鲁东南杨树人工林土壤微生物生物量碳的时空动态
2013-07-29尉海东董彬
尉海东,董彬,
1. 临沂大学资源环境学院,山东 临沂276000;
2. 南京师范大学地理科学学院江苏省环境演变与生态建设重点试验室,江苏 南京210023
土壤微生物量是土壤有机碳中最活跃的一部分,是土壤养分转化过程中的一个重要的活性汇或源, 在土壤有机质和养分循环转化、陆地生态系统的长期稳定中发挥着重要的作用。土壤微生物生物量碳是指土壤中体积小于5×103μm活的微生物生物量碳,约占土壤总有机碳的1%~5%[1],是促使土壤中有机物和植物养分转化、循环的动力,是形成土壤有机质的重要碳源[2-3],因而是土壤营养库的重要组成部分,在全球碳循环和碳平衡中有着十分重要的作用。土壤微生物量碳作为土壤有机质中最活跃和最易变化的组分,具有极高的灵敏性,可以在土壤总碳变化之前反映出土壤有机质成分微小的变化[4]。
森林土壤是森林生态系统中最大的碳汇,因此了解森林土壤碳是研究陆地生态系统碳循环的重要前提[5],随着全球碳贸易、碳循环等问题受到越来越广泛地关注,森林生态系统碳吸收特征和森林土壤微生物生物量碳的研究已成为土壤碳循环研究领域的热点[6-7]。杨树Populus tremuloides是一种生长迅速、适应性广、抗性强的优良树种,是我国北方的主要用材树种,占人工林总面积的70%以上,鲁东南地区的种植面积已达1.1×105hm2[8],在我国森林生态系统碳吸收中起着重要作用。但目前我国有关杨树人工林土壤微生物的研究仅有零星报道[9-12],但对杨树人工林土壤微生物生物量碳的时空动态的系统研究还未见报道,而且目前有关鲁东南杨树人工林土壤微生物生物量的数据非常缺乏。鉴此,本研究以鲁东南祊河林场不同发育阶段的杨树人工林为研究对象,开展土壤微生物生物量碳时空动态的研究,揭示其在土壤碳循环中的作用,旨在为正确评价我国杨树人工林碳汇功能提供数据基础,为杨树人工林可持续经营管理科学依据。
1 材料和方法
1.1 试验地概况
试验地位于山东省临沂市祊河林场(118°04′E, 35°15′N),属于暖温带季风区大陆性气候,四季分明,光照充足,雨量充沛。年平均气温14.1 ℃,极端最高气温39.5 ℃,最低气温-11.1 ℃,全年无霜期200 d以上,年平均日照时数为2532 h,年均降雨量850 mm,降水量的75%集中于夏季6-8月份。土壤类型为褐土,土壤质地为中壤,微酸,pH值5.6~6.7,结构良好,透气透水性较强[8]。
试验地为杨树Populus tremula人工纯林,分别为1994年(中龄林middle-aged Populus tremuloides plantation MAP)和2003年营造的2年生种苗(成熟林mature Populus tremuloides plantation MP),林木平均胸径分别为32.4和48.5 cm、平均树高分别为19.1和22.6 m、平均栽植密度分别为820和680 株·hm-2。林下植被稀少,仅见少量矮小草本,有灰绿藜Chenopodium glaucum,紫花苜蓿Medicago sativa,黄香草木樨Melilotus officinalis,蒺藜Tribulus terrestris等[8]。
1.2 样品的采集
在两种林分中各设立5个20 m×20 m的标准样地。每个样地随机选取9个样方, 每个样方采取对角线法布5点, 用环刀分别在0-10、10-20、20-40、40-60 cm层取样, 相同层次样品混合均匀后装入无菌自封袋, 带回实验室, 去除石粒、根等杂物后过2 mm筛, 将每份土样分成2份,其中1份土样保存在4 ℃冰箱内,并在一周内完成土壤微生物生物量碳(Cmic)的测定。另1份土样在室温条件下风干,碾磨过0.25 mm筛,用于测定土壤有机碳。每个样方设3个重复。从2011年2月至2011年12月采集土壤样品, 每两个月中旬定期采集一次。
1.3 样品的测定
土壤基本理化性质指标(土壤全N、全P、全K、有效P、土壤pH、有机质)和土壤有机碳采用文献11中方法测定。土壤微生物生物量碳采用氯仿熏蒸浸提法测定[12]。准确称取相当于20.0 g烘干土质量的土壤6份,预培养1周后,将其中3份土样用无水乙醇氯仿熏蒸, 于25 ℃放置24 h, 然后取出,反复抽真空去除氯仿;另外3份不进行熏蒸处理,作为对照。用0.5 mol L-1K2SO4(土液比为1:2.5)浸提土样,浸提液中有机碳采用重铬酸钾氧化法测定,土壤微生物生物量碳(mg∙kg-1)由下式求得:
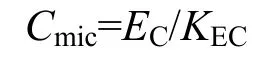
式中:EC为熏蒸与未熏蒸土样浸提液中有机碳的差值;KEC为校正系数,取值0.38。
1.4 数据分析
利用SPSS 17.0软件包进行平均值和标准差分析,并对不同林龄杨树林微生物生物量碳、土壤有机碳和土壤微生物熵进行单因素方差分析(one-w ay ANOVA),显著性水平均为p=0.05。采用OriginPro 8.0软件分析土壤微生物生物量碳的时空变化和土壤微生物生物量碳与土壤有机碳的关系。
2 结果与分析
2.1 杨树人工林土壤微生物生物量碳的季节动态
中龄杨树人工林和成熟杨树人工林土壤微生物生物量碳季节变化基本呈双峰趋势(图1),即生长季开始前下降、生长季结束时上升。两种林龄的土壤微生物生物量的变化范围分别为120.69~323.92 mg·kg-1和183.69~418.21 mg·kg-1,其峰值均出现在4月和10月,以4月最高,MAP和MP的分别达到323.92和418.21 mg.kg-1,其中0-10 cm土层的分别达到633.51和752.03 mg·kg-1;最低值均出现在12月,MAP和MP的分别为120.69和183.69 mg·kg-1,其中0-10 cm土层的分别为196.92和267.31 mg·kg-1。两种林龄中,0-20 cm土层季节变化均较明显,随着土层加深,土壤微生物生物量碳季节变化趋向平缓(图1)。
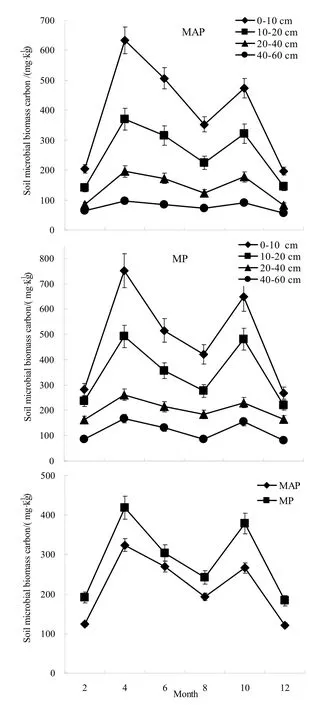
图1 杨树人工林土壤微生物生物量碳的季节动态 Fig.1 Seasonal dynamics of soil microbial biomass carbon for Populus tremula plantations
2.2 杨树人工林土壤微生物生物量碳的空间动态
土层、林龄和取样时间对杨树人工林土壤微生物生物量碳有显著影响(表1)。成熟杨树人工林土壤微生物生物量碳显著高于中龄杨树人工林的(图2),两种林龄的土壤微生物生物量碳均随土层深度的增加而显著降低(图2),土壤微生物生物量碳主要集中在0-40 cm土层,约占总土壤微生物生物量碳的89.77%~91.00%。中龄杨树人工林和成熟杨树人工林0-10 cm土层的土壤微生物生物量碳分别约为10-20 cm土层的1.56和1.38倍,为20-40 cm土层的2.83和2.37倍,为40-60 cm土层的5.04和4.09倍。土层和林龄对杨树人工林土壤有机碳亦有显著影响(表1、图2),与土壤微生物生物量碳的变化相似,两种林龄的土壤有机碳碳均随土层深度的增加而降低(图2)。成熟杨树人工林土壤微生物生物量碳和土壤有机碳显著高于中龄杨树人工林的(图2)。

表1 土壤微生物生物量碳、土壤有机碳和微生物熵的方差分析表 Table 1 The result of ANOVA for soil microbial biomass carbon, soil organic carbon and Cmic:Corg
2.3 杨树人工林土壤微生物生物量碳与土壤有机碳的关系
土壤微生物生物量碳与土壤有机碳的比值称为微生物熵(Cmic:Corg),是衡量土壤有机碳积累或损失的一个重要指标[3,13-14],能反映土壤中活性有机碳所占的比例,可作为土壤碳动态和土壤质量研究的有效指标[15],该值越高表示土壤碳呈积累趋势[16-17]。本研究中,林龄、取样时间和土层对杨树人工林土壤微生物熵有显著影响(表1),随土层深度的增加,两种林龄杨树人工林土壤微生物熵升高(图3),本研究中两种林龄的杨树人工林该值分别为2.22~3.18和2.62~3.85,土壤碳素均处于积累状态,表明深层土壤碳积累强度高于表层土壤,土壤有机碳逐渐由土壤表层向土壤深层转移,与易志刚等对鼎湖山保护区森林[17]和魏媛等对贵州喀斯特森林[18]研究结果一致。图3表明,中龄林已开始积累土壤有机碳,呈现碳汇功能,如果能准确判断土壤有机碳开始积累的时期,可为选择合适的轮伐时间提供依据。与中龄杨树人工林相比,成熟杨树人工林土壤微生物熵明显提高(图3),表明成熟杨树人工林土壤碳积累强度更高,可增加土壤碳储量,提高土壤肥力,改善土壤环境。杨树人工林土壤微生物生物量碳与土壤有机碳含量呈极显著线性正相关关系(r2=0.946, p﹤0.01)(图4)。可能主要是因为土壤有机质丰富,土壤微生物数量多,微生物对土壤有机物分解改造作用也强烈[3]。
3 讨论
3.1 杨树人工林土壤微生物生物量碳的季节动态
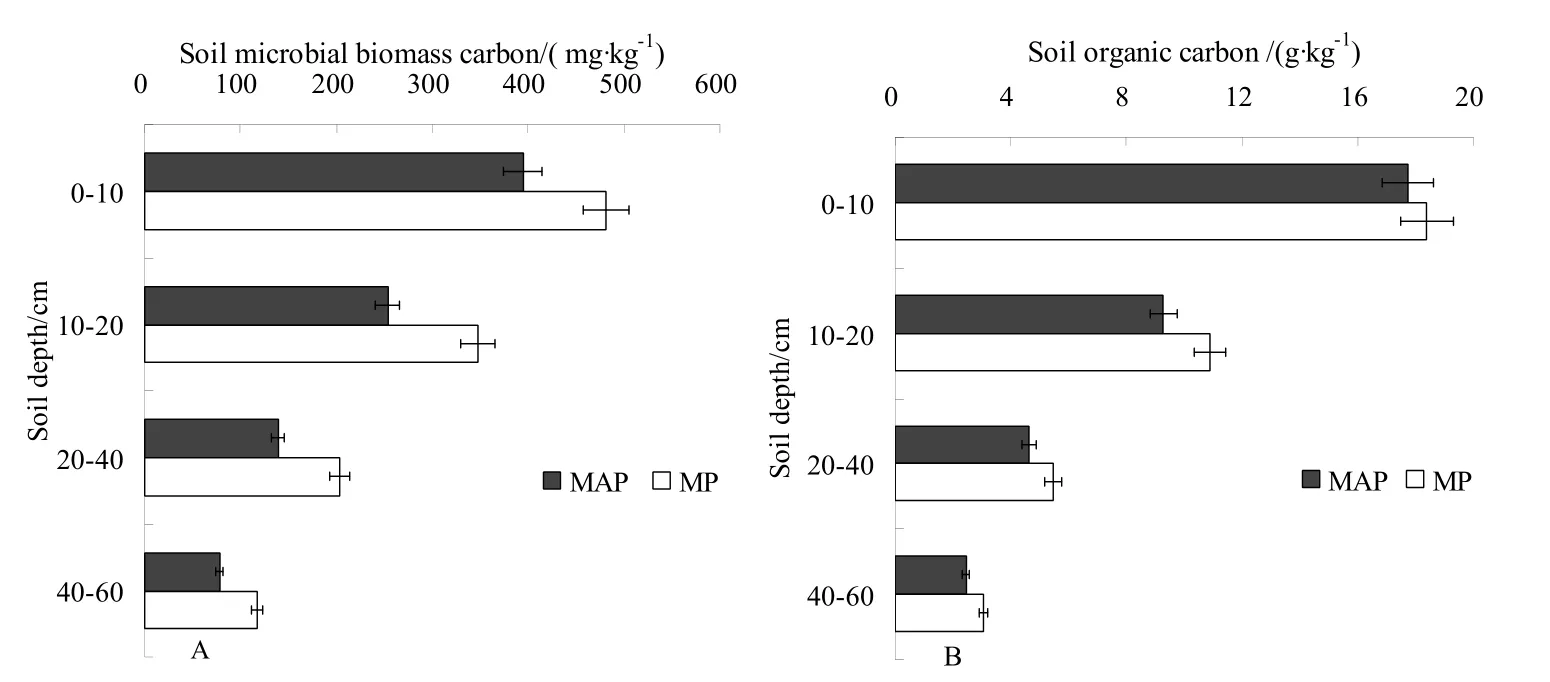
图2 土壤微生物生物量碳和土壤有机碳的垂直变化 Fig. 2 Vertical changes in soil microbial biomass carbon and soil organic carbon for two forest age
两种林龄的杨树人工林土壤微生物生物量碳有类似的季节变化趋势(图1)。土壤微生物生物量碳的季节变化受土壤温度[19]、土壤湿度[20]、植物生长节律和土壤中可利用碳和养分资源的限制[21]等因素的影响。土壤温度降低,微生物活性降低,土壤微生物生物量碳减少;温度升高,土壤微生物活性和数量增加,土壤微生物生物量碳含量增加。2月处于冬春交替时期,土壤温度开始回升,土壤水分和养分开始供应,土壤微生物开始活动,微生物生物量碳开始增加。进入4月,随着土壤温度的升高,微生物活性增强,林木根系和微生物共同固持养分可能形成“春坝”作用为土壤微生物的快速生长提供养分,从而使土壤微生物生物量碳迅速增加,出现全年最高峰,0-10 cm土层的分别达到633.51和752.03 mg·kg-1。6月,林木进入生长期需要从土壤中摄取更多的营养物质,在一定程度上形成与微生物之间的营养需求竞争,致使微生物的营养可获得量降低,微生物的繁殖和生长受到限制[22];土壤温度迅速升高,微生物活动增强,同时杨树根系亦进入快速生长状态,需要吸收大量养分,二者产生能源竞争,促使土壤进入快速矿化期[16],土壤微生物生物量碳开始降低。进入8月,高温高湿的环境条件有利于植物的快速生长,林木生长旺盛,对土壤N、P等养分的需求量大,郁闭度很高,林下光照不足,植被盖度较低,影响了土壤中微生物生物量;而且高温高湿的环境条件下,土壤微生物的活性增加,分解利用能源物质的速率加快,维持了较快的周转速率,由于受可利用资源的限制,土壤微生物生物量较小。进入10月,土壤温度明显降低,林木逐渐进入非生长期,能源消耗减少,秋季地表枯落物的增加,林内光照增多,大量养分开始由地上向地下转移,为微生物提供了大量能源,导致微生物生物量碳明显增加,出现了秋季高峰(图1)。12月份,土壤温度迅速下降,凋落物分解速率下降,能源供应不足,加之微生物活性迅速减弱,微生物生物量碳处于全年最低值。这一生长季开始前下降、生长季结束时上升(春秋高夏冬低)的季节变化趋势与邸雪颖等[19]对兴安落叶松林Larix gmelinii Rupr.土壤微生物生物量碳的研究、刘爽和王传宽[23]对黑龙江五种温带森林土壤微生物生物量碳和王国兵等[24]对北亚热带次生栎林和火炬松Pinus taeda L.人工林土壤微生物生物量碳的研究结果相似。而与刘满强等[25]的林地土壤微生物生物量碳夏季和冬季的数值高于春季和秋季的不一致。影响土壤微生物生物量碳主导因子不同是其产生不同的季节变化格局的主要原因。本研究中,土壤微生物生物量碳的季节变化可能受土壤温度、土壤湿度和土壤基质的综合影响。

图3 土壤微生物熵的垂直变化 Fig. 3 Vertical changes in soil Cmic:Corg for two forest age
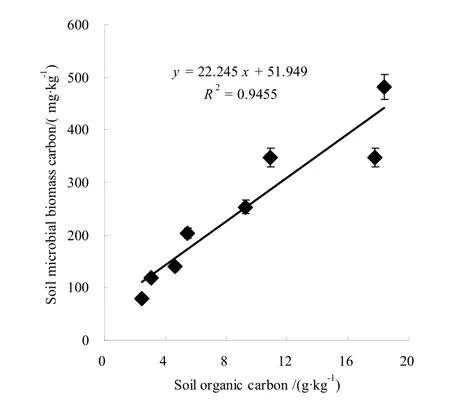
图4 杨树人工林土壤有机碳与土壤微生物生物量碳的关系 Fig.4 Relationship between soil organic carbon and soil microbial carbon for Populus tremula plantations
3.2 杨树人工林土壤微生物生物量碳的垂直动态
土壤微生物生物量碳含量为110~2240 kg/hm2表土,与土壤有机质呈正相关,与生态系统类型有关,草地最高,其次是林地,耕地最小。我国土壤微生物生物量碳为42~2046 kg/ hm2,为土壤有机碳的2%~4%[1],本研究中,土壤微生物生物量碳为土壤有机碳的2.22%~3.85%,在此范围之内。本研究结果0-10 cm土壤微生物生物量碳为394.56~481.33 mg·kg-1,低于刘爽等报道的硬阔叶林(1302 mg·kg-1)、杨桦林(1074 mg·kg-1)、红松林(748 mg·kg-1)、蒙古栎林(721 mg·kg-1)和落叶松林(534 mg·kg-1)[23];低于李灵等报道的福建三明格氏栲人工林[(1035±103) mg·kg-1]和杉木人工林的[(820±76) mg·kg-1][26];高于张秀玲等报道的鲁西北7年生速生杨人工林的(355.76 mg·kg-1)[27]。本研究中,杨树人工林土壤微生物生物量碳随土层的加深而降低(图2),这与严登华等对滦河流域杨树人工林的研究、邵玉琴等对皇甫川流域人工油松林Pinus tabulaeformis的研究[29]、李灵等对福建三明格氏栲人工林、杉木人工林的研究[26]、王春燕等对橡胶林的研究[6]、刘爽等对温带森林的研究[23]结果一致。而且本研究中,相同土壤深度下,成熟杨树人工林土壤微生物生物量碳显著高于中龄林的。土壤微生物多分布在土壤主要分布在0-20 cm,一方面因为在地表聚积大量枯枝落叶,有充分的营养源,水热和通气状况较好,利于微生物的生长和繁殖;另一方面也与上层土壤中相对密集的细根和较多土壤有机质、良好的土壤通气状况有关,随着土层的加深, 生境条件变差,从而影响微生物的生物量分布。
4 结论
1)鲁东南杨树人工林土壤微生物生物量碳存在明显的双峰季节变化趋势,即生长季开始前下降、生长季结束时上升。
2)鲁东南杨树人工林土壤微生物生物量碳存在明显垂直分布规律,即随土层深度的增加而显著降低。
3)鲁东南中龄杨树人工林和成熟杨树人工林土壤碳均处于积累状态,呈碳汇功能,且成熟杨树人工林积累碳能力比中龄林强。由于空间的限制,本研究只选择了两种林龄,未能准确判断杨树人工林土壤碳开始积累的关键时期,如果选择生境条件一致的4~5个林龄的杨树人工林进行深入研究,有可能为选择合适的轮伐时间提供依据。
[1] 张成霞,南志标. 土壤微生物生物量的研究进展[J]. 草业科学,2010, 27(6): 50-57.
[2] PANDEY C B, SINGH G B, SINGH S K, et al. Soil nitrogen and microbial biomass carbon dynamics in native forests and derived agricultural land uses in a humid tropical climate of India [J] Plant Soil,2010, 333(1-2):453-467.
[3] KUMAR, S CHAUDHURI, S MAITI, S K. Soil microbial biomass carbon in natural and degraded soil-A review [J] Environment and Ecology, 2011, 29 (3): 1689-1695.
[4] WANG X F, LI S Y, BAI K J, et al. Influence of doubled CO2on plant growth and soil microbial biomass C and N [J]. Acta Botanica Sinica, 1998, 40 (12):1169-1172.
[5] 吴艺雪,杨效东,余广彬. 两种热带雨林土壤微生物生物量碳季节动态及其影响因素[J]. 生态环境学报,2009, 18(2): 658-663.
[6] 王春燕,陈秋波,李光明,等. 不同树龄橡胶林土壤微生物生物量碳的垂直分布特性[J]. 林业资源管理,2012,2:59-64.
[7] 马和平,郭其强,刘合满,等. 西藏色季拉山土壤微生物量碳和易氧化态碳沿海拔梯度的变化[J]. 水土保持学报,2012, 26(4):163-171.
[8] 董彬. 土壤湿度对鲁东南杨树人工林地土壤呼吸的影响[J]. 西北农林科技大学学报: 自然科学版,2012,40(11):131-138.
[9] 安然,龚吉蕊,尤鑫,等. 不同龄级速生杨人工林土壤微生物数量与养分动态变化[J]. 植物生态学报, 2011, 35 (4): 389-401.
[10] 毛瑢,崔强,赵琼,等. 不同林龄杨树农田防护林土壤微生物生物量碳、氮和微生物活性[J]. 应用生态学报,2009, 20( 9): 2079-2084.
[11] 鲁如坤.土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 1999: 107-108, 147-148.
[12] 吴金水,林启美,黄巧云,等. 土壤微生物生物量测定方法及其应用[M].北京: 气象出版社, 2006: 54-87.
[13] DING M, YI W, LIAO L, et a1. Effect of afforestation on microbial biomass and activity in soils of tropical China[J]. Soil Biology and Biochemistry, l992, 24(9): 865-872.
[14] WOLTERS V, JOERGENSEN R G. Microbial carbon turnover in beech forest soils at different stages of acidification [J]. Soil Biology and Biochemistry, 1991, 23(9): 897-902.
[15] SPARLING G P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of change in soil organic matter [J]. Australia Journal of Soil Research, 1992, 30(2): 195-207.
[16] SINGH J S, REGHBANSHI A S, SINGH R S, et al. Microbial biomass acts as a source of plant nutrients in dry tropical forest and savanna [J]. Nature, 1989, 338: 499-500.
[17] 易志刚,蚁伟民,丁明懋,等. 鼎湖山自然保护区土壤有机碳、微生物生物量碳和土壤CO2浓度垂直分布[J]. 生态环境, 2006, 15(3): 611-615.
[18] 魏媛,张金池,俞元春,等. 贵州高原退化喀斯特森林恢复过程中土壤微生物生物量碳、微生物熵的变化[J]. 农业现代化研究, 2009, 30 (4): 487-490.
[19] 邸雪颖,耿莹莹,孙龙,等. 兴安落叶松林土壤微生物生物量季节动态及影响因素[J]. 东北林业大学学报, 2012,40(5):1-4.
[20] DEVI N B, YADAVA P S. Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-East India [J]. Applied Soil Ecology, 2006, 31(3): 220-227.
[21] RAPHAĚL C, SYLVIE B, OLIVIER C, et al. Impact of organic amendments on the dynamics o f soil microbial biomass and bacterial communities in cultivated land [J]. Applied Soil Ecology, 2007, 35(3): 511-522.
[22] BARBHUIYA A R, ARUNACHALAM A, PANDEY H N, et al. Dynamics of soil microbial biomass C, N and P in disturbed and undisturbed stands of a tropical wet-evergreen forest[J]. 2004, 40(3/4): 113-121.
[23] 刘爽,王传宽. 五种温带森林土壤微生物微生物量碳氮的时空格局[J]. 生态学报,2010, 30(12): 3135-3143.
[24] 王国兵,阮宏华,唐燕飞,等. 北亚热带次生栎林与火炬松人工林土壤微生物生物量碳的季节动态[J]. 应用生态学报, 2008, 19(1): 37-42.
[25] 刘满强,胡锋,何园球, 等. 退化红壤不同植被恢复下土壤微生物量季节动态及其指示意义[J]. 土壤学报, 2003, 40(6): 937-944.
[26] 李灵,张玉,王利宝,等. 不同林地土壤微生物生物量垂直分布及相关性分析[J]. 中南林业科技大学学报,2007,27(2): 52-57.
[27] 张秀玲, 李君剑, 石福臣. 速生杨人工林对土壤碳氮含量及微生物生物量的影响[J]. 生态与农村环境学报,2008, 24(2): 32-35.
[28] 严登华,王刚,金鑫,等. 滦河流域不同土地利用类型土壤微生物量C、TN、TP垂直分异规律及其影响因子研究[J]. 生态环境学报, 2010, 19(8): 1844-1849.
[29] 邵玉琴, 赵吉, 岳冰. 皇甫川流域人工油松林地土壤微生物的垂直分布[J]. 内蒙古大学学报:自然科学版, 2002, 33(5): 541-545.
