高温环境下三峡库区引进树种水紫树(Nyssa aquatic)的光响应*
2013-07-12王轶浩
王轶浩,罗 韧
(重庆市林业科学研究院,重庆 400036)
三峡工程正常运行后,在库区形成高程落差30 m(高程145 m~175 m)的季节性涨落消落区,它的形成将带来一系列的生态环境问题,诸如水土流失加剧、生物多样性降低、水质污染等,这些都严重影响着三峡水库的生态安全。如何实现三峡库区消落带的生态治理已成为国内外学者关注的焦点[1~3],以往研究通过筛选消落带的适生树(草)种对其生态治理取得一定的试验效果,但这些树(草)种可否在消落带进行大面积示范推广还有待进一步验证。不过可以肯定得是针对消落带严酷的自然环境条件及其对栽植植物的苛刻要求(夏季需耐高温、干旱;冬季需耐水淹),目前所筛选的消落带适生树(草)种种类仍然偏少,对于构建稳定的消落带库岸防护林生态系统还有很大差距,因此,还需加强对消落区适宜树(草)种的选育、引种研究。
水紫树为兰果树科紫树属,是美国特有种,耐水淹,其自然分布在北美东南部以及南部1 000 m 以下的海(河)岸、沼泽地。2009年经美国田纳西州首次引种到三峡库区消落带栽植,经过两年的淹水试验后表现较好,但其对三峡库区夏季高温环境的适应性如何还不甚了解。光合特性作为树种的重要生理指标之一,被广泛地用于表征树种的适应性研究[4],目前国内关于水紫树的光合特性研究也未见报道。本文对水紫树在三峡库区高温环境条件下的光响应进行研究,旨在掌握水紫树引种后对三峡库区高温环境的适应能力以及为丰富消落区的适生树种资源库提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于重庆市林科院的歌乐山试验苗圃,属中梁山山脉,距离沙坪坝区中心——三峡广场约5 km。该苗圃地势较平坦,海拔在500 m 左右。亚热带湿润季风气候,年均气温16.1℃,降水量1 210 mm,年均空气相对湿度84%,日照时数1 115 h。土壤为黄壤,pH 值为6.7,容重为1.16 g·cm-3,总孔隙度为55.67%。
1.2 供试材料
测试材料为水紫树的2 a 生实生移植苗,苗均高60 cm,株行距50 cm×50 cm,常规田间水肥管理。测定时选择一株具有代表性的健康植株,并在其向阳处的中上部挑选3 片生长健康、无机械损害、颜色正常的成熟叶片作为试材。
1.3 光响应曲线测定
采用Li-6400 便携式光合测定仪(美国Li-Cor公司生产),于2012年8月下旬选择光照强度、温度和湿度相对稳定的“桑拿天”进行光响应曲线测定。测定时,利用LED 红蓝光源控制响应因子,设定叶室中光合有效辐射(PAR)梯度为0、50 μmol·m2·s-1、100 μmol·m2·s-1、150 μmol·m2·s-1、200 μmol·m2·s-1、400 μmol·m2·s-1、600 μmol·m2·s-1、800 μmol·m2·s-1、1 000 μmol·m2·s-1、1 200 μmol·m2·s-1和1 500 μmol·m2·s-1,同时,通过系统控制温度为35℃、叶室CO2含量为(390 ±5)μmol·mol-1。每次间隔2.5 min,由仪器自动记录相应的净光合速率(Pn)、蒸腾速率(E)、气孔导度(Gs)、胞间CO2浓度(Ci)等生理指标。
1.4 数据处理
表观光能利用效率(LUE)[5]和表观CO2利用效率(CUE)[6]分别采用如下公式计算:
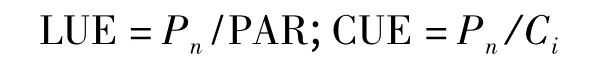
利用Farquhar 的非直角双曲线模型[7-8]来拟合光响应曲线,公式如下:
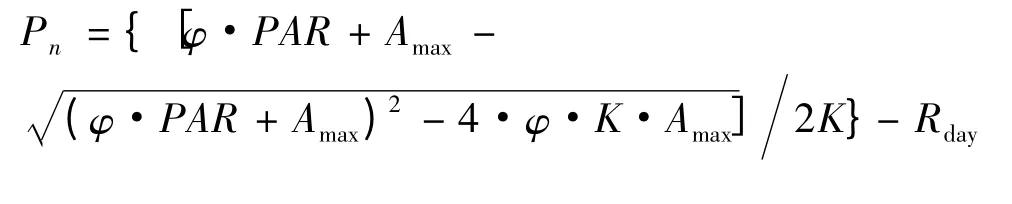
式中,Pn为净光合速率;φ 为表观量子效率;PAR 为光合有效辐射;Amax为最大净光合速率;K 为光响应曲线曲角;Rday为光下呼吸速率。根据上式可求得光补偿点(LCP)和光饱和点(LSP)。
采用Excel 2003 对数据进行图表处理,用SPSS13.0 统计软件进行非直角双曲线方程拟合。
2 结果与分析
2.1 光合光响应曲线
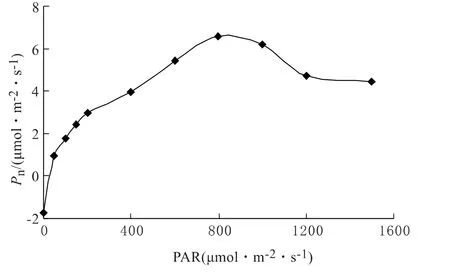
图1 水紫树的光合光响应曲线
植物光合作用受到光照、气温、CO2浓度和空气相对湿度等环境因子的影响,但其中光照为主导因子,它的变化使得其它三者产生相应的变化[9]。由净光合速率的光响应曲线(图1)可知,PAR 为0 时,Pn为负值,这是因为此时水紫树仅有呼吸作用而无光合作用所致。之后Pn随PAR 增大而先迅速再缓慢增加,然后略有下降,说明一开始光合有效辐射强度的增加对净光合速率呈正效应作用,当增加到一定程度时,净光合速率达到峰值(即光饱和点),之后,光合有效辐射强度将对净光合速率产生一定的抑制作用。这是因为PAR 增大能促进叶片的蒸腾速率(图2-a),而蒸腾作用的增强进一步引起水紫树的生理反馈,导致气孔关闭以减少蒸腾失水,气孔导度降低(图2-b),从而净光合速率下降。
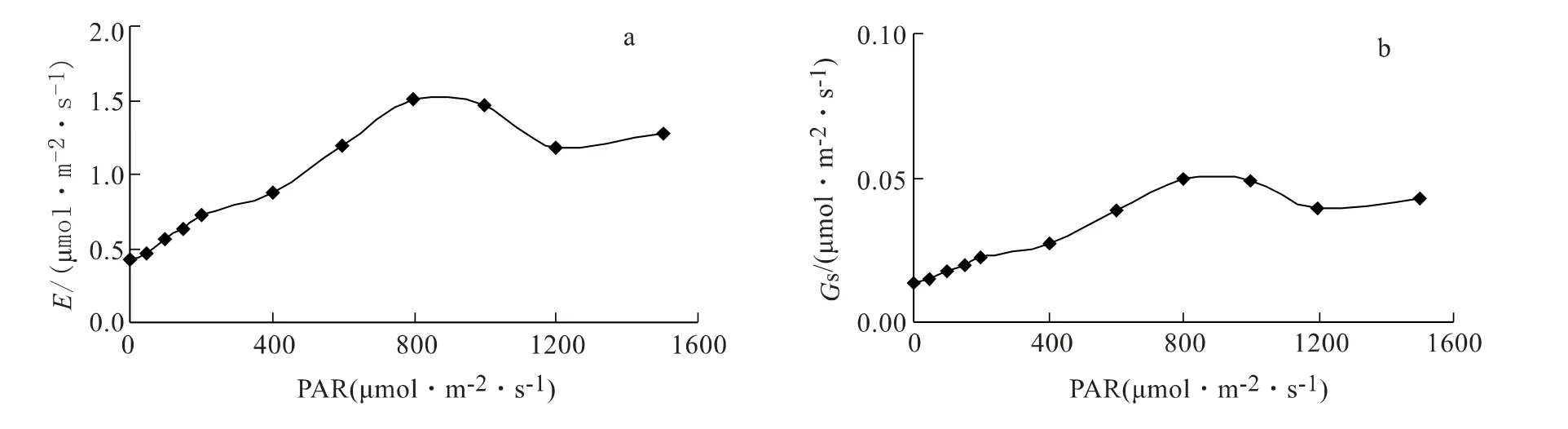
图2 蒸腾速率和气孔导度对光合有效辐射的响应
2.2 蒸腾速率和气孔导度
由图2 看出,蒸腾速率(E)和气孔导度(Gs)随PAR 的变化规律与Pn基本一致,均呈先增加、之后略有下降的趋势,但它们增加的趋势更平缓,这说明E 和Gs对PAR 的响应更缓和。
蒸腾作用能促进植物根系从土壤中吸收水分,同时,也是对外界环境条件的适应性表现,而气孔导度是表示植物气孔传导CO2和水分的能力。在35℃高温条件下,E 随PAR 变化的最高点为1.50 mmol·m2·s-1,Gs的最高点为0.05 mmol·m2·s-1;而在无光照的情况下,E 值为0.42 mmol·m2·s-1,Gs值为0.01 mmol·m2·s-1。
2.3 光能和CO2利用率
水紫树叶片的光能利用率(LUE)随PAR 增加而减小,且在低于400 μmol·m2·s-1时变化更快(图3),可见,水紫树叶片在PAR 较小时,其LUE 较大,反之,则较小。水紫树叶片的CO2利用率(CUE)随PAR 的变化则与Pn一致,也是先迅速再缓慢增加,然后略有下降,说明Pn的光响应变化主要是在PAR 作用下CUE 的变化引起的。
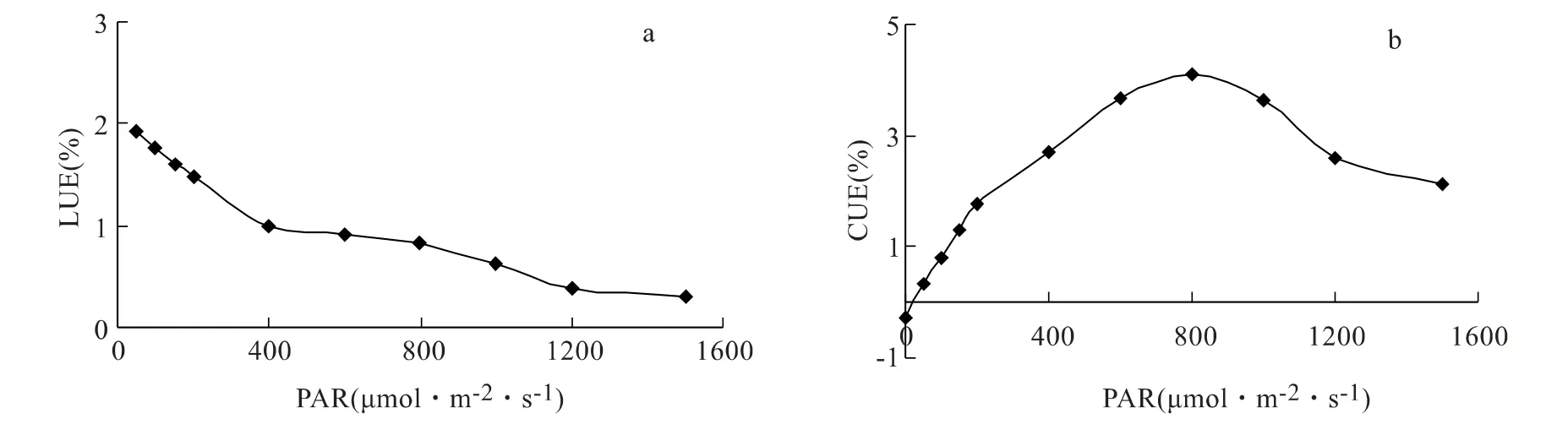
图3 光能和CO2利用率对光合有效辐射的响应
2.4 光响应参数
对水紫树叶片的光合光响应曲线进行非直角双曲线方程拟合,各特征参数见表1。可知,非直角双曲线方程对水紫树的光合光响应拟合效果较好(R2=0.939)。在35℃高温条件下,水紫树的最大净光合速率为6.587 μmol·m2·s-1,说明水紫树对高温环境的适应性较好;LCP 和LSP 分别为16.69、481.82 μmol·m2·s-1,均较小,这说明水紫树的Pn容易达到饱和点,对光强的要求不高且对弱光的利用率较高,耐阴性较强,这也为解释水紫树之所以耐水淹提供了重要科学佐证。

表1 水紫树的光响应(35℃)特征参数
3 结论与讨论
(1)在35℃高温条件下,水紫树的净光合速率、蒸腾速率、气孔导度、光能和CO2利用率对光强的响应明显,除光能利用率随光合有效辐射增加而减小外,其它指标均随光合有效辐射增加而呈先迅速再缓慢增加、然后略有下降的趋势。其最大净光合速率为6.587 μmol·m2·s-1,过高的光强导致蒸腾速率增大、叶片水蒸气压亏缺增大,为防止过度失水,部分气孔关闭、气孔导度下降,净光合速率减小,可见,引进树种水紫树在三峡库区高温条件下仍能发挥正常的光合生理调节反馈机制,适应性较好。
(2)LCP 和LSP是研究植物光合生理特征的两个重要参数,反映植物对弱光和强光的利用能力,它们受到温度明显影响,但温度对其的作用方向仍没有统一,比如谭晓红等[10]在研究普通刺槐和红花刺槐的光响应时发现,高温能降低LSP,但对四倍体刺槐则相反;周洪华等人[8]研究胡杨的光响应时,也发现高温能提高LCP 和LSP,不过研究普遍认为,高温能促使Amax 降低。本研究发现,在35℃高温条件下,水紫树的LCP 和LSP 分别为16.69 μmol·m2·s-1、481.82 μmol·m2·s-1,均低于刺槐、胡杨在高温条件下的LCP 和LSP,而Hands 等[11]研究发现多数植物的光饱和点在500 μmol·m2·s-1~1 000 μmol·m2·s-1,说明水紫树的耐阴性较强且对弱光的利用率较高。
综上所述,水紫树既能适应库区夏季高温条件,又具有较强的耐阴性,能较好的适应库区生态环境,可作为三峡库区生态恢复造林树种。
[1]涂建军,陈治谏,陈国阶,等.三峡库区消落带土地整理利用——以重庆市开县为例[J].山地学报,2002,20(6):712~717.
[2]刘云峰,刘正学.三峡水库消落区极限条件下狗牙根适生性试验[J].西南农业大学学报(自然科学版),2005,27(5):661~663.
[3]王轶浩,耿养会,李彬,等.三峡库区消落带香根草水土保持功能研究[J].四川林业科技,2010,31(2):68~71.
[4]芮雯奕,田云录,张纪林,等.干旱胁迫对6个树种叶片光合特性的影响[J].南京林业大学学报(自然科学版),2012,36(1):68~72.
[5]Nijs I,Ferris R,Blum H.Stomata Regulation in a Changing Climate:A Field Study Using Free Air Temperature Increase (FATI)and Free Air CO2Enrichment[J].Plant,Cell and Environment,1997,20:1041~1050.
[6]何维明,马凤云.水分梯度对沙地柏幼苗荧光特征和气体交换的影响[J].植物生态学报,2000,24(5):630~634.
[7]Farquhar G D,von Caemmerer S,Berry J A.A biochemical model of photosynthetic CO2assimilation in leaves of C3 species[J].Planta,1980,149:78~90.
[8]周洪华,陈亚宁,李卫红,等.干旱区胡杨光合作用对高温和CO2浓度的响应[J].生态学报,2009,29(6):2797~2810.
[9]何斌,王兰珍,刘勇.3个种源异株荨麻光合特性的比较[J].吉林农业大学学报,2010,32(4):355~361,370.
[10]谭晓红,彭祚登,贾忠奎,等.不同刺槐品种光合光响应曲线的温度效应研究[J].北京林业大学学报,2010,32(2):64~68.
[11]Hans M,Peter S.Plant Physiology[J].Economic Botany,1996,50(3):339.