饲食感染SRBSDV 的水稻病株对传毒介体白背飞虱相关能源物质含量的影响
2013-06-11秦小娃董家红张仲凯
张 洁,秦小娃,郑 雪,董家红*,张仲凯*
(1.云南省农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室,昆明 650223;2.中山大学昆虫学研究所/有害生物控制与资源利用国家重点实验室,广州 510275)
南方水稻黑条矮缩病由南方水稻黑条矮缩病毒(Southern rice black-streaked dwarf virus,SRBSDV)引起,于2001年在我国广东省首次发现。2002 至2008年,南方水稻黑条矮缩病主要在华南局部地区为害,近年来,该病在我国南方稻区迅速扩散蔓延,发生面积逐年扩大,呈现出典型的灾变特征(周国辉等,2008;Zhang et al.,2008;Wang et al.,2010)。2009年,该病在我国广东、湖南等9 省(自治区)发生严重(周国辉等,2010;刘万才等,2010);2010年全国累计发病达120万hm2以上。SRBSDV 已经成为影响我国南方水稻生产的重要病害之一(丁铭等,2011;翟保平等,2011)。
SRBSDV属呼肠孤病毒科Reoviridae 斐济病毒属Fijivirus,病毒粒体球状,基因组由10 条dsRNA组成(周国辉等,2008)。SRBSDV 不仅会对水稻造成危害,还可以侵染玉米及多种南方田间常见的禾本科杂草,如稗草Echinochloa crusgalli、牛筋草Eleusine indica 等作为田间重要的中间寄主或侵染源(朱俊子等,2012)。SRBSDV 主要由白背飞虱Sogatella furcifera(Horváth)以持久增殖型方式传毒,白背飞虱一旦获毒,可终身带毒,若虫与成虫均可传毒,并可在水稻生育期的各阶段传毒危害。白背飞虱是一种典型的迁飞性害虫,其主要越冬基地为西南半岛及海南岛,每年早春随西南气流不断北迁,秋季再随东北气流南回(巫国瑞等,1997)。对于我国广大稻区,SRBSDV 初侵染源主要为春季迁入的带毒白背飞虱,病毒在早季稻或其他寄主植物上扩繁后,再经白背飞虱传播侵染单季稻或晚季稻秧苗及分蘖期稻株,造成严重为害(周国辉等,2010)。
糖类、脂肪、蛋白质等是储存于昆虫体内的主要能源物质,昆虫生命活动尤其是昆虫远距离迁飞所需的能量主要由体内的能源物质代谢产生(邹运鼎等,1983)。在昆虫正常的生命活动中,消耗的主要能源物质是糖类。当环境因子发生改变如受杀虫剂胁迫等条件下,糖类、脂肪与蛋白质之间的积累或利用会随之发生转化,从而影响昆虫对不良因子的抵抗能力。
目前,病毒及其感病植株对其传毒介体昆虫生理及代谢影响的研究较少。有研究表明,饲食感染中国番茄黄曲叶病毒(TYLCCNV)的烟草和棉花后,烟粉虱分泌的蜜露中氨基酸和糖含量会发生不同程度的变化(Wang et al.,2012)。然而,SRBSDV 作为一种病毒因子侵染水稻后,对其传毒介体白背飞虱体内相关能源物质影响的研究尚未见报道,SRBSDV 发生流行与其传毒介体白背飞虱暴发的区域性灾变机制尚不明确,饲食感染SRBSDV 水稻病株对白背飞虱远距离迁飞的影响尚不清楚。本研究旨在探讨饲食感染SRBSDV 水稻病株对白背飞虱体内可溶性蛋白、糖类、游离氨基酸及甘油酯含量的影响,希望为揭示饲食感染SRBSDV 水稻病株对白背飞虱体内生理代谢的影响提供实验依据,为进一步明确SRBSDV 流行与白背飞虱暴发的区域性灾变机制提供理论依据。
1 材料与方法
1.1 供试虫源、供试毒源
白背飞虱采自云南元江稻田实验基地,在云南省农科院生物技术与种质资源研究所不接触任何农药等外界刺激的条件下饲养15代以上供试,饲养水稻品种为感虫品种“TN1”。饲养条件为(25±1)°C、14L∶10D,湿度为(70±5)%。
分别于2011年和2012年在云南芒市稻田内将疑似感染SRBSDV 的水稻(品种为滇屯502)整株拔起,保持鲜活,移栽至实验室内,取幼嫩叶组织立即检测或保存于-20℃冰箱待检。所有样本经RT-PCR 检测,序列测定确定后,备用。
1.2 饲食感染SRBSDV 水稻病株条件下白背飞虱体内能源物质含量的测定
于生长期相同(分蘖期)的感染SRBSDV 及健康的水稻茎杆上(对照)接入健康白背飞虱3龄若虫50头、200头、200头,分别饲养至5龄若虫及3日龄和5日龄雌成虫后,各取30头测定若虫与雌成虫体内可溶性蛋白、糖类、游离氨基酸及甘油酯含量。
具体接虫方法为:将处于分蘖期感染SRBSDV水稻或健康水稻(水稻品种均为滇屯502)连根拔出,洗净根部泥土,剪成约10 cm 长的带根稻茎,每3株一组。将其放入培养杯中(培养杯为500mL底部开小口、四周用大头针扎孔的一次性塑料杯)。将靠近根部的水稻茎部用海绵包住穿过培养杯底部小孔,套在装有水稻营养液的200mL的一次性塑料杯中,使水稻根部浸泡在营养液中保湿。约20 min 后,接入白背飞虱3龄若虫。静置约10 min 后剔除受伤个体并补足虫口数量。3 d左右更换一次水稻茎杆。饲养条件为(25±1)℃、14L∶10D,湿度为(70±5)%。实验均重复5次(分别对白背飞虱各种群取虫5次测定各能源物质含量)。各能源物质含量的测定均参照张洁等(2011)的方法进行。
1.2.1 可溶性糖含量的测定
采用蒽酮法测定白背飞虱体内可溶性糖的含量。(1)标准曲线的制定。配制浓度为100 mg/L的葡萄糖标准溶液,分别取0、0.2、0.4、0.6、0.8和1.0mL 于试管中,然后加蒸馏水至1mL,加5mL 蒽酮试剂,混匀,沸水浴加热10 min,取出后在自来水中冷却20 min。用752N 型分光光度计(下同)于620 nm 处测定OD 值,绘制葡萄糖浓度标准曲线。(2)样品的测定。每个处理取白背飞虱30头,称重,加1mL 30% KOH 溶液研磨,沸水浴30 min;加入3mL 无水乙醇,混匀,4500 r/min 离心5 min,取上清液,加3mL 0.6 mol/l盐酸溶液,沸水浴2 h,过滤,加入5mL蒽酮试剂按上述方法测定糖含量。
1.2.2 甘油酯含量的测定
采用索氏脂肪提取法提取白背飞虱体内总的脂类物质。然后将抽提的脂类物质经过皂化、氧化和显色等步骤,最终生成物在420 nm 处比色,测定OD 值。以三油酸甘油酯标准液作对照,计算甘油酯的含量。
1.2.3 游离氨基酸含量的测定
采用茚三酮法测定白背飞虱体内游离氨基酸的含量。(1)标准曲线的制作。配制0.6 mmol/L的标准谷氨酸溶液,分别取0、0.2、0.4、0.6、0.8、1.0mL 于试管中,加蒸馏水补足至1.0mL。各加入1mL pH5.4、2 mol/L 醋酸缓冲液,再加入1.0mL 茚三酮显色液,充分混匀,沸水浴15 min,用自来水冷却。放置5 min 后,加入3.0mL 60%乙醇稀释,充分摇匀,570 nm 处比色测定OD 值(脯氨酸和羟脯氨酸与茚三酮反应呈黄色,应于440 nm 处测定OD 值)。(2)样品的测定。各处理取白背飞虱30头,称重后加入1.2mL 蒸馏水研磨均匀得到样品液。取样品液1.0mL,加入pH5.4、2.0 mol/L 醋酸缓冲液1.0mL和茚三酮显色液1.0mL,混匀后于沸水浴中加热15 min,自来水冷却。放置5 min 后,加3mL 60%乙醇稀释,摇匀后于570 nm 处比色测定OD 值。
1.2.4 可溶性蛋白含量的测定
采用考马斯亮蓝G-250 法测定白背飞虱体内可溶性蛋白的含量。(1)标准曲线制定,配制1.0 mg/mL的标准牛血清蛋白溶液,用蒸馏水稀释成100 ug/mL 的标准蛋白溶液。在5mL 离心管内各加入0、0.1、0.2、0.3、0.4、0.5mL 标准蛋白溶液,用蒸馏水补足至至0.5mL,每管加考马斯亮蓝G-250 2.5mL,混合摇匀,放置5 min,在595 nm 波长下测OD 值。(2)样品测定。于各管中加入酶原0.1mL、蒸馏水0.4mL、考马斯亮蓝G-250 2.5mL,混合摇匀,放置5 min 后于595 nm波长下测定OD 值。
1.3 数据分析
所有数据均采用SPSS17.0 软件进行t 测验分析。
2 结果与分析
2.1 饲食感染SRBSDV 水稻病株对白背飞虱体内可溶性糖含量的影响
饲食感染SRBSDV 水稻病株对白背飞虱体内可溶性糖含量的影响如图1 所示。由图1 可以看出,与对照(健康水稻饲养)相比,饲食感染SRBSDV 水稻病株对白背飞虱5龄若虫体内可溶性糖含量影响不明显;但饲食感染SRBSDV 水稻病株使白背飞虱3日龄和5日龄雌成虫体内可溶性糖含量明显上升,分别是对照组含量的1.18和1.21倍,均显著高于对照组(P<0.05)。
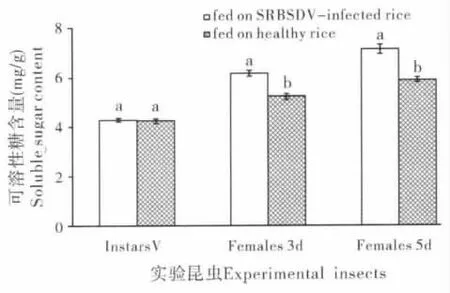
图1 饲食感染SRBSDV水稻病株及健康水稻对白背飞虱体内可溶性糖含量的影响Fig.1 The influence of feeding on SRBSDV-infected rice and healthy rice on soluble sugar contents of Sogatella furcifera
2.2 饲食感染SRBSDV 水稻病株对白背飞虱体内甘油酯含量的影响
由图2 可以看出,饲食感染SRBSDV 水稻病株对白背飞虱体内甘油酯含量的影响因白背飞虱虫龄的不同而不同。饲食感染SRBSDV 水稻病株后白背飞虱5龄若虫和3日龄成虫体内甘油酯含量与对照组相比差异不显著(P>0.05);而白背飞虱5日龄雌成虫体内甘油酯含量则有明显升高,是对照组的1.33倍,达到显著水平(P<0.05)。
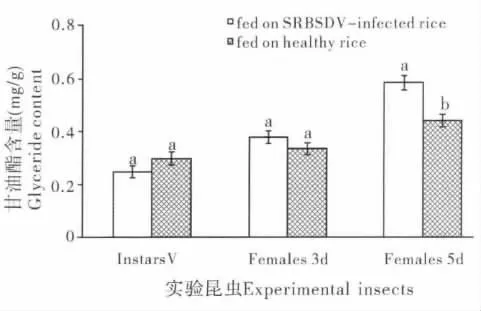
图2 饲食感染SRBSDV 水稻病株及健康水稻对白背飞虱体内甘油酯含量的影响Fig.2 The influence of feeding on SRBSDV-infected rice and healthy rice on glyceride contents of Sogatella furcifera
2.3 饲食感染SRBSDV 水稻病株对白背飞虱体内游离氨基酸含量的影响
饲食感染SRBSDV 水稻病株对白背飞虱体内游离氨基酸含量的影响如图3 所示。由图3 可以看出,饲食感染SRBSDV 水稻病株使白背飞虱3日龄和5日龄雌成虫体内游离氨基酸含量明显升高(P<0.05)分别是对照组的1.46和1.63倍,均达到显著水平(P<0.05),而对白背飞虱5龄若虫体内游离氨基酸含量影响不显著(P>0.05)。
2.4 饲食感染SRBSDV 水稻病株对白背飞虱体内可溶性蛋白含量的影响
饲食感染SRBSDV 水稻病株对白背飞虱体内可溶性蛋白含量的影响如图4 所示。可以看出,饲食感染SRBSDV 水稻病株对白背飞虱5龄若虫和3日龄雌成虫体内可溶性蛋白含量的影响与对照组相比差异不显著(P>0.05);而处理组白背飞虱5日龄雌成虫体内可溶性蛋白含量显著升高(P<0.05),是对照组的1.13倍。

图3 饲食感染SRBSDV 水稻病株及健康水稻对白背飞虱体内游离氨基酸含量的影响Fig.3 The influence of feeding on SRBSDV-infected rice and healthy rice on free amino acid contents of Sogatella furcifera
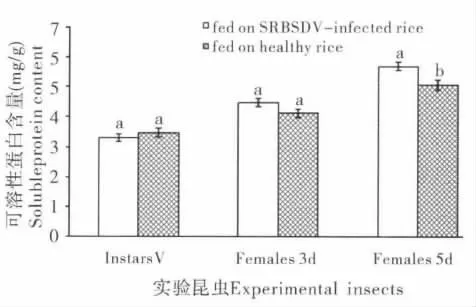
图4 饲食感染SRBSDV 水稻病株及健康水稻对白背飞虱体内可溶性蛋白含量的影响Fig.4 The influence of feeding on SRBSDV-infected rice and healthy rice on soluble protein contents of Sogatella furcifera
3 结论与讨论
许多病毒依赖昆虫传播,病毒对植物的侵染又可对介体昆虫的寿命、产卵量等生物学特性产生影响,这种由寄主植物介导的介体昆虫与病毒的互作可能是引起田间病害流行和介体昆虫种群数量变化的重要因素(Stout et al.,2006)。介体昆虫—病毒互作包括直接和间接两个方面,直接互作主要包括介体昆虫对病原物的传播及病原物在昆虫体内循环、复制等活动对介体昆虫的影响,而间接互作主要指寄主植物感染病毒后对介体昆虫产生的影响(Jiu et al.,2007)。有报道称,饲食感病植株的蚜虫生殖力提高可能与感病植株韧皮部汁液中个别氨基酸含量升高密切相关(Wilkinson and Ishikawa,1999;Sandstrom,2000);感染东非木薯花叶病毒的植株上烟粉虱种群增长迅速,很可能是由于感病植株韧皮部汁液中个别游离氨基酸浓度升高所致(Colvin et al.,2006)。由SRBSDV 引起的南方水稻黑条矮缩病是近些年在我国南方稻区发生流行的一种新病害,其危害逐渐加重的原因与白背飞虱的暴发流行密切相关。
在昆虫正常的生命活动中,糖类是昆虫活动时首先动用的能源物质,也是最主要能量提供者,可直接被代谢供组织利用,亦可将糖转变为海藻糖和糖原,并在需要时相互转化(Wyatt,1967;胡美英等,2002);在远距离迁飞或饥饿等特殊条件下,脂肪和氨基酸会被动员和消耗。脂肪是昆虫体内的主要储备能源和重要的营养物质,在昆虫飞行运动过程中可提供能量,甘油酯是脂肪储存的主要形式(王荫长等,1994)。蛋白质是维系昆虫结构与功能的重要物质,在昆虫生长发育过程中,经过蛋白酶作用可分解成各种氨基酸。在外界不良环境因子胁迫下,糖类、脂肪与蛋白质之间可发生相互转化,从而影响昆虫对不良因子的忍耐能力。目前,关于环境因子对稻飞虱体内能源物质含量影响的研究主要集中于化学杀虫剂方面,如印建莉等(2008)和Yin 等(2008)的研究表明,在经三唑磷和溴氰菊酯处理过的水稻上饲养的褐飞虱,其成虫体内可溶性糖含量和粗脂肪含量有显著差异,亚致死剂量的杀虫剂处理能够促进褐飞虱体内生化物质的积累。
本研究结果表明,饲食感染SRBSDV 水稻病株后,白背飞虱5龄若虫的可溶性糖含量虽高于对照组,但差异不显著,这可能是5龄若虫体内可溶性糖向脂类转化的结果;而其3日龄和5日龄雌成虫体内可溶糖的含量显著升高,说明SRBSDV作为一种病毒因子能够刺激白背飞虱雌成虫对糖的吸收。饲食感染SRBSDV 水稻病株对白背飞虱体内游离氨基酸含量的影响与对可溶性糖含量的影响相似,对5龄若虫体内游离氨基酸含量的影响不显著,但均显著提高了3日龄和5日龄雌成虫体内的游离氨基酸含量。
甘油酯含量的测定结果表明,白背飞虱5龄若虫及3日龄雌成虫体内甘油酯的含量与对照组无显著性差异;而5日龄雌成虫体内甘油酯的含量则显著高于对照。这可能是由于饲食感染SRBSDV 水稻病株对白背飞虱的影响具有时间效应,即随着白背飞虱携毒时间的延长,SRBSDV 在其体内大量积累,导致甘油酯含量的变化显著;也可能是由于若虫通过增加吸收可溶性糖、氨基酸等能量物质与大量消耗脂类相结合的方法来提高代谢活性,而雌成虫则更倾向于增加体内脂类物质的储存以应对不利环境。饲食感染SRBSDV水稻病株对白背飞虱体内可溶性蛋白含量的影响与对甘油酯含量的影响相似,显著提高了5日龄雌成虫体内的可溶性蛋白含量,这可能是由于白背飞虱获取的氨基酸除一部分用于代谢外,更多的部分则用于合成体内蛋白和脂类物质。
昆虫飞行所利用的能源物质主要有三类:碳水化合物、脂类和蛋白质类,不同昆虫飞行期间所消耗的能源物质种类及其过程有所不同(Hoffmann,1985)。脂类是飞虱远距离迁飞的主要能量提供者,糖类和氨基酸在飞行初期提供能量(陈若篪,1983)。对2010年全国各地入侵代白背飞虱的带毒率进行监测的结果表明,在北迁过程中,白背飞虱群体带毒率呈不断上升趋势,暗示白背飞虱带毒对迁飞能力没有不利影响(翟保平等,2011)。本研究结果表明,饲食感染SRBSDV水稻病株会促进白背飞虱雌成虫体内糖类、脂类、氨基酸等物质的积累,有可能可导致其迁飞能力的提升。
本课题组前期的研究结果表明,饲食感染SRBSDV 水稻病株会诱导白背飞虱长翅型成虫的产生(另文发表),作为SRBSDV 的主要传毒介体,白背飞虱长翅型成虫数量的增多和与迁飞相关的体内糖类、脂类、氨基酸等能源物质的积累,增加了SRBSDV 的暴发流行的可能性。
References)
Chen RC,1983.Studies on lipids as fuel of flight in the brown planthopper(Nilaparvata lugens Stål).Acta Entomologica Sinica,26(1):42-48.[陈若篪,1983.脂类作为褐飞虱飞行能量来源的研究.昆虫学报,26:42-48]
Colvin J,Omongo CA,Govindappa MR,Stevenson PC,Maruthi MN,Gibson G,Seal SE,Muniyappa V,2006.Host-plant viral infection effects on arthropod-vector population growth,development and behaviour,management and epidemiological implications.Advances in virus research,67:419-452.
Ding M,Yin YY,Fang Q,Wei JG,Gu ZL,Li HX,Lu X,Zhao J,Zhang ZG,2011.Detection of Southern rice black streaked dwarf virus(SRBSDV) in rice in Yunnan Province.Acta Phytopathologica Sinica,41(6):640-644.[丁铭,尹跃艳,方琦,韦加贵,顾中量,李红祥,卢训,赵健,张仲凯,2011.云南水稻上检测到南方水稻黑条矮缩病毒,植物病理学报,41(6):640-644]
Hoffmann KH,1985.Environment Physiology and Biochemistry of Insects.New York:Springer Berlin Heidelberg Press.112-118.
Hu MY,Zhong GH,Lin JT,Effects of Rhodojaponin-Ⅲon the Energetic Contents against Imported Cabbage Worm(Pieris rapae L.).Journal of Huazhong Agricultural University,21:338-342.[胡美英,钟国华,林进添,孙之潭,2002.闹羊花素-Ⅲ对昆虫血淋巴能源物质含量的影响.华中农业大学学报,21:338-342]
Jiu M,Zhou XP,Tong L,Xu J,Yang X,Wan FH,Liu SS,2007.Vector-Virus mutualism accelerates population increase of an invasive whitefly.PLoS ONE,1:e182.
Liu WC,Liu Y,Guo R,2010.Occurring characteristics and controlling strategy of Southern rice black-streaked dwarf virus(SRBSDV).China Plant Protection,30(3):17-18.[刘万才,刘宇,郭荣,2010.南方水稻黑条矮缩病发生现状及防控对策.中国植保导刊,30(3):17-18]
Sandstrom J,2000.Nutritional quality of phloem sap in relation to host plant-alternation in the bird cherry-oat aphid.Chemoecology,10:17-24.
Stout MJ,Thaler JS,Thomma BPHJ,2006.Plant-mediated interactions between pathogenic microorganisms and herbivorous arthropods.Annual Review of Entomology,51:663-689.
Wang YC,Fan JQ,Tian XZ,Gao BZ,Fan YR,1994.Studies on the resurgent question of planthoppers induced by Deltamethrin and methamidophos.Entomological Knowledge,31:257-262.[王荫长,范加勤,田学志,高保宗,范岳荣,1994.溴氰菊酯和甲胺磷引起稻飞虱再猖獗问题的研究.昆虫知识,31:257-262]
Wang J,Li BX,Li M,Ye GY,Liu SS,2012.Infection of tobacco plants by a begomovirus improves nutritional assimilation by a whitefly.Entomologia Experimentalis et Applicata,144:191-201.
Wang Q,Yang J,Zhou GH,Zhang HM,Chen JP,Adams MJ,2010.The complete genome sequence of two isolates of Southern rice black streaked dwarf virus,a new Fijivirus.Journal of Phytopathology,158:733-737.
Wilkinson TL,Ishikawa H,1999.The assimilation and allocation of nutrients by symbiotic and aposymbiotic pea aphids,Acyrthosiphon pisum.Entomologia Experimentalis et Applicata,91:195-201.
Wu GR,Yu XP,Tao LY.1997.Long-term Forecast on the Outbreak of Brown Planthopper(Nilaparvata lugens Stål)and white-backed Planthopper(Sogatella furcifera Horvath).Scientia Agricultura Sinica,30(4):25-29.[巫国瑞,俞晓平,陶林勇,1997.褐飞虱和白背飞虱灾害的长期预测.中国农业科学,30(4):25-29]
Wyatt GR,1967.The biochemistry of sugars and polysaccharides in insects.Advances in Insect Physiology,4:287-360.
Yin JL,Hu JH,Xu HW,Wu JC,Wang F,Ynang GQ,2008.Comparisons of levels of crude fat,soluble sugars,and free amino acids in offsprings of the immigrant and nonimmigrant populations of the brown planthopper,Nilaparvata lugens(Stål)(Homoptera:Delphacidae),under insecticide stress.Acta Entomologica Sinica,51:1103-1112.[印建莉,胡君欢,徐海伟,吴进才,王飞,杨国庆,2008.杀虫剂胁迫下褐飞虱迁飞虫和本地虫后代体内粗脂肪、可溶性糖及氨基酸含量的比较.昆虫学报,51:1103-1112]
Yin JL,Xu HW,Wu JC,Hu JH,Yang GQ,2008.Cultivar and insecticide applications affect the physiological development of the brown planthopper,Nialaprvata lugens(Stål)(Homoptera:Delphacidae).Environmental Entomology,37:206-212.
Zhai BP,Zhou GH,Tao XR,Chen X,Shen HM,2011.Macroscopic patterns and microscopic mechanisms of the outbreak of rice planthoppers and epidemic SRBSDV.Chinese Journal of Applied Entomology,48(3):480-487.[翟保平,周国辉,陶小荣,陈晓,沈慧梅,2011.稻飞虱暴发与南方水稻黑条矮缩病流行的宏观规律和微观机制.应用昆虫学报,48(3):480-487]
Zhang J,Qin XW,Yuan FH,Zhang RJ,2011.Effects of Sub-Lethal Concentration of Nitenpyram on Soluble Protein,Glyceride,Free Amino Acid and Soluble Sugar of Brown Planthopper,Nilaparvata lugens(Stål)(Homoptera: Delphacidae).Acta Scientiarum Naturalium Universitatis Sun Yat-sen,50(6):20-25.[张洁,秦小娃,袁凤辉,张润杰,2011.亚致死剂量烯啶虫胺对褐飞虱体内脂肪、蛋白质、可溶性糖及游离氨基酸含量的影响.中山大学学报,50(6):20-25]
Zhang HM,Yang J,Chen JP,Adams MJ,2008.A black-streaked dwarf disease on rice in China is caused by a novel fijivirus.Archives of Virology,153:1893-1898.
Zhou GH,Wen JJ,Cai DJ,Li P,Xu DL,Zhang SG,2008.Southern rice black-streaked dwarf virus:A new proposed Fijivirus species in the family Reoviridae.Chinese Science Bulletin,53(23):2500-2508.[周国辉,温锦君,蔡德江,李鹏,许东林,张曙光,2008.呼肠孤病毒科斐济病毒属一新种:南方水稻黑条矮缩病毒.科学通报,53(20):2500-2508]
Zhou GH,Zhang SG,Zou SF,Xu ZW,Zhou ZQ,2010.Occurrence and damage analysis of a new rice dwarf disease caused by Southern rice black-streaked dwarf virus.Plant Protection,36(2):235-238.[周国辉,张曙光,邹寿发,许兆伟,周志强,2010.水稻新病害—南方水稻黑条矮缩病发生特点及为害趋势分析.植物保护,36(2):235-238]
Zhu JZ,Zhou Q,Cui Y,Gao BD,2012.New natural host of Southern rice black-streaked dwarf virus.Journal of Hunan Agricultural University,38(1):58-60.[朱俊子,周倩,崔亚,高必达,2012.南方水稻黑条矮缩病毒的新的自然寄主.湖南农业大学学,38(1):58-60]
Zou YD,Chen JC,Wang SH,1983.Study on the relationship between the internal contents of energy substance and the vitality of lipase with the migration of rice leaf roller(Cnaphlocrocis medinalis Guenee)and brown planthopper(Nilaparvata lugens Stål).Acta Ecologica Sinica,3(1):63-68.[邹运鼎,陈基诚,王士槐,1983.稻纵卷叶螟、褐飞虱体内能源物质动态与迁飞关系的研究.生态学报,3(1):63-68]
