烤烟种质资源形态学标记及SSR标记的多样性研究
2013-07-12陈建军赵伟才蔡一霞邓世媛
陈 杰,杨 静,陈建军,赵伟才,蔡一霞,邓世媛,王 维
(1 华南农业大学 农学院,广东 广州510642;2 贵州省黔东南州烟草公司,贵州 凯里556000;3 广东省烟草南雄科学研究所,广东 南雄512400)
烟草Nicotiana tabacum 属茄科Solanaceae 烟草属Nicotiana,是我国乃至全世界最重要的经济作物之一.烟草品种是烟叶生产的基础,通过改良品种提高烟叶质量,以满足卷烟工业的要求,是一条投资少、收益高的最佳途径[1].近年来,在烟草育种中过度使用主体亲本,尤其是我国烟草育种以NC89、G28、K326 等优质品种作为中心品种进行烟草育种,育种目标单一,使得育成品种遗传背景相似、品种单一化现象严重[2-3].育种中心品种的选择范围较狭窄,育成的品种往往具有比较相同的优缺点,对病害的抗感性也基本一致,一旦病害大面积发生,对烟叶生产会造成不利影响.大多数烟区品种单一化问题较为突出,急需新品种,但现在可供选择的品种较少,无法满足生产的要求[4].因此,烟叶生产迫切要求育种工作者更多、更快、更好地推出新品种.遗传多样性是生物多样性研究的核心,它反映出物种的遗传背景、育种潜力和利用价值,对烟草种质遗传多样性及不同烟草类型间的遗传关系进行研究,对充分发掘利用现有种质、合理选配亲本、拓宽育成品种遗传基础和保护优质种质具有十分的重要意义[5].近年来,利用DNA 分子标记技术对烟草种质资源遗传多样性开展了大量的研究[6-16],并由单一的分子标记技术发展到多种分子标记技术对同一供试材料进行亲缘关系的研究[17-20],均得出烟草种间遗传多样性较丰富、普通烟草栽培种内亲缘关系较近的结论.利用形态学标记和分子标记2 种标记结合的方法对种质材料进行多态性分析并不多见[21-22],在烟草种质遗传多样性的分析中鲜见相关报道.
烟草作为重要的经济作物,叶片是其收获器官,现蕾期的形态学性状作为衡量烟草产量和品质的一个重要参考依据,现蕾天数影响到着生叶片数,而叶片数直接控制着产量的高低.田间杂交育种对亲本的选择不仅要看其亲缘关系的远近更要看其在当地气候条件下的表型特征及抗逆性,并于现蕾期选择生长健壮、性状优良的烟株进行留种,待开花进行杂交.因此烟草现蕾期形态学性状的考察对杂交育种工作的顺利开展具有十分重要的意义.本文以烤烟现蕾期的形态指标作为形态学标记,结合SSR 分子标记技术对69份烤烟种质材料进行遗传多态性分析,试图为烤烟田间育种中优良品种、品系的选育,尤其是烤烟早花育种中对抗早花育种材料的直观选择提供依据,最大限度地挖掘利用现有种质资源.
1 材料与方法
1.1 材料
参试烤烟种质69 份,由广东省烟草南雄科学研究所提供.材料名称及来源见表1.
SSR 引物信息来自Bindler 等[23]公开发表的烟草SSR 连续图谱及其引物序列,在各条染色体上均匀选取2~3 对(共42 对)SSR 引物用于遗传多样性检测.序列由生工生物工程(上海)有限公司合成;Taq DNA Polymerase 和dNTPs 均购自新标科信生物科技(北京)有限公司.
1.2 方法
1.2.1 形态学性状调查 烟草成苗移栽,各个材料种植3 个小区,每小区10 株,株距为0.6 m,行距为1.2 m.记录移栽至现蕾天数,调查农艺性状包括着生叶片数、株高、最大叶长和最大叶宽,按《烟草种质资源描述规范和数据标准》[24]调查数据.
1.2.2 DNA 提取 按上海植物生理研究所的TPS抽提法稍加修改进行.具体过程如下:剪取新鲜叶片1 g 左右,放入2 mL 离心管中,加入液氮,迅速捣碎,加入800 μL 预热的TPS 抽提液[100 mmol/L Tris-Cl(pH 8.0),10 mmol/L EDTA(pH 8.0),1.0 mol/L KCl],75 ℃水浴30 min,12 000 r/min 离心15 min,取上清液,加入2/3 体积的苯酚/三氯甲烷/异戊醇(体积比25∶24∶1)溶液,摇动数分钟,12 000 r/min 离心10 min,取上清液,加入2 倍体积的无水乙醇,-20 ℃放置30 min,12 000 r/min 离心10 min,倒掉上清液,用φ(乙醇)为70%的溶液洗2 次,风干,溶于200 μL 0.5×TE 溶液中,-4 ℃保存备用.
1.2.3 SSR-PCR 反应条件 PCR 反应总体积为20 μL,反应体系中含1×PCR Buffer,dNTPs 0.2 mmol·L-1,1.5 U Taq 酶,0.2 μmol·L-1引物,20 ng 模板DNA.PCR扩增在Eppendorf mastercycler gradient 22331 hamburg上进行,先在94 ℃下预变性3 min;再进行35 个循环的变性(94 ℃50 s)、退火(55 ℃45 s)、延伸(72℃1 min)步骤;然后在72 ℃下继续延伸5 min;最后慢慢冷却至10 ℃保持.
1.2.4 电泳及检测 PCR 扩增产物加1/6 体积的上样缓冲液,取3 μL 利用质量分数为6%聚丙烯酰胺(PAGE)凝胶电泳分离,5 V/cm 恒压进行电泳,电泳后,拆下胶板,蒸馏水中漂洗2 次,显影,待条带出现,拍照记录.
1.2.5 表型数据分析 利用DPS 6.55(Data processing system)软件对形态学数据进行基本分析,计算广义遗传率,进行主成分分析.对农艺性状进行Q型聚类分析,采用效果较好的类平均法(Unweigted pair-group method using arithmetic averages,UPGMA)进行聚类,绘出聚类树形图.根据聚类结果,以类群结合的水平为纵坐标,结合次数为横坐标,将全部聚合过程画成一条阶梯式的折线图(聚合结合线),用于确定一条分类等级的划分界限.
1.2.6 基因型数据分析 扩增产物按同一迁移位置的条带在各材料中有(记为1)或无(记为0)进行统计.利用NTSYS-pc Version 2.10e 软件包计算遗传相似系数,并按照UPGMA 法进行聚类分析,构建聚类图[25].
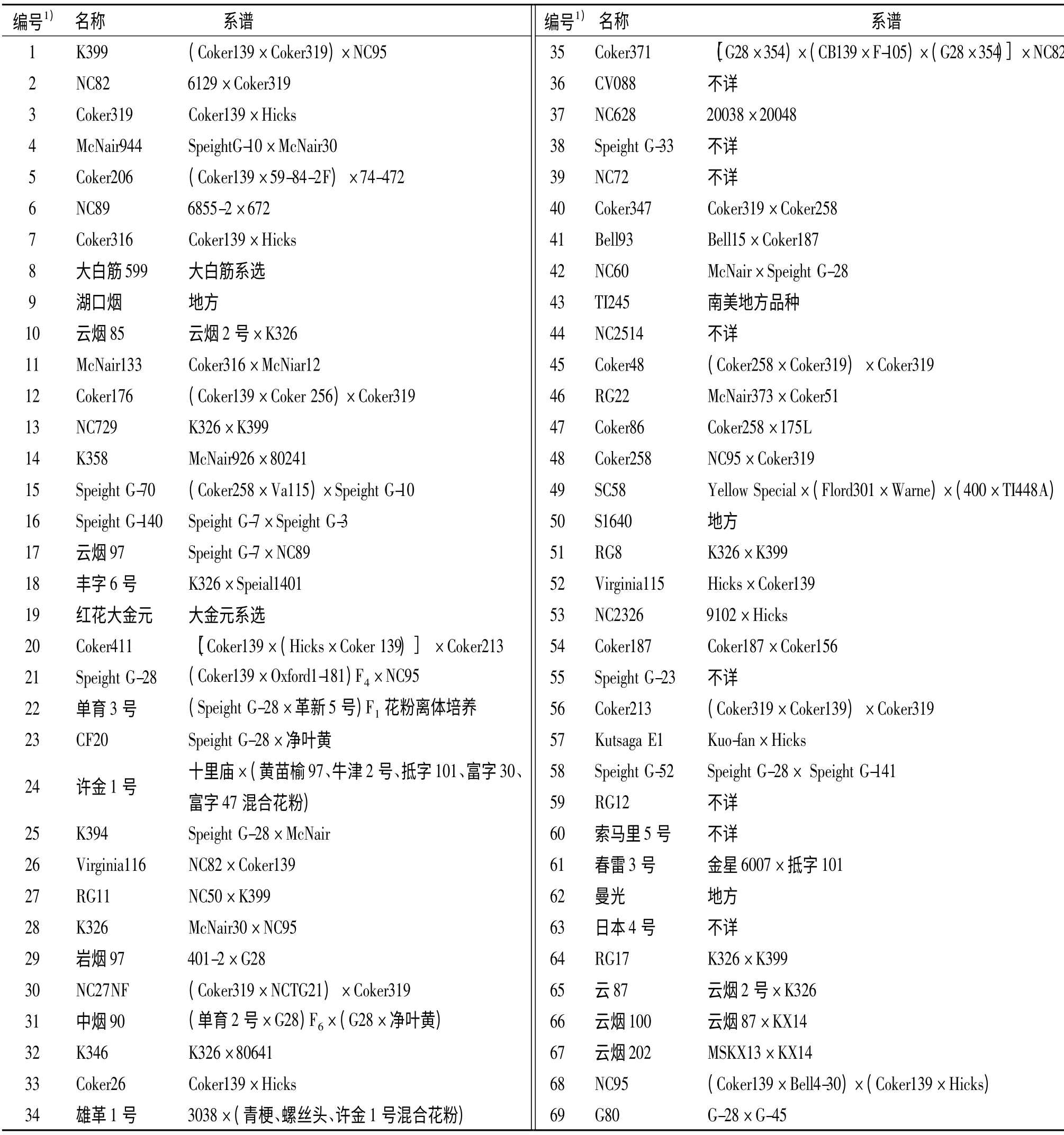
表1 69 份供试烤烟品种编号及来源Tab.1 The number and origin of 69 flue-cured tobacco varieties studied in the experiment
2 结果与分析
2.1 形态学性状分析
对烤烟农艺性状指标分析(表2)可知,参试品种的各个指标都具有广泛的变异,各个指标的变异系数都达到了13%以上,其中以现蕾天数的变异系数最小,为13.1%,变异范围为65.31~87.20 d.株高的变异系数最大,达到了36.1%,变异范围为60.22~115.30 cm.各个指标的广义遗传率较高,都达到了70% 以上,其中以现蕾天数最高,为87.59%,说明对各个指标早期进行选择效果较好.对现蕾期各个指标进行主成分分析,分别计算出各个指标的贡献率,其中以现蕾天数的贡献率最高,为75.48%,其对应的变异系数为各指标中最低,在各个指标中最为稳定,是反映品种特性、影响现蕾期的最主要指标.着生叶片数的贡献率在所有指标中居第2 位,为13.40%,是影响现蕾期的次要指标.
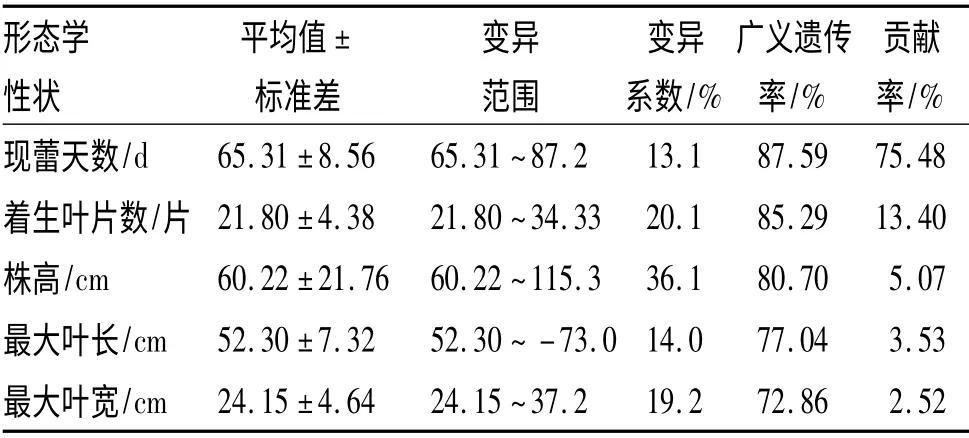
表2 烤烟种质形态性状分析Tab.2 The morphological characters analysis of flue-cured tobacco germplasm
2.2 SSR 标记的遗传多态性分析
选择遗传差异大的6 份烟草种质(编号分别为4、24、31、43、57 和62)对42 对SSR 引物进行筛选,初步筛选出15 对多态性好、重复性好的引物用于烟草种质的SSR 分析,结果有6 对引物在所有材料中能扩增出清晰且稳定的多态性带,其余9 对引物因扩增带模糊或多态性带较弱而无法统计.6 对SSR 引物共扩增出76 条稳定的条带,其中多态性条带66 条,平均每个引物扩增11 条多态性条带,引物的平均多态性比率为86.8%(表3).在所用的SSR 引物中,以引物PT20485 区分69 个品种的效果最为明显,每个品种都有其各自独特的指纹图谱(图1).即使是亲缘关系很近的品种,也可根据指纹将其分开,如1 号K399、28 号K326 和51 号RG8(RG8 由K326 和K399 杂 交 选 育 而 成).这 说 明SSR 标记在烤烟遗传资源中具有丰富的遗传多态性,可以很好地揭示供试材料间的遗传差异和亲缘关系.

表3 筛选出的6 对SSR 引物名称、序列及其扩增结果Tab.3 The six screened primer pairs applied to SSR analysis
2.3 烤烟种质资源的遗传相似性与差异性分析
由形态学性状数据估算出烤烟种质材料间的遗传距离,69 份烤烟种质材料间的遗传距离在0.32~7.83 之间,平均为2.80.其中,来自美国的烤烟品种Coker316(Coker139×Hicks 回交多次)与美国品种SpeightG-52(SpeightG-28×SpeightG-141)的遗传距离最小,为0.32,相似度最高;品种Coker316 与其他各个品种的遗传距离也较小,平均值最小,仅为1.99;南美收集的地方品种TI245 与品种Coker86(Coker258×175L)的遗传距离最远,为7.83,与其他各个品种的遗传距离的平均值较高,为4.99;中国烤烟品种春雷3 号(金星6007×抵字101)与其他各个品种的遗传距离的平均值最大,达到了5.20.
由SSR 标记数据估计烤烟种质材料间的遗传相似性系数,结果表明69 份材料的遗传相似系数在0.30~0.96 之间,平均为0.70.其中来自日本的烤烟品种日本4 号与K326 的遗传相似系数最低,为0.30;同是来自美国的烤烟品种Virginia115(Hicks×Coker139)与NC2326(9102×Hicks 与Hicks 回交3 次)之间的遗传相似系数最高,达到了0.96,2 个品种与其他品种的遗传相似系数也较大,平均值都较高;来自中国河南的地方品种曼光与其他各个品种的遗传相似系数的平均值最小,为0.54;同样来自中国的烤烟品种CV088(来源不详)与其他各个品种的遗传相似系数的平均值最大,为0.76.
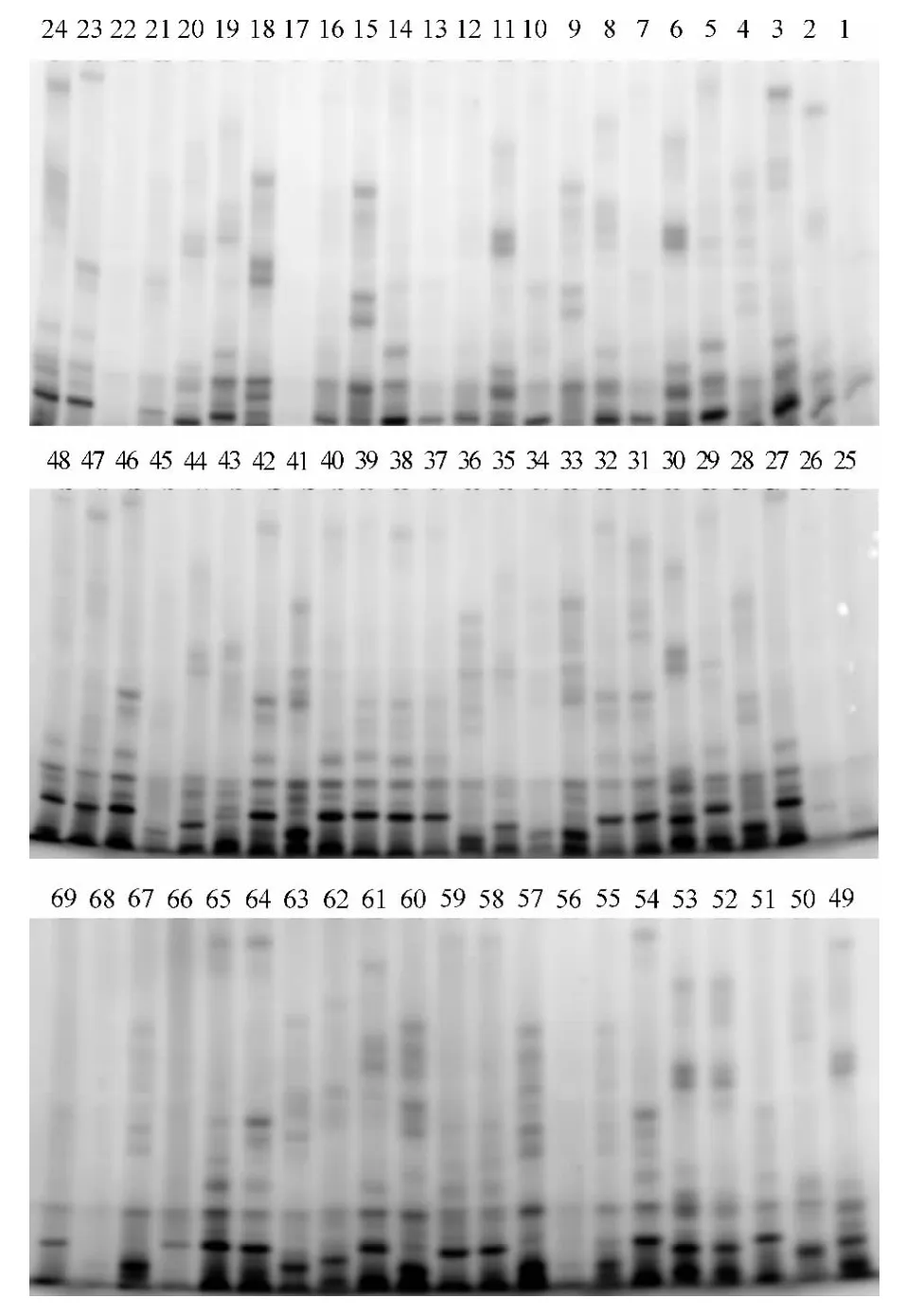
图1 引物PT20485 对69 份烤烟品种扩增产物的聚丙烯酰胺电泳图Fig.1 Polyacrylamide gel electrophoresis profile of SSR fragments from 69 flue-cured tobacco varieties amplified with the primer PT20485
2.4 烤烟种质资源的聚类分析
反映聚合过程的Q 型聚类结合线(Cluster combined-line)如图2 所示.从图2 可以看出,品种类群的聚合在纵轴上的分布是不均匀的,类群结合水平先后出现过几次飞跃,这些飞跃便将类群的结合区分为不同等级,从而形成系统分类的等级性.选择适当的飞跃,取其跳跃位置的中点作结合线L1和L2:
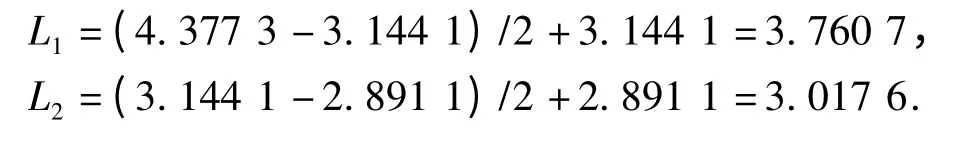
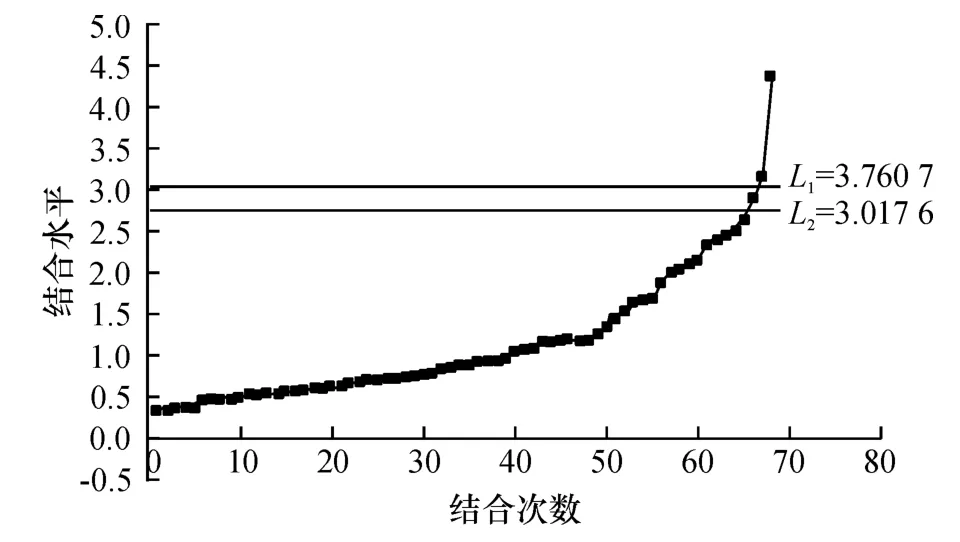
图2 Q 型聚类结合线Fig.2 Q type cluster joint line
将L1和L2分别标在聚类结果图(图3)上.从图3可以看出,L1将各个品种明细地划分为2 大类群:A大类包括1 号、16 号、26 号、41 号等52 个品种;B 大类包括3 号、12 号、31 号、47 号等17 个品种.A 类群的各项性状指标均比B 类群要小,A 类群各个品种的平均现蕾天数为61.62 d,着生叶片数20.07 片,植株高度49.93 cm,最大叶长48.99 cm,最大叶宽22.37 cm;B 类群各个品种的平均现蕾天数为76.60 d,着生叶片数27.10 片,植株高度91.73 cm,最大叶长62.41 cm,最大叶宽29.59 cm.B 大类各项农艺性状指标比A 大类分别高24.31%、35.02%、83.73%、27.38%和32.29%.
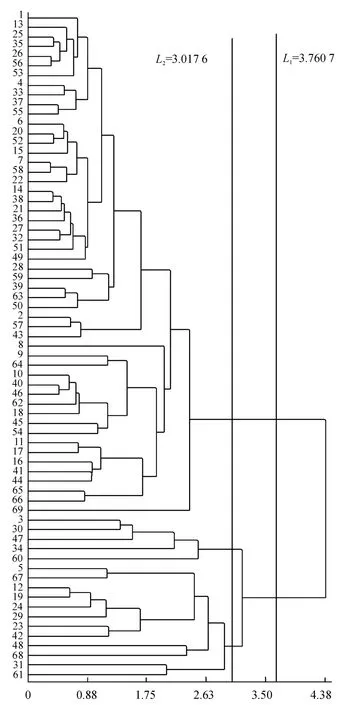
图3 基于形态学性状确定的遗传距离的品种聚类图Fig.3 Dendrogram of flue-cured tobacco cultivars based on the morphological characters
L2将69 个品种划分为3 类:第1 类为A 大类,包括52 个品种;第2、3 大类同属于B 大类,这2 类的划分主要依据为着生叶片数,第2 类包括3 号、30号、34 号、47 号、60 号5 个品种,各个品种的着生叶片数都达到了30 片以上;第3 类包括5 号、12 号、24号、42 号、67 号等在内的12 个品种,各个品种的着生叶片数都没有超过30 片,平均值为24.59 片.结合表2 分析的各个农艺性状指标的贡献率来看,现蕾天数贡献率最高,为品种类群划分的主要依据,着生叶片数的贡献率第2,为其划分类群的次要依据,根据着生叶片数的不同将B 类群又划分为2 大类群.
根据SSR 分子标记估算的各品种的Jaccard 相似系数(图4)可知,第1 等级按Jaccard 相似系数(JS1)=0.63 可将69 份材料分为2 大类:第1 大类包括1 号、3 号、10 号、38 号等66 个品种,这些品种大多具有NC82、NC89、G28、G80、K326、K346 等品质优良且作为育种核心种质资源的品种的血缘关系;第2 大类包括43 号、62 号、63 号3 个品种,这3 个品种分别为来自南美、中国、日本的地方烤烟品种,来源不详,与其他种资的遗传相似系数较小.
第2 等级按Jaccard 相似系数(JS2)=0.67 将所有种质资源划分为4 大类.第1 等级划分中的第1 大类被细分为2 大类:1)a 小类,大多为利用品质优良的品种通过各种育种手段改良的后代,如13 号品种NC729 为K326 与K399 的杂交后代,17 号品种云烟97 为Speight G-7×NC89 的后代;2)b 小类,大多是利用地方品种与品质优良品种杂交选育的后代,如31 号品种中烟90 是(单育2 号×G28)F6×(G28×净叶黄)的后代选育而成,49 号品种SC58 由Yellow Special×(Flord301×Warne)×(400×TI448A)选育而成.第1 级划分中的第2 大类被细分为2 大类,其中来自南美的地方品种TI245 单独划分为一类,来自中国和日本的地方品种划分为一类,这可能与烟草传入我国路线之一是由日本传入我国东北有关[26].
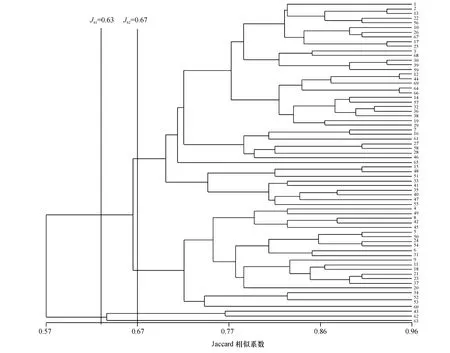
图4 基于SSR 分子标记确定的69 份烤烟品种遗传相似系数的品种聚类图Fig.4 Dendrogram of 69 flue-cured tobacco cultivars based on the SSR molecular markers
3 讨论与结论
3.1 烤烟SSR 标记的遗传多态性
SSR 引物在烟草上的研究应用相对较晚,2007年,Bindler 等[23]报道了烟草的微卫星标记遗传图谱,2011年Bindler 等[27]在前期工作的基础上筛选出2 317 个多态性和重复性好、条带清晰的SSR 标记,构建了包括2 317 个SSR 标记、2 363 个位点在内的24 个连锁群,遗传图谱总遗传长度为3 267 cM、标记间的平均距离小于1.5 cM.这是目前世界上包含标记位点最广、标记密度最大、平均图距最小的烟草分子连锁图谱.近年来SSR 分子标记技术在烟草遗传育种上的应用越来越深入、广泛[28-31].Cossirat[32]用160 个RAPD 引物对32 个烟草品种进行遗传多样性分析,其中只有9 个(占5.6%)引物能够扩增出29 条稳定、清晰的多态性条带.郭金平等[33]等利用150 个RAPD 随机引物对22 份烤烟种质进行遗传多样性分析,筛选出的7 个引物(占4.67%)共扩增出条带清晰、稳定的38 个谱带,平均每个引物扩增出5.4 条带,多态性条带比例为86.8%.聂琼等[30]从22 对SSR 引物中筛选到多态性、重复性、稳定性都较好的8 对引物,8 对引物共检测到72 条谱带,60 条遗传多态性带,多态比例83.33%.徐军等[31]利用Bindler 等[23]公布的286 对SSR 引物中筛选出的8 对反应稳定、扩增条带清晰、多态性强的引物对80 份普通烟草种质的指纹图谱进行分析,共扩增出85 个多态性位点,平均每对引物10.6 个.本研究通过对42 个SSR 引物的PCR 分析,有6 个引物(占14.3%)能够在69 个烤烟品种中共扩增出清晰、重复性好的条带76 条,平均每个引物扩增12.7 条,多态性比率为86.8%.由此表明,SSR 分子标记比RAPD 更适于烟草种质遗传多态性研究;同时,SSR 分子标记技术具有对DNA 模版质量要求不高、电泳结果清晰、重复性好等优点,使得SSR 分子标记技术容易掌握,对种质的多样性研究的结果准确度高,能很好的揭示其亲缘关系.
3.2 烤烟品种间的遗传多样性和亲缘关系
种质遗传多样性及遗传关系的研究对作物的杂交育种亲本的合理选择、利用具有十分重要的意义[5].本研究中69 份烤烟种质资源间的遗传距离变化范围在0.32~7.83 之间,平均值为2.80,遗传变化范围较大,总体遗传距离较近.遗传相似系数及聚类分析表明,遗传相似系数的范围较宽,品种间相似性较高,遗传相似系数在0.30~0.96 之间,平均为0.70.可见,69 份烤烟种质材料的遗传多样性较低,亲缘关系较近.这与前人[32,34-38]对烤烟种质资源多态性研究的结果基本一致.长期以来,烟草育种的主体亲本主要集中在从古老的Orinoco 衍生出来的Hicks、Coker139 及其衍生的K326、K346、NC82、NC89、G28 等少数优质种质[39],加之定向选择,使得育成品种遗传基础狭窄、品种单一化现象极为严重.目前,我国大面积种植的烤烟品种K326、云烟87(云烟2 号×K326)、云烟85(云烟2 号×K326)等品种亲缘关系十分相似.近年来,我国各个烟草产区,根据自身的气候生态条件及其对不同风格香型的烟草产品的要求,育成一批品种遗传性状稳定、品质优良、抗性较强的烤烟新品种,如粤烟97(MS207×粤烟96,粤烟96 为K326 优良变异系)、中烟100(9201×NC89多次回交)、豫烟6 号(MSK326×自育的农大202)、湘烟3 号(MSY2206-9×82-11-7,MSY2206-9 具有MSK326 的血缘,82-11-7 由K349 与云烟87 杂交而成)等品种,这些品种在选育过程中所用的亲本范围比较狭窄,育种目标基本一致,造成了某些基因的定向选择,导致许多非育种目标的多样化性状丢失.这可能是导致烤烟种质遗传多样性较低的主要原因.
3.3 形态标记和分子标记
形态标记是遗传标记的一种,是人们最早利用的遗传标记.由于其操作简单、直观、便于观察,长期以来,物种的分类及鉴定都是以形态标记为主要或初步的指标.但形态学标记的缺点是遗传表达不太稳定,易受环境及基因影响.分子标记是继形态标记、细胞标记和生化标记之后发展起来的一种最为可靠的遗传标记技术.分子标记技术不受环境、季节的影响,与形态标记相比,更适用于植物分类和遗传多样性研究.本试验利用形态学标记,根据现蕾期农艺性状指标将69 个品种划分为早花类型品种和晚花类型品种.利用SSR 分子标记遗传相似系数的聚类结果,从系谱血缘关系及地理远缘关系2 个方面较好地揭示了烤烟资源种内的遗传差异,将69 份种质划分为从古老的Orinoco 衍生出来的优质种质及具有其血缘的后代、国内外地方品种.本研究中2 种方法在类群内划分表现出的差异性较大,亲缘关系较近的品种在利用形态学标记分析中并没有将其划分为一类.如品种Coker319(3 号)和Coker316(7 号)都是从Coker139 与Hicks 的杂交后代中选育出的品种,利用SSR 标记分析将2 个品种划分为一类,而形态学标记将2 个品种划分为完全不同的2 个类群,2个品种的遗传距离为4.14,远大于平均遗传距离2.80.烤烟品种农艺性状特征受环境影响较大,同时由于烤烟品种基本是通过常规杂交选育而成,受育种目标的影响,使育成品种之间类别的划分已不能完全依赖于品种系谱.本研究表明,分子标记技术为确定品种间的遗传差异提供了可靠的检测方法,但在烤烟育种中进行亲本选择时不能仅从亲缘关系上进行考虑,根据育种目标的不同也要把品种在当地气候条件下的农艺性状特征作为选择依据,全面挖掘烤烟材料的育种价值.
[1]卢秀萍.中国烟草品种现状及育种对策[J].西南农业学报,2006,19(增刊):400-404.
[2]王元英,周健.中美主要烟草品种亲源分析与烟草育种[J].中国烟草学报,1995,2(3):11-22.
[3]艾树理.我国烤烟育种进展与问题的探讨[J].中国烟草科学,1999,20(1):44-46.
[4]李永平,马文广.美国烟草育种现状及对我国的启示[J].中国烟草科学,2009,30(4):6-12.
[5]肖炳光,杨本超.利用ISSR 标记分析烟草种质的遗传多样性[J].中国农业科学,2007,40(10):2153-2161.
[6]常爱霞,瞿永生,贾兴华.烟草RAPD 反应体系优化及品种多态性标记研究[J].中国烟草科学,2004,25(2):9-13.
[7]刘建丰,王志德,刘艳华,等.应用SRAP 标记研究烟草种质资源的多样性[J].中国烟草科学,2007,28(5):49-53.
[8]王日新,任民,贾兴,等.烟草主要栽培类型的SRAP 标记研究[J].生物技术通报,2009(6):100-104.
[9]龙腾,刘雷,黄玉碧.四川部分晾晒烟种质遗传关系的SRAP 分析[J].作物学报,2009,35(1):173-178.
[10]刘艳华,王志,牟建民,等.不同烟草群体间遗传多样性分析[J].中国烟草科学,2009,30(增刊):19-24.
[11]杨友才,周清明,尹晗琪.烟草种质资源遗传多样性及亲缘关系的AFLP 分析[J].中国农业科学,2006,39(11):2194-2199.
[12]杜传印,刘洪祥,田纪春.部分烟草种质亲缘关系的AFLP 分析[J].作物学报,2006,32(10):1592-1596.
[13]祁建民,王涛,陈顺辉,等.部分烟草种质遗传多样性与亲缘关系的ISSR 标记分析[J].作物学报,2006,32(3):373-378.
[14]梁景霞,祁建民,方平平,等.烟草种质资源遗传多样性与亲缘关系的ISSR 聚类分析[J].中国农业科学,2008,41(1):286-294.
[15]叶兰钦,幸明明,杜金昆,等.SSR 标记应用于烟草品种遗传多样性研究[J].中国农学通报,2009,25(1):56-62.
[16]李凤霞,王卫锋,王鲁,等.烟草属植物遗传多样性和亲缘进化关系的荧光AFLP 分析[J].中国农业科学,2010,43(12):2418-2427.
[17]杨友才,周清明,尹晗琪,等.利用RAPD 和AFLP 标记分析烟草种质资源的遗传多样性[J].农业生物技术学报,2006,14(4):585-593.
[18]陶爱芬,刘中华,祁建民,等.烟草种质资源遗传多样性的IRAP 和ISSR 标记比较分析[J].武汉植物学研究,2009,27(6):589-594.
[19]聂琼,刘仁祥.23 份烟草种质遗传多样性的SSR 和ISSR 标记分析[J].西南农业学报,2011,24(1):15-19.
[20]祁建民,梁景霞,陈美霞,等.应用ISSR 与SRAP 分析烟草种质资源遗传多样性及遗传演化关系[J].作物学报,2012,38(8):1425-1434.
[21]武耀廷,张天真,殷剑美.利用分子标记和形态学性状检测的陆地棉栽培品种遗传多样性[J].遗传学报,2001,28(11):1040-1050.
[22]彭瑜,苏智先,张素兰.利用叶片形态学性状和ISSR 标记检测柚类的遗传多样性[J].西北农林科技大学学报,2008,36(4):105-110.
[23]BINDLER G,HOEVEN R,GUNDUZ I,et al.A microsatellite marker based linkage map of tobacco[J].Theor Appl Genet,2007,114(2):341-349.
[24]王志德,王元英,牟健民,等.烟草种质资源描述规范和数据标准[M].北京:中国农业出版社,2006:3-40.
[25]ROHLF F J.NTSYS-pc:Numerical taxonomy and multivariate analysis system,Version2.0[M].New York:Exeter Software,Setauket,1988:34-48.
[26]佟道儒.烟草育种学[M].北京:中国农业出版社,1997:15-45.
[27]BINDLER G,PLIESKE J,BAKAHER N,et al.A high density genetic map of tobacco(Nicotiana tabacum L.)obtained from large scale microsatellite marker development[J].Theor Appl Genet,2011,123(2):219-230.
[28]吴海乔.烟草青枯病抗性基因连锁分子标记的检测[D].福州:福建农林大学,2010.
[29]蒋彩虹,罗成刚,任民,等.一个与净叶黄抗赤星病基因紧密连锁的SSR 标记[J].中国烟草科学,2012,33(1):19-22.
[30]聂琼,刘仁祥.同科植物SSR 引物在烟草遗传差异分析中的应用研究[J].中国烟草科学,2011,32(2):52-56.
[31]徐军,刘艳华,任民,等.普通烟草种质资源的SSR 标记与指纹图谱分析[J].中国烟草科学,2011,32(2):62-65.
[32]COSSIRAT J C.Genetic diversity and varietal identification in the species Nicotiana tabacum by RAPD markers[J].Annaesddll Tabac,1994,26:1-7.
[33]郭金平,朱惠丽,周以飞,等.部分烤烟种质遗传多样性与亲缘关系的RAPD 分析[J].中国烟草科学,2009,30(增刊):15-18.
[34]何川生,张汉尧,卢江平,等.RAPD 技术在烤烟品种资源鉴定及纯度分析中的应用[J].河南农业大学学报,2000,34(3):240-243.
[35]肖炳光,卢江平,卢秀萍,等.烤烟品种的RAPD 分析[J].中国烟草学报,2000,6(2):10-15.
[36]杜传印,王玉军,李斯深,等.39 个烤烟种质亲缘关系的AFLP 分析[J].中 国 农 业 科 学,2008,41(9):2741-2747.
[37]杨本超,肖炳光,陈学军,等.基于ISSR 标记的烤烟种质遗传多样性研究[J].遗传,2005,27(5):753-758.
[38]杨春元,任学良,王志红,等.贵州烤烟种质资源主要特征特性的遗传多样性分析[J].中国烟草科学,2009,30(增刊):47-49.
[39]许美玲,卢秀萍.烤烟推广品种亲缘关系分析与育种亲本的选择[J].种子,1999(1):49-51.
