143(3):357-364.
[27]Dove H.Using the n-alkanes of plant cuticular wax to estimate the species composition of herbage mixtures.Australian Journal of Agricultural Research,1992,43(8):1711-1724.
[28]Dove H,Mayes R W.The use of plant wax alkanes as marker substances in studies of the nutrition of herbivores:a review.Australian Journal of Agricultural Research,1991,42(6):913-952.
[29]Valiente O L,Delgado P,de Vega A,Guada J A.Validation of the n-alkane technique to estimate intake,digestibility,and diet composition in sheep consuming mixed grain:roughage diets.Australian Journal of Agricultural Research,2003,54(7):693-702.
[30]Kuzyk G W,Hudson R J.Using n-alkane markers to estimate forage intake of mule deer.Canadian Journal of Zoology,2006,84(11):1576-1583.
[31]Liu G H,Lin L J,Zhang Y J,Wang S P,Han J G,Ma X Z.Study on the accuracy and precision of n-alkanes as markers for estimating of diet composition and herbage intake of sheep.Scientia Agricultura Sinica,2006,39(7):1472-1479.
[32]Fuls E R.Ecosystem modification created by patch-overgrazing in semi-arid grassland.Journal of Arid Environments,1992,23(1):59-69.
[33]Pickup G,Chewings V H.A grazing gradient approach to land degradation assessment in arid areas from remotely-sensed data.International Journal of Remote Sensing,1994,15(3):597-617.
[34]Dove H,Moore A D.Using a least-squares optimization procedure to estimate botanical composition based on the alkanes of plant cuticular wax.Australian Journal of Agricultural Research,1995,46(8):1535-1544.
[35]Van Dyne G M,Heady H F.Botanical composition of sheep and cattle diets on a mature annual range.Hilgardia,1965,36(13):465-492.
[36]Wang S P.The dietary composition of fine wool sheep under different stocking rates and relationship between dietary diversity and plant diversity in Inner Mongolia steppe.Acta Ecologica Sinica,2001,21(2):237-243.
[37]Zhang J T.Quantitative Ecology.Beijing:Science Publishing House,2004:77-97.
[38]Wang G J,Wang S P,Hao Y B,Cai X C.Effect of grazing on the plant functional group diversity and community biomass and their relationship along a precipitation gradient in Inner Mongolia Steppe.Acta Ecologica Sinica,2005,25(7):1649-1656.
[39]Jiang C,Zhang Y J,Luo H L,Chang S J.Optimum combination of plant wax compositions as markers in the composition estimation of dietary plant species of desert steppe in Inner Mongolia.Acta Agrestia Sinica,2012,20(3):576-582.
[40]Newman J A,Cribari-neto F,Jensen M J.The sensitivity of n-alkane analysis to measurement error:implications for use in the study of diet composition.The Journal of Agricultural Science,1998,131(4):465-476.
[41]Martins H,Elston D A,Mayes R W,Milne J A.Assessment of the use of n-alkanes as markers to describe the complex diets of herbivores.The Journal of Agricultural Science,2002,138(4):425-434.
[42]Wang S P.The dietary composition of fine wool sheep and plant diversity in Inner Mongolia steppe.Acta Ecologica Sinica,2000,20(6):951-957.
[43]Boo R M,Lindstrom L I,Elia O R,Mayor M D.Botanical composition and seasonal trends of cattle diets in central Argentina.Journal of Range Management,1993,46(6):479-482.
[44]Dummont B,Petit M,Dhour P.Choice of sheep and cattle between vegetative and reproductive cocksfoot patches.Applied Animal Behaviour Science,1995,43(1):1-15.
[45]Wang S P,Li Y H.Behavior ecology of grazing sheep V.Interrelation between ingestion behavior and sward characteristics.Acta Prataculturae Sinica,1997,6(4):31-38.
[46]Dong Q M,Zhao X Q,Ma Y S,Dai Y,Wang Q J,Shi J J,Wang L Y.Effects of grazing intensity on mixed-sown community of Elymus natans and Puccinellia tenuflora and Ochotona curzoniae in Yangtze and Yellow River headwater region.Acta Agriculturae Boreali-Occidentalis Sinica,2006,15(2):28-33.
[47]Zhao T B,Liang W,Qin F C,Liu Y H,Wang L M,Li J Z.A brief note on ecological study on Spermophilus dauricus.Journal of Inner Mongolia Normal University:Natural Science Edition,2000,29(2):125-129.
[48]Bai X W,Shi X M,Dong G R,Du G Y,Zhang Y Z.Feeding habits of Spermophilus dauricus Brandt in the north frontier administration areas of Hebei province.Chinese Journal of Vector Biology and Control,2010,12(4):382-383.
[49]Cao Y F,Lin G H,Lu X F,Su J P.Food habits of Ochotona koslowi.Chinese Journal of Zoology,2009,44(1):58-62.
[50]Yang W K,Liu W,Huang Y,Qiao H H,Xu W X,Xia C J,Li J,Xu F,Blank D.Food habits of great gerbil(Rhombomys opimus)in south of Gurbantunggut Desert.Arid Land Geography,2011,34(6):912-918.
[51]Nelson M L,Gangwere S K.A key to grasshopper food plants based on anatomical features.The Michigan Botanist,1981,20(3):111-126.
[52]Wang S G,Ma X M.Food selection and food niche of locust Catantops pinguis from Hangzhou,Zhejiang.Plant Protection,2009,35(4):39-43.
[53]Norbury G L,Sanson G D.Problems with measuring diet selection of terrestrial,mammalian herbivores.Australian Journal of Ecology,1992,17(1):1-7.
[54]Evans E W.Absence of inter specific competition among Tallgrass Prairie grasshoppers during a drought.Ecology,1992,73(3):1038-1044.
[55]Zuo X A,Zhao X Y,Zhao H L,Guo X R,Li Y Q,Zhao Y P.Changes on species composition and function diversity in degraded process of grassland in Horqin Sand Land.Journal of Soil and Water Conservation,2006,20(1):181-185.
[56]William J,Zielinski N,Duncan P.Diets of Sympatric Populations of American Martens(Martes americana)and Fishers(Martes pennanti)in California.Journal of Mammalogy,2004,85(3):470-477.
[57]Baldi R,Pelliza-Sbriller A,Elston D,Albon S.High potential for competition between guanacos and sheep in Patagonia.Journal of Wildlife Management,2004,68(4):924-938.
[58]Wissinger S A.Niche overlap and the potential for competition and intraguild predation between size-structured populations.Ecology,1992,73(4):1431-1444.
[59]Cheng Z Q,Zhang K B,Liu J,Chang J,Wang X,Wang L L,Sun P F.The study on vegetable niche of natural grassland of desertification grassland region in Yanchi County,Ningxia.Research of Soil and Water Conservation,2011,18(3):36-40,47-47.
参考文献:
[1]陈佐忠,汪诗平.中国典型草原生态系统.北京:科学出版社,2000:125-169.
[2]许志信,李永强.草地退化对水土流失的影响.干旱区资源与环境,2003,17(1):65-68.
[3]李博.中国北方草地退化及其防治对策.中国农业科学,1997,30(6):1-9.
[4]孙海群,周禾,王培.草地退化演替研究进展.中国草地,1999,(l):51-56.
[5]汪诗平,王艳芬,陈佐忠.放牧生态系统管理.北京:科学出版社,2003:150-290.
[6]宛新荣,刘伟,王广和,钟文勤.浑善达克沙地小毛足鼠的食量与食性动态.生态学杂志,2007,26(2):223-227.
[7]李希来,张静,张国胜.高寒地区草地放牧演替与害鼠密度关系的研究.草业科学,1996,13(1):44-50.
[8]黄成定.高原鼢鼠危害对高寒草甸植被的影响.草业科学,2004,21(10):58-62.
[9]江小蕾.植被均匀度与高原鼠兔种群数量相关性研究.草业学报,1998,7(1):60-64.
[10]康乐.放牧干扰下的蝗虫-植物相互作用关系.生态学报,1995,15(1):1-11.
[11]贺达汉,郑哲民.荒漠草原蝗虫营养生态位及种间食物竞争模型的研究.应用生态学报,1997,8(6):605-611.
[12]汪诗平,李永宏,王艳芬,陈佐忠.不同放牧率对内蒙古冷蒿草原植物多样性的影响.植物学报,2001,43(l):89-96.
[13]李永宏.内蒙古锡林河流域羊草草原和大针茅草原在放牧影响下的分异和趋同.植物生态学与地植物学学报,1988,12(3):189-196.
[14]汪诗平.内蒙古典型草原放牧绵羊体增重与放牧率之间的关系.草业学报,2000,9(2):10-16.
[16]邱星辉,李鸿昌.围栏禁牧对羊草草原和大针茅草原蝗虫丰富度的影响.应用生态学报,1997,8(4):403-406.
[17]刘伟,周立,王溪.不同放牧强度对植物及啮齿动物作用的研究.生态学报,1999,19(3):376-382.
[18]江廷安,王胜琪,薛志德.陕北黄土高原退耕还林地甘肃鼢鼠数量与植被覆盖度的相关性研究.水土保持通报,2005,25(5):24-27.
[19]付和平,武晓东,杨泽龙.不同干扰条件下荒漠啮齿动物生态位特征.生态学报,2005,25(10):2637-2643.
[20]康乐,陈永林.草原蝗虫营养生态位的研究.昆虫学报,1994,37(2):178-189.
[21]王桂明,周庆强,钟文勤.内蒙古典型草原4种常见小哺乳动物的营养生态位及相互关系.生态学报,1996,16(1):71-76.
[23]付和平,马春梅,艾东,黄英,武晓东.内蒙古阿拉善荒漠主要啮齿类种群生态位.内蒙古农业大学学报:自然科学版,2003,24(4):22-25.
[31]刘贵河,林立军,张英俊,汪诗平,韩建国,马秀枝.饱和链烷技术测定绵羊食性食量精确性研究.中国农业科学,2006,39(7):1472-1479.
[36]汪诗平.不同放牧率下绵羊的食性及食物多样性与草地植物多样性间的关系.生态学报,2001,21(2):237-243.
[37]张金屯.数量生态学.北京:科学出版社,2004:77-97.
[38]王国杰,汪诗平,郝彦宾,蔡学彩.水分梯度上放牧对内蒙古主要草原群落功能群多样性与生产力关系的影响.生态学报,2005,25(7):1649-1656.
[39]姜超,张英俊,罗海玲,常书娟.内蒙古荒漠草原区分采食成分的最佳蜡层指示剂组合.草地学报,2012,20(3):576-582.
[42]汪诗平.不同放牧季节绵羊的食性及食物多样性与草地植物多样性间的关系.生态学报,2000,20(6):951-957.
[46]董全民,赵新全,马玉寿,代勇,王启基,施建军,王柳英.放牧强度对江河源区垂穗披碱草 (Elymus natans)/星星草 (Puccinellia tenuflora)混播草地群落和高原鼠兔(Ochotona curzoniae)的影响.西北农业学报,2006,15(2):28-33.
[47]赵天飙,梁炜,秦丰程,刘艳华,王林梅,李建中.草原黄鼠生态学研究概述.内蒙古师大学报:自然科学版,2000,29(2):125-129.
[48]白雪薇,史献明,董国润,杜国义,张月芝.河北省塞北管理区达乌尔黄鼠食性调查.中国媒介生物学及控制杂志,2010,12(4):382-383.
[49]曹伊凡,林恭华,卢学峰,苏建平.柯氏鼠兔的食性分析.动物学杂志,2009,44(1):58-62.
[50]杨维康,刘伟,黄怡,乔洪海,徐文轩,夏参军,林杰,徐峰,Blank D.古尔班通古特沙漠南缘大沙鼠的食性.干旱区地理,2011,34(6):912-918.
[52]王世贵,马小梅.浙江杭州地区红褐斑腿蝗的食性选择及其取食生态位.植物保护,2009,35(4):39-43.
[55]左小安,赵学勇,赵哈林,郭轶瑞,李玉强,赵玉萍.科尔沁沙地草地退化过程中的物种组成及功能多样性变化特征.水土保持学报,2006,20(1):181-185.
[59]程中秋,张克斌,刘建,常进,王晓,王黎黎,苏鹏飞.宁夏盐池荒漠草原区天然草地植物生态位研究.水土保持研究,2011,18(3):36-40,47-47.
