三裂叶蟛蜞菊入侵对土壤酶活性和理化性质的影响
2013-07-13柯展鸿邱佩霞胡东雄朱慧宋莉英
柯展鸿,邱佩霞,胡东雄,朱慧,宋莉英*
1. 广州大学生命科学学院,广东 广州 510006;2. 华南师范大学生命科学学院,广东 广州 510631;
3. 韩山师范学院生物系,广东 潮州 521041
生物入侵已经成为影响全球的重大环境和经济问题,不仅严重威胁着当地的生物多样性、生态系统的结构和功能[1],而且给社会带来严重的经济损失[2]。据不完全统计,目前我国有主要外来有害杂草107种,外来害虫32种,外来病原菌23种。这些外来生物的入侵给我国社会经济造成巨大危害,仅对农林业造成的直接经济损失每年就高达574亿元[3]。外来生物给入侵地造成严重破坏,随着时间的推移其对生态和经济发展的影响可能要更加严重。
作为生物入侵最直接的影响对象,土壤的质量状况(生物因素和理化因素)常常受到外来入侵种的影响[4]。许多研究表明,入侵植物能够通过化感作用、凋落物和根系结构等影响土壤生物多样性,同时通过对土壤酶活性、N循环、C循环和pH值的影响改变土壤的理化性质[5-9]。入侵植物在引起入侵地生物群落及土壤理化性质变化的过程中,形成对自身有利的土壤环境可能是其成功入侵的原因之一。
三裂叶蟛蜞菊Wedelia trilobata又称南美蟛蜞菊,为菊科Compositae多年生草本,原产南美洲及中美洲地区,20世纪70年代作为地被植物引入我国,很快逃逸为野生,目前已经成为华南地区最常见的杂草,所到之处,能够排挤本地植物,形成单优群落,严重威胁着当地的物种多样性[10],已被列为“世界最有害的100种外来入侵种”之一[11]。蟛蜞菊Wedelia chinensis分布于我国东、南部省区,与三裂叶蟛蜞菊同属菊科,二者具有相同的生活史特征,但蟛蜞菊生长较慢,对自然生态系统未造成危害。目前的研究认为,三裂叶蟛蜞菊之所以能够入侵成功主要归功于其快速的繁殖能力[12-13]、较高的光合速率[14-15]和较强的化感作用[16-18]。有关三裂叶蟛蜞菊对土壤酶活性和土壤理化性质的影响还未见报道。
本研究通过野外采样和盆栽试验,比较了三裂叶蟛蜞菊及其本地近缘种蟛蜞菊对土壤酶活性和土壤理化性质的影响,为评估三裂叶蟛蜞菊入侵对生态系统土壤微环境的影响提供试验依据。
1 材料和方法
1.1 植物材料
盆栽试验所用的三裂叶蟛蜞菊W. trilobata和蟛蜞菊W. chinensis均采自广州市华南植物园内的自然种群。分别剪取5 cm带芽茎段,进行扦插繁殖。经2~4周预培养后,选取生长健壮、长势一致的再生小苗移栽到塑料盆中(直径25 cm,高20 cm,容积8.5 L),基质采用营养土、河沙和塘泥(三者体积比为1:1:1)的混合物,每种植物各种植12盆,每盆移栽3株小苗,同时以不移栽小苗的12盆土样作为空白对照(CK)。
1.2 土样采集
1.2.1 野外土壤的采集
2012年4月从华南植物园内三裂叶蟛蜞菊和蟛蜞菊相邻生长的自然群落中进行土壤取样。由于二者生长在相邻的生境中,我们假定它们所生长的土壤初始条件基本一致。对于各物种生长的土壤,随机设置5个样方(1 m×1 m),分别采用5点取样法取样,先清除样地地面植物和凋落物,用土钻随机钻洞取0~15 cm层的土样250~500 g,取土样时避免阳光照射,去除石块、植物根系和土壤动物等,立即带回实验室过孔径为2 mm的筛。将过筛后的土样用于土壤含水量、土壤pH值、有机质、全氮、速效磷和速效钾含量的测定。
1.2.2 盆栽土样的采集
分别收集经受试植物正常生长3个月后的12盆土样,随机两两混合作为一个土样,分别置于聚乙烯袋中带回实验室,过孔径为2 mm的筛后用于土壤含水量、土壤pH值、有机质、全氮、速效磷、速效钾以及土壤酶活性的测定。
1.3 土样测定
参考刘凤枝和刘潇威[19]的方法,用sartorius PB-10酸碱度测定仪测定土样pH值,用四苯硼钠比浊法测定土壤速效钾含量,用纳氏比色法测定铵态氮含量,用酚二磺酸比色法测定硝态氮含量;参考刘守春和曾煜权[20]的方法,用钼锑抗比色法测定土壤速效磷含量;参考周建青和徐爱列[21]的方法,用重铬酸钾氧化-容量法测定土壤有机质含量;参考郭小颖[22]的方法,用开氏消煮滴定法测定土壤全氮含量;参考周礼恺和张志明[23]的方法测定土壤脲酶活性,其活性用单位土壤质量的NH3-N的毫克数表示(mg·g-1,以NH3-N计),用滴定法测定土壤过氧化氢酶活性,其活性用单位土壤质量的0.1 N高锰酸钾的mL数表示(mL·g-1,以0.1 N KMnO4计);参考沈桂琴[24]的方法测定磷酸酶活性,其活性用24 h单位土壤质量的苯酚的毫克数表示(μg·g-1·d-1, 以Phenol计);土壤含水量采用土壤烘干法测定,计算公式为:土壤含水量=[(原土壤湿质量-烘干土壤质量)/原土壤湿质量]×100%。
1.4 数据分析
数据统计采用SPSS 11.5 One Way ANOVA进行方差分析。
2 结果和分析
2.1 野外土壤的理化性质
由表1知,野外分别生长了三裂叶蟛蜞菊和蟛蜞菊的土壤理化性质有显著差异。与本地蟛蜞菊相比,三裂叶蟛蜞菊生长的土壤环境中pH值显著降低(P<0.01),而土壤有机质含量、全氮含量、速效磷和速效钾含量则显著提高(P<0.01),与蟛蜞菊相比分别提高了71.74%、78.89%、188.57%和46.36%,说明三裂叶蟛蜞菊入侵会导致土壤微环境的改变,提高土壤中可利用的营养物质含量。
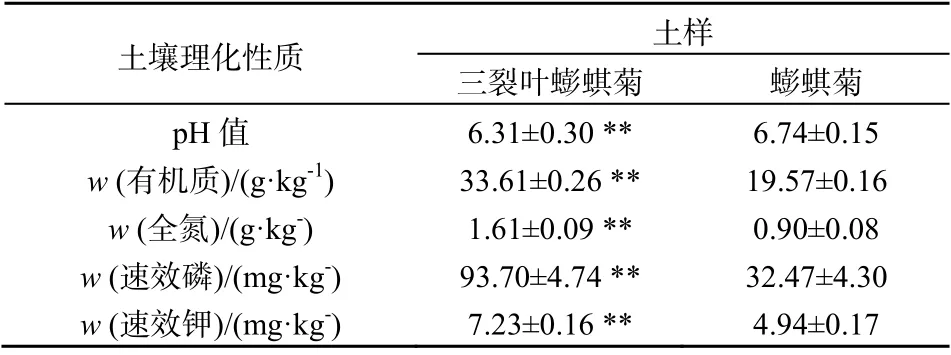
表1 三裂叶蟛蜞菊和蟛蜞菊群落野外生长样地土壤理化性质的比较 Table 1 Comparison on soil physical-chemical properties of field growth plot of Wedelia trilobata and Wedelia chinensis
2.2 盆栽土壤的理化性质
由表2知,经三裂叶蟛蜞菊和蟛蜞菊生长3个月的盆栽土壤,与CK相比,pH值均有显著下降,其中三裂叶蟛蜞菊生长过的土壤酸性显著低于本地蟛蜞菊。三裂叶蟛蜞菊生长后,土壤的全氮和硝态氮含量分别下降了9.94%和33.21%,速效钾含量上升了55.27%,与CK相比差异显著,而蟛蜞菊生长对土壤全氮、硝态氮和速效钾含量的影响不大。此外,三裂叶蟛蜞菊和蟛蜞菊生长后的土壤有机质、铵态氮、速效磷含量与CK相比,无显著差异。
2.3 盆栽土壤的酶活性
三裂叶蟛蜞菊和蟛蜞菊的生长会导致土壤酶活性的改变。其中三裂叶蟛蜞菊和蟛蜞菊生长的土壤中脲酶活性与CK相比分别下降了76.14%和65.19%(图1:A);三裂叶蟛蜞菊生长的土样中磷酸酶活性与CK相比没有显著性差异,而蟛蜞菊生长的土壤中磷酸酶活性与CK相比提高了12.5%(图1:B);过氧化氢酶活性在三裂叶蟛蜞菊、蟛蜞菊和CK之间均没有显著性差异(图1:C)。
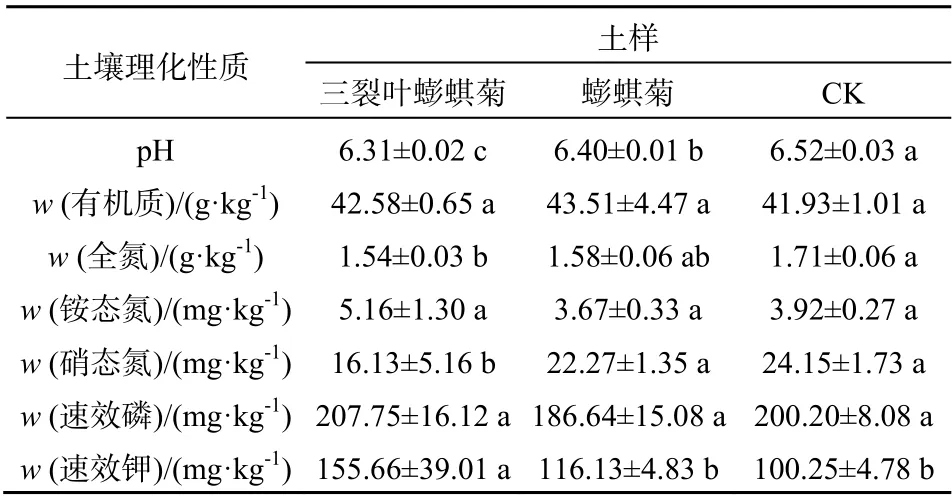
表2 三裂叶蟛蜞菊和蟛蜞菊盆栽土壤理化性质的比较 Table 2 Comparison on potting soil physical-chemical properties of Wedelia trilobata and Wedelia chinensis
3 讨论
植物入侵对土壤生态系统的生物多样性及土壤的性质能产生影响,但存在不一致的格局。入侵植物与土著植物在化感物质、凋落物的质与量、根系结构特征、生理生态特性的差异可能是形成格局多样性和影响机制复杂性的最主要原因[25]。周远谋[26]发现,马樱丹侵入木麻黄林后,木麻黄高径生长量分别提高7.4%和14.0%,混生林防风效能增加10.9%,土壤物理和化学性质都有不同程度的改善;朱慧和吴双桃[27]研究了杂草五爪金龙对入侵地植物群落和土壤肥力的影响,结果表明五爪金龙入侵导致入侵地植物群落丰富度和多样性显著降低,同时改善了土壤酶活性及养分水平,形成了对自身生长发育和种群扩增有利的生态微环境。
本研究通过对野外分别生长了三裂叶蟛蜞菊和蟛蜞菊的土壤的理化性质的比较,发现三裂叶蟛蜞菊入侵后土壤pH值显著降低,而土壤养分含量显著提高(表1)。有研究报道,不同群落下土壤肥力的差异可能与凋落物的质和量有关[28]。三裂叶蟛蜞菊入侵后能够快速形成单优群落,且群落植株密度大,生物量大,凋落物多,这可能是其提高土壤肥力的原因之一。此外,许多研究表明入侵植物可以通过改变土壤酶活性以及与土壤中具有一定生理功能的微生物类群之间的互作,增强土壤养分循环,提高植物可以直接吸收利用的土壤养分含量[5,6,25,28]。本研究对三裂叶蟛蜞菊盆栽土壤酶活性的测定结果显示土壤中脲酶的活性发生了显著变化,由此,我们推测,三裂叶蟛蜞菊入侵后也可能通过以上这种方式改变了土壤养分循环的过程,从而提高了土壤肥力。
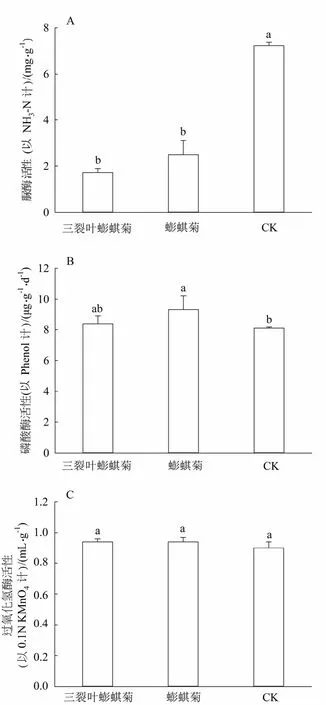
图1 三裂叶蟛蜞菊入侵对土壤酶活性的影响 Fig.1 Efects of Wedelia trilobata invasion on soil enzyme activities
由于入侵植物和入侵地土壤环境之间的互作存在复杂性,不同植物入侵、入侵时间长短、入侵种的生长节律以及入侵地的土壤环境等都会导致入侵的结果不同。盆栽试验中三裂叶蟛蜞菊生长的土壤环境中全氮含量和硝态氮含量与对照相比显著降低,而有机质含量、速效磷、铵态氮含量则没有发生显著性变化(表2),这与野外试验的结果有所不同。可能原因是三裂叶蟛蜞菊在前期种群建成的生长过程中需要消耗大量的土壤养分进行快速生长,导致土壤养分的降低,赵国晶和马云萍[29]对紫茎泽兰的研究也得到类似的结果;而野外自然种群中由于长期生物量的积累和凋落物的增加、以及它们对土壤酶活性及微生物结构的改变,提高了土壤的肥力。由此可见,入侵时期的不同可能导致三裂叶蟛蜞菊对土壤养分的影响不同。
研究表明,土壤脲酶直接参与土壤中含氮有机化合物的转化,其活性强度常用来表征土壤氮素供应强度,研究脲酶活性能够了解氮素转化能力和氮素有效化强度[30]。磷酸酶可以将土壤中的有机磷转化为自由离子,供植物吸收利用,对土壤中磷的代谢有重要的作用[31]。过氧化氢酶几乎存在于所有的生物体里,它能促进过氧化氢对各种化合物的氧化,有机质含量高的土壤, 过氧化氢酶的活性较强,其活性可以表征土壤总的生物学活性和肥力状况。本研究中,三裂叶蟛蜞菊生长的土壤环境中脲酶活性与CK相比显著降低(图1: A),这与表2中全氮含量和硝态氮含量降低的结果相对应。而磷酸酶活性和过氧化氢酶活性与CK相比则没有显著性差异(图1: B,C),该结果分别与表2中速效磷和有机质含量测定的结果相一致,说明它们受三裂叶蟛蜞菊生长的影响不大,栽培时间较短可能是原因之一,还有待更多的试验验证。由于盆栽试验所用土壤基质一致,所以栽种前土壤肥力及土壤酶活性差异不大,可以排除潜在的土壤养分和土壤酶活性异质性的影响,因此,我们认为群落间土壤养分和酶活性的差异是由群落物种的不同造成的,三裂叶蟛蜞菊生长的土壤环境中理化性质及土壤酶活性的改变是其入侵的结果。
4 结论
综上所述,三裂叶蟛蜞菊入侵到一个新的生境中,在前期的群落建成过程中能够通过快速消耗土壤中的养分含量,促进自身群落的构建;而在中后期群落形态结构成型后,能够通过降低土壤pH值、提高土壤养分含量,同时通过改变土壤酶活性等手段,改善土壤微环境,加速土壤养分循环,增强自身对养分的吸收,进而促进自身的生长、竞争和扩张。这可能是三裂叶蟛蜞菊入侵成功的原因之一。
[1] MACK R N, SIMBERLOFF D, LONSDALE W M, et al. Biotic invasions: causes, epidemiology, global consequences and control[J]. Ecological Applications, 2000, 10(3): 689-710.
[2] PIMENTEL D, MCNAIR S, JANECKA J, et al.. Economic and environmental threats of alien plant, animal, and microbe invasions[J]. Agriculture, Ecosystems and Environment, 2001, 84(1): 1-20.
[3] 刘用场. 生物入侵的危害及管理与控制对策[J]. 福建农业科技, 2005, 36(6): 69-70.
[4] 梁羽, 胡章立, 雷安平. 外来入侵种对土壤环境的影响[J]. 安徽农业科学, 2010, 38(3): 1652-1654.
[5] 李会娜, 刘万学, 戴莲, 等. 紫茎泽兰入侵对土壤微生物、酶活性及肥力的影响[J]. 中国农业科学, 2009, 42(11): 3964-3971.
[6] 牛红榜, 刘万学, 万方浩. 紫茎泽兰(Ageratina adenophora)入侵对土壤微生物群落和理化性质的影响[J]. 生态学报, 2007, 27(7): 3051-3060.
[7] 吴双桃. 薇甘菊对入侵地植物群落及土壤理化性质的影响[J]. 湖北农业科学, 2011, 50(18): 3711-3713.
[8] 高志亮, 过燕琴, 邹建文. 外来植物水花生和苏门白酒草入侵对土壤碳氮过程的影响[J]. 农业环境科学学报, 2011, 30(4): 797-805.
[9] 杜照奎, 李钧敏, 钟章成. 空心莲子草水浸提液的化感作用及化感物质分析[J]. 西南大学学报:自然科学版, 2012, 34(8): 53-60.
[10] 吴彦琼, 胡玉佳, 廖富林. 从引进到潜在入侵的植物—南美蟛蜞菊[J]. 广西植物, 2005, 25(5): 413-418.
[11] IUCN. 100 of the World’s Worst Invasive Alien Species[M]. Auckland: Invasive Species Specialist Group, 2001.
[12] 吴彦琼, 胡玉佳, 陈江宁. 外来植物南美蟛蜞菊的繁殖特性[J]. 中山大学学报:自然科学版, 2005, 44(6): 93-96.
[13] 陈笛, 刘家亮, 孟祥春,等. 南美蟛蜞菊花的生长发育[J]. 植物学通报, 2006, 23(1): 37-43.
[14] 刘金祥, 莫穗秋. 三裂叶蟛蜞菊光合生理特征对光合有效辐射增强的响应[J]. 草原与草坪, 2005, 25(109): 27-31.
[15] 孙蓓育, 孙兰兰, 宋莉英, 等. 入侵植物三裂叶蟛蜞菊叶片叶绿素荧光对温度的响应[J]. 华南师范大学学报:自然科学版, 2009, 44(4): 100-105.
[16] 聂呈荣, 曾任森, 黎华寿, 等. 三裂叶蟛蜞菊对菜心化感作用的生理机理[J]. 华南农业大学学报:自然科学版, 2003, 24(4): 106-107.
[17] 聂呈荣, 曾任森, 骆世明, 等. 三裂叶蟛蜞菊对水稻化感作用的初步研究[J]. 作物学报, 2002, 30(9): 942-946.
[18] 许华, 汪波, 魏宇昆, 等. 三裂叶蟛蜞菊对两种草坪植物的化感作用[J]. 广东农业科学, 2011, (14): 53-55.
[19] 刘凤枝, 刘潇威. 土壤和固体废弃物监测分析技术[M]. 北京: 化学工业出版社, 2007: 325-375.
[20] 刘寿春, 曾煜权. 土壤酸碱度及速效氮磷钾的测定[J]. 广西农业科学, 1964, 25(5): 36-42.
[21] 周建青, 徐爱列. 青海地区泥炭土壤中有机质含量测定方法的比较[J]. 安徽农业科学, 2010, 38(36): 20696-20697, 20700.
[22] 郭小颖. 开氏消煮-常量蒸馏-滴定法测定土壤中总氮量[J]. 环境研究与检测, 2011, (2): 33-35.
[23] 周礼恺, 张志明. 土壤酶活性的测定方法[J]. 土壤通报, 1980, 24(5): 37-49.
[24] 沈桂琴. 土壤中磷酸酶活性的测定方法[J]. 土壤肥料, 1987, 24(1): 40-42.
[25] 徐春荣, 肖文军. 植物入侵对土壤生物多样性及土壤理化性质的影响[J]. 安徽农业科学, 2010, 38(17): 9113-9115.
[26] 周远谋. 马樱丹入侵对木麻黄林生长和土壤肥力的影响[J]. 安徽农学通报, 2012, 18(3): 30-31.
[27] 朱慧, 吴双桃. 杂草五爪金龙对入侵地植物群落和土壤肥力的影响[J]. 生态与农村环境学报, 2012, 28(5): 505-510.
[28] 刘潮, 冯玉龙, 田耀华. 紫茎泽兰入侵对土壤酶活性和理化因子的影响[J]. 植物研究, 2007, 27(6): 729-735.
[29] 赵国晶, 马云萍. 云南省紫茎泽兰的分布与危害的调查研究[J]. 杂草学报, 1989, 3(2): 37-40.
[30] 陆建忠, 裘伟, 陈家宽, 等. 入侵种加拿大一枝黄花对土壤特性的影响[J]. 生物多样性, 2005, 13(4): 347-356.
[31] 雷瑞德, 党坤良, 张硕新, 等. 秦岭南坡中山地带华北落叶松人工林对土壤的影响[J]. 林业科学, 1997, 33(5): 463-470.
