训练者和无训练者脂肪氧化动力学与最大脂肪氧化强度研究
2013-05-23李之俊
张 勇,李之俊
运动强度对脂肪动员和氧化利用具有重要作用。在中低强度范围,机体脂肪氧化率随着运动强度的增加逐渐增加,但是,随着运动强度的进一步增加,机体脂肪氧化率可随着运动强度的增加会逐渐下降。在运动强度与脂肪氧化率相互关系之间存在诱导脂肪氧化率出现最大值的运动强度,称为脂肪最大氧化强度(The exercise intensity eliciting maximal fat oxidation,FATmax)[23],FATmax 在 脂 肪代谢的运动干预实践应用中具有重要作用,已成为研究热点[15,16,51]。以前的研究大多是在中等强度范围内预先给定3~5个不同强度,然后通过比较相应几个强度下的机体脂肪氧化率来确定脂肪最大氧化的运动强度,很明显,这些研究涉及的强度范围并不完整,研究结果的准确性有待商榷[2,40,43]。近几年来,有人开始尝试在更广的强度范围通过一次递增负荷运动对脂肪氧化动力曲线和FATmax进行研究,发现在相同运动强度下,3阶数多项式(P3)曲线和正弦(SIN)曲线上对应的脂肪氧化率与实测值之间无显著性差异[14,39,48]。有人对 FATmax 测试方案也进行了研究,证明了一次递增负荷模式(每级负荷持续3min)和分多次递增负荷模式(每次持续30min以内)测试的FATmax无显著性差异[2,16,52],证明了采用每级持续3min以上的递增负荷方案确定FATmax是可靠的。这些研究结果为采用一次递增负荷模式在完整强度范围研究运动对脂肪氧化动力曲线和FATmax提供了依据,但遗憾的是仍鲜见相关研究的报道。另外,从相关理论和已有研究来看,FATmax与运动者的年龄、性别、体能水平、训练背景有一定 关 系[8,12,17,21,39,43,48],不 同 人 群 的 FATmax 可 能 存 在 一 定差异,研究不同人群的FATmax对脂肪代谢运动干预具有重要实践意义。本研究试图采用一次递增负荷模式对有训练者和无训练者的FATmax进行测试,并对其结果进行比较分析,借以间接了解长期训练对脂肪氧化动力学的影响,为运动健康促进的实践应用提供一定参考。
1 研究对象与方法
1.1 研究对象
研究对象为无训练健康女生12名(每周运动2h以下)和有训练健康女生12名(每周运动8h以上;表1)。
表1 研究对象基本资料一览表Table 1 Characteristics of the Subjects(±SD)
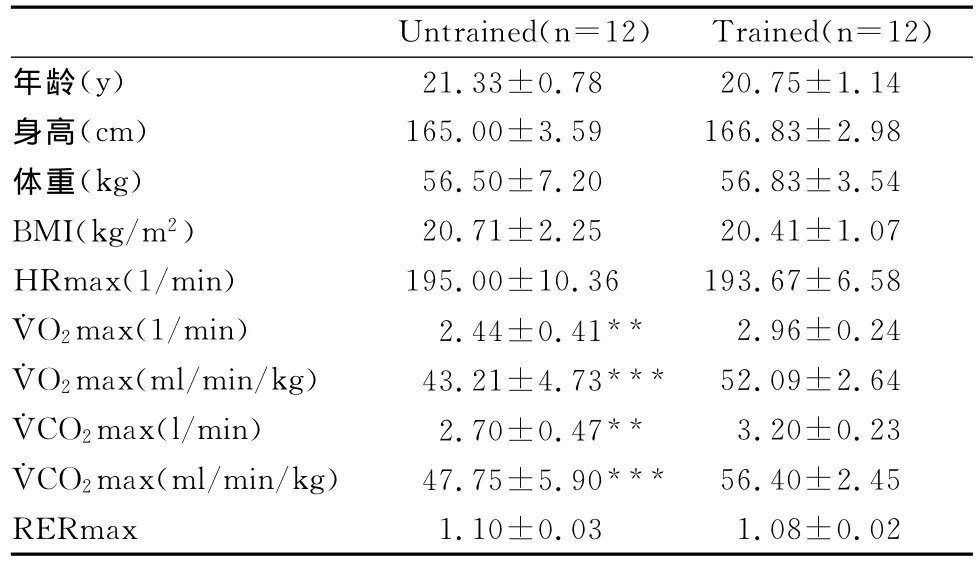
表1 研究对象基本资料一览表Table 1 Characteristics of the Subjects(±SD)
注:HRmax,最大心率;RERmax,最大呼吸商;与训练者比较,***P<0.001,**P<0.01。
Untrained(n=12) Trained(n=12)年龄(y) 21.33±0.78 20.75±1.14身高(cm) 165.00±3.59 166.83±2.98体重(kg) 56.50±7.20 56.83±3.54 BMI(kg/m2) 20.71±2.25 20.41±1.07 HRmax(1/min) 195.00±10.36 193.67±6.58 ˙VO2max(1/min) 2.44±0.41** 2.96±0.24 ˙VO2max(ml/min/kg) 43.21±4.73*** 52.09±2.64 ˙VCO2max(l/min) 2.70±0.47** 3.20±0.23 ˙VCO2max(ml/min/kg) 47.75±5.90*** 56.40±2.45 RERmax 1.10±0.03 1.08±0.02
1.2 研究程序和方案
实验研究之前测试身高、体重、肺功能、血压、心电图等,调查本人及家族心血管疾病情况、运动经历,以保证运动的安全性。调查研究对象月经周期,以确保相关测试在卵泡期早期(月经周期3~8天)进行,减少雌激素水平对脂代谢的影响。向受试者介绍实验过程和相关注意事项,征得同意并签订协议书。整个运动实验在清晨空腹状态进行。在实验期间,研究对象需完成2个运动测试。实验前1周熟悉实验程序。正式实验期间不吸烟、不喝酒、不喝咖啡,保证正常饮食,未服任何治疗药物,女性未服避孕药。测试前48h不得参加剧烈运动。
1.2.1 最大耗氧量测试
采用德国h/p/cosmos公司 mercury 4.0跑台和CORTEX MetaMax 3B心肺功能仪完成最大耗氧量测试。受试者早晨缓慢步行到实验室,准备活动5min,随后采用气体代谢仪进行最大耗氧量测试。室内温度控制在25℃,相对湿度为40%。无训练者测试方案为5km/h热身5min,然后6km/h起,递增幅度1km/2min,至10km/h后,速度不变,坡度每2min递增1%。有训练者测试方案为5 km/h热身5min,然后8km/h起,递增幅度1km/2min,至11km/h后,速度不变,坡度每2min递增1%。
最大耗氧量通过下列标准确定:1)心率达到年龄预测最大心率的±10次/min;2)呼吸商大于1.10;3)随着负荷的增加,耗氧量停止线性增加并达到一个平台或开始缓慢下降,最后两个值的差异小于±2ml/kg/min;4)受试者力竭,经激仍无法保持预定负荷。以上标准满足3个或3个以上即可判断达到˙VO2max。
1.2.2 最大脂肪氧化强度测定
休息至少48h以后,受试者早晨空腹步行至实验室,室内温度控制在25℃,相对湿度为40%。采用德国h/p/cosmos公司mercury 4.0跑台和CORTEX MetaMax 3B心肺功能仪完成递增负荷运动机体气体代谢指标测试,确定最大脂肪氧化强度。测试方案为无训练者5km/h开始,递增幅度为1km/3.5min,保持水平坡度不变,直至呼吸商达1.00以上。有训练者6km/h开始,递增幅度为1 km/3.5min,保持水平坡度不变,直至呼吸商达1.00以上。采用每级负荷后30s气体代谢数据计算脂肪氧化率,通过3阶数多项式拟合曲线确定脂肪氧化动力曲线和FATmax。脂肪氧化率计算公式[37]:脂肪的氧化(g/min)=1.6946˙VO2(L/min)-1.7012˙VCO2(L/min)。
1.3 数据处理
所有数据采用 Matlab R2010a和SPSS 11.5for Win-dows软件进行处理。所有数据采用±SD表达,采用单变量双因素方差分析对不同组和不同强度运动脂肪氧化率的差异进行分析,并对强度之间差异进行事后比较检验(post hoc test);采用独立样本t检验比较相同强度组间脂肪氧化率差异。采用Pearson相关对最大脂肪氧化率和最大脂肪氧化强度与最大耗氧量之间相关性进行分析,所有统计分析显著性水平为P<0.05。
2 研究结果
2.1 不同强度运动脂肪氧化率
方差分析表明,运动强度和训练对脂肪氧化率均有影响(P<0.001)。随着运动强度的增加,脂肪氧化率呈现先增加后减少的变化趋势(表2,图1~图4)。无训练者脂肪氧化率在50%˙VO2max强度之前随着运动强度的增加逐渐增加,在65%˙VO2max强度之后随着运动强度的增加开始明显下降,在50%~65%˙VO2max强度阶段脂肪氧化率呈现相对较高水平,脂肪氧化率在6.36~6.67mg/min/kg之间。训练者脂肪氧化率在60%˙VO2max强度之前随着运动强度的增加逐渐增加,在70%˙VO2max强度之后随着运动强度的增加明显下降,在60%~70%˙VO2max强度阶段脂肪氧化率呈现相对较高水平,脂肪氧化率在12.20~12.84mg/min/kg之间。另外,独立样本t检验结果进一步表明,相同强度运动训练者脂肪氧化率均高于无训练者。
表2 本研究无训练者与训练者不同强度脂肪氧化率测试结果一览表Table 2 Fat Oxidation Rate in Different Exercise Intensity for Trained and Untrained Women%˙VO2max±SD)
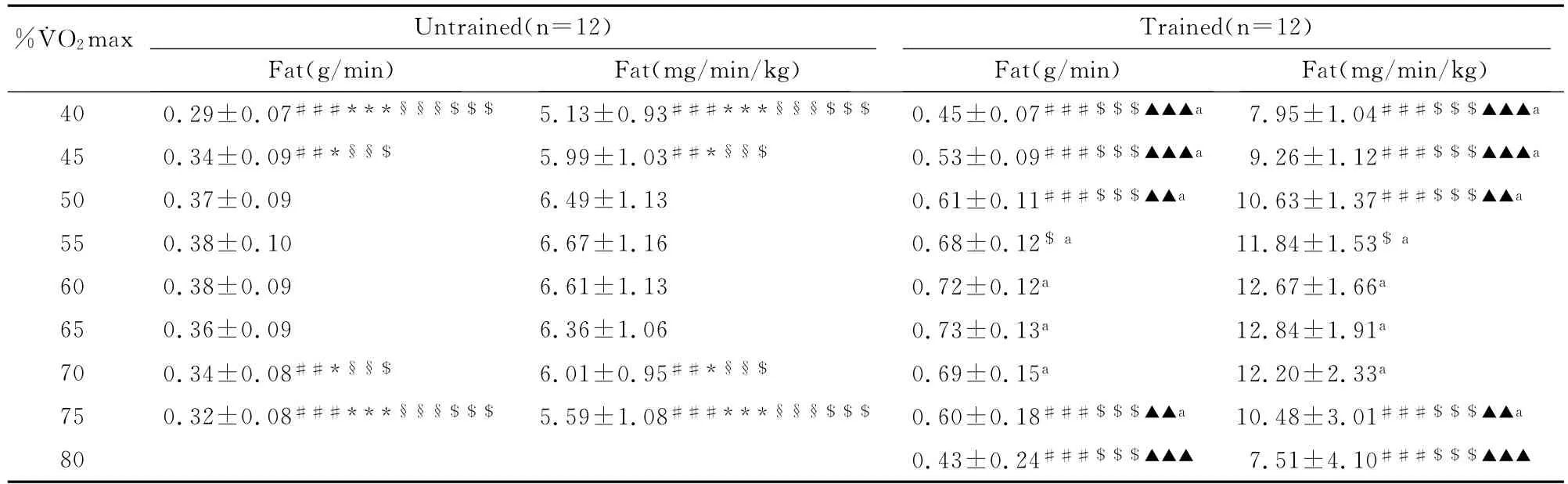
表2 本研究无训练者与训练者不同强度脂肪氧化率测试结果一览表Table 2 Fat Oxidation Rate in Different Exercise Intensity for Trained and Untrained Women%˙VO2max±SD)
注:与50%˙VO2max强度比较,***P<0.001,*P<0.05;与55%˙VO2max强度比较,§§§P<0.001,§§P<0.01;与60%˙VO2max强度比较,###P<0.001,## P<0.01;与65%˙VO2max强度比较,$$$ P<0.001,$ P<0.05;与70%˙VO2max强度比较,▲▲▲P<0.001,▲▲P<0.01;与无训练者比较,aP<0.001。
%˙VO2max Untrained(n=12)Trained(n=12)Fat(mg/min/kg)40 0.29±0.07###***§§§$$$ 5.13±0.93###***§§§$$$ 0.45±0.07###$$$▲▲▲a 7.95±1.04###$$$▲▲▲a Fat(g/min)Fat(mg/min/kg)Fat(g/min)45 0.34±0.09##*§§$ 5.99±1.03##*§§$ 0.53±0.09###$$$▲▲▲a 9.26±1.12###$$$▲▲▲a 50 0.37±0.09 6.49±1.13 0.61±0.11###$$$▲▲a 10.63±1.37###$$$▲▲a 55 0.38±0.10 6.67±1.16 0.68±0.12$a 11.84±1.53$a 60 0.38±0.09 6.61±1.13 0.72±0.12a 12.67±1.66a 65 0.36±0.09 6.36±1.06 0.73±0.13a 12.84±1.91a 70 0.34±0.08##*§§$ 6.01±0.95##*§§$ 0.69±0.15a 12.20±2.33a 75 0.32±0.08###***§§§$$$ 5.59±1.08###***§§§$$$ 0.60±0.18###$$$▲▲a 10.48±3.01###$$$▲▲a 80 0.43±0.24###$$$▲▲▲ 7.51±4.10###$$$▲▲▲
2.2 最大脂肪氧化率与最大脂肪氧化强度
表3的研究结果表明,有训练者最大脂肪氧化率和最大脂肪氧化强度均高于无训练者。训练者最大脂肪氧化强度为63.95%±3.16%˙VO2max,最大脂肪氧化率为13.04±1.82mg/min/kg。无训练者最大脂肪氧化强度为56.51%±2.50%˙VO2max,最大脂肪氧化率为6.71±1.15 mg/min/kg。
考虑到在实践中的应用,从最大脂肪氧化率的角度出发,对不同强度脂肪氧化率均值进行了三项式拟合分析,进一步确定95%最大脂肪氧化率和90%最大脂肪氧化率及所对应的运动强度(表4、图5、图6)。结果表明,无训练者在48.21%~65.41%˙VO2max强度范围,脂肪氧化率即可达到最大脂肪氧化率的95%以上,在45.12%~69.85%˙VO2max强度范围,脂肪氧化率可达到最大脂肪氧化率的90%以上;训练者在57.14%~69.86%˙VO2max强度范围,脂肪氧化率即可达到最大脂肪氧化率的95%以上,在53.78~72.10%˙VO2max强度范围,脂肪氧化率可达到最大脂肪氧化率的90%以上。
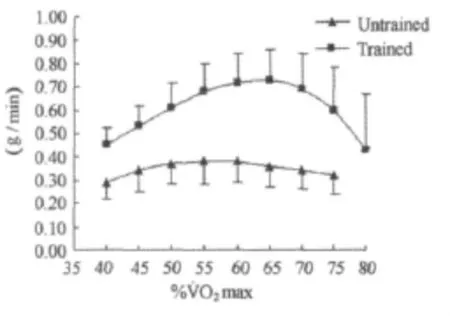
图1 本研究不同强度运动脂肪氧化率曲线图Figure 1. Fat Oxidation Rate in Different Exercise Intensity(g/min)
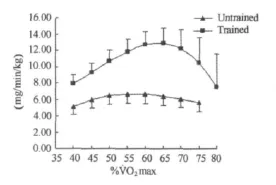
图2 本研究不同强度运动脂肪氧化率曲线图Figure 2. Fat Oxidation Rate in Different Exercise Intensity(mg/mim/kg)
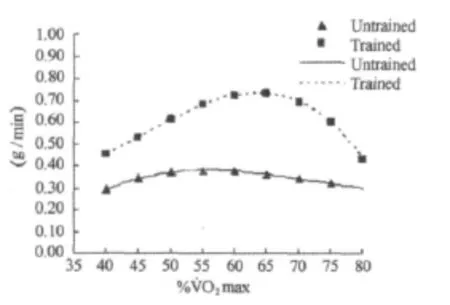
图3 本研究不同强度运动脂肪氧化率P3曲线图Figure 3. The 3rd Polynomial Fitting Curve for Fat Oxidation Rate in Different Exercise Intensity(g/min)
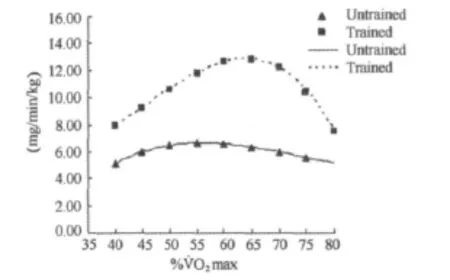
图4 本研究不同强度运动脂肪氧化率P3曲线图Figure 4. The 3rd Polynomial Fitting Curve for Fat Oxidation Rate in Different Exercise Intensity(mg/mim/kg)
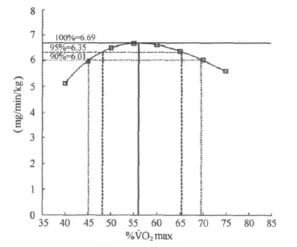
图5 本研究无训练者运动强度与脂肪氧化率P3曲线图Figure 5. The 3rd Polynomial Fitting Curve for Fat Oxidation Rate in Different Exercise Intensity for Untrained Women
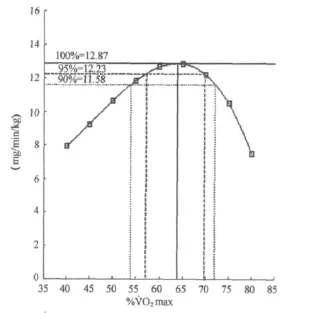
图6 本研究训练者运动强度与脂肪氧化率P3曲线示意图Figure 6. The 3rd Polynomial Fitting Curve for Fat Oxidation Rate in Different Exercise Intensity for Trained Women
2.3 最大脂肪氧化率和最大脂肪氧化强度与最大耗氧量的关系
最大脂肪氧化率和最大脂肪氧化强度与最大耗氧量之间的关系的研究结果表明(表5),最大脂肪氧化率、最大脂肪氧化强度、最大摄氧量之间存在正相关。
表3 本研究跑台运动最大脂肪氧化强度与最大脂肪氧化率测试结果一览表Table 3 Exercise Intensity Eliciting Maximal Fat Oxidation and Maximum Fat Oxidation Rate in Running Exercise(±SD)

表3 本研究跑台运动最大脂肪氧化强度与最大脂肪氧化率测试结果一览表Table 3 Exercise Intensity Eliciting Maximal Fat Oxidation and Maximum Fat Oxidation Rate in Running Exercise(±SD)
注:FATmax,最大脂肪氧化强度,MFO,最大脂肪氧化率,下同;与无训练者相比,***P<0.001。
FATmax(%˙VO2max) FATmax(km/h) MFO(g/min) MFO(mg/min/kg)71±1.15 Trained 63.95±3.16*** 8.52±0.78*** 0.74±0.13*** 13.04±1.82 Untrained 56.51±2.50 7.29±0.68 0.38±0.10 6.***
表4 本研究均值三项式拟合后最大脂肪氧化运动强度及最大脂肪氧化率测试结果一览表Table 4 Exercise Intensity Eliciting Maximal Fat Oxidation and Maximum Fat Oxidation Rate From the 3rd Polynomial Fitting Curve for Fat Oxidation Rate in Different Exercise Intensity (±SD)
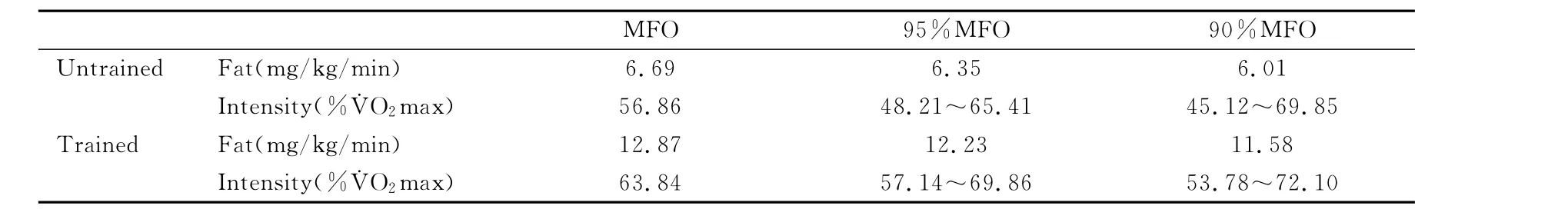
表4 本研究均值三项式拟合后最大脂肪氧化运动强度及最大脂肪氧化率测试结果一览表Table 4 Exercise Intensity Eliciting Maximal Fat Oxidation and Maximum Fat Oxidation Rate From the 3rd Polynomial Fitting Curve for Fat Oxidation Rate in Different Exercise Intensity (±SD)
6.69 6.35 6.01 Intensity(%˙VO2max) 56.86 48.21~65.41 45.12~69.85 Trained Fat(mg/kg/min) 12.87 12.23 11.58 Intensity(%˙VO2max) 63.84 57.14~69.86 53.78~72.10 MFO 95%MFO 90%MFO Untrained Fat(mg/kg/min)
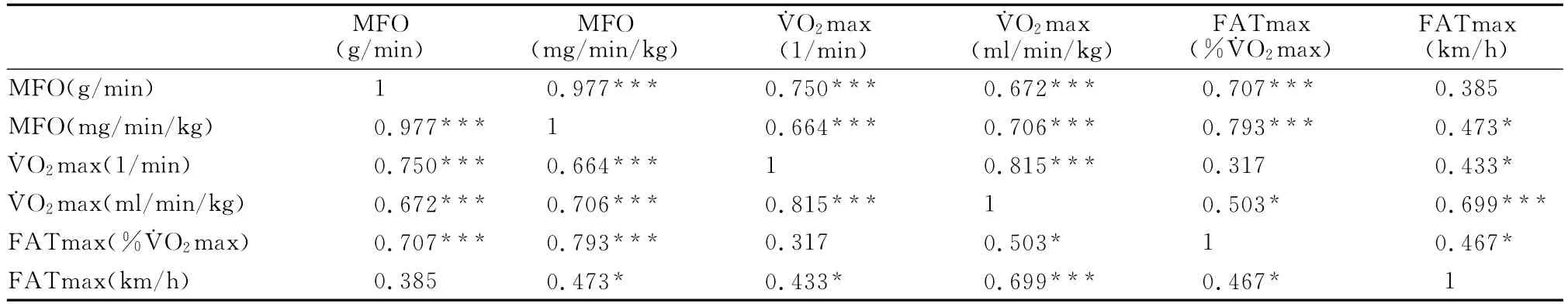
表5 本研究最大脂肪氧化率和最大脂肪氧化强度与最大耗氧量的关系一览表Table 5 Correlation Between the Maximum Fat Oxidation Rate,Exercise Intensity Eliciting Maximal Fat Oxidation and Maximal Oxygen Uptake (n=24)
3 分析与讨论
3.1 不同强度运动脂肪氧化率
本研究表明,运动强度对脂肪氧化率有重要作用,无论训练者还是无训练者,脂肪氧化率随着运动强度的增加均呈现先增加后减少的变化趋势。无训练者脂肪氧化率在50%˙VO2max强度之前随着运动强度的增加逐渐增加,在65%˙VO2max强度之后随着运动强度的增加开始明显下降,在50%~65%˙VO2max强度阶段脂肪氧化率呈现相对较高的稳定状态。训练者脂肪氧化率在60%˙VO2max强度之前随着运动强度的增加逐渐增加,在70%˙VO2max强度之后随着运动强度的增加明显下降,在60%~70%˙VO2max强度阶段脂肪氧化率呈现相对较高的状态。
从运动与机体能耗及底物代谢的角度分析,认为在中低强度范围运动,随着运动强度的逐渐增加,脂肪参与供能比例逐渐减少,但机体的能量需求增加更加明显[7],脂肪氧化率仍然呈现逐渐升高趋势,但随着运动强度的进一步增强,机体总能耗急剧增加,脂肪氧化供能速度较慢,已跟不少机体需要,以糖为底物的快速供能系统参与供能比例急剧增加,以满足机体的能量需求,脂肪氧化率开始急剧下降。运动科学家把运动强度由低到高逐渐增加过程中脂肪氧化率出现最高值时对应的运动强度,也就是促使机体出现脂肪氧化利用最大化的运动强度称为最大脂肪氧化运动强度(intensity eliciting maximal fat oxidation),Jeukendrup和Achten把它定义为FATmax[23],FATmax 在运动实践中具有重要的应用价值。
那么,这种不同运动强度下脂肪参与氧化供能水平的调节机制是什么?从现有文献来看,认为肌肉收缩、神经内分泌反应、血液循环水平的增强、代谢产物等可能在不同强度运动时的脂肪氧化供能水平的调节中具有作用。研究已经表明,肌肉收缩和血液循环水平对脂肪酸转运、摄入和氧化有调节作用。肌肉收缩在诱导质膜位的脂肪酸移位酶(FAT/CD36)、质膜脂肪酸结合蛋白(FABPpm)、脂肪酸转运蛋白(FATP)表达和转运中具有重要作用。肌肉收缩不仅可诱导FABPpm和FAT/CD36向质膜转移,也可诱导FAT/CD36和FABPpm的表达,进而使得脂肪酸摄入 和 氧 化 增 加[11,22,35,44]。 然 而 ,具 体 准 确 的 诱 导 这 种 转 运产生的信号途径还有待进一步解释。现有动物研究提示,AMPK(一磷酸腺苷激活蛋白激酶)可能在运动诱导的转运中具有一定作用[20,29]。神经内分泌反应对脂肪分解具有重要调节作用。运动时肾上腺素、去甲肾上腺素、胰岛素、钠尿肽等激素水平的增加、胰岛素水平的下降都具有促 脂 解 作 用[1,5,10,27,45,53]。
在中低强度范围,随着运动强度的增加,骨骼肌收缩和血液循环水平增加对脂肪氧化利用产生的调节作用增强,脂肪酸摄入和氧化增加。同时,随着运动强度的进一步增加,儿茶酚胺水平升高、钠尿肽水平增加、胰岛素水平下降,神经内分泌调节下的脂解作用增强,脂肪氧化率增加[19,24,26,30]。但是,当运动强度增加至较高水平 时,机体能量需求速率急剧增加,葡萄糖和糖原成为主要的供能底物,同时,随着运动强度的增加,乳酸产生增加,可激活GPR81(与烟酸受体GPR109a高度相似的Gi蛋白耦联受体),进而通过抑制性G蛋白偶联受体通路相关的腺苷酸环化酶 的 下 降 来 抑 制 脂 解[5,28,41]。 而 Starritt[46]和 Bezaire[9]等以前认为,乳酸积累导致PH下降可通过增强丙二酰辅酶A敏感性来抑制肉碱棕榈酰转移酶Ⅰ(carnitine palmitoyltransferase-I,CPTI)活性,进而抑制游离脂肪酸向线粒体的转运。乳酸抑制脂肪氧化利用的主要调节机制还需要进一步研究。另外,高强度运动时脂肪组织血流减少,影响脂肪酸转运,长链脂肪酸进入线粒体减少与脂肪动员减少可 能 也 有 一 定 关 系[35,36,42]。 因 此 ,综 合 认 为 ,随 着 运 动 强度的增加,肌肉收缩和促脂解激素反应增强,脂肪动员、转运、摄入和氧化利用增加。但当运动强度增加到较高水平时,一方面,可导致乳酸浓度升高,对脂解产生强烈抑制作用;另一方面,可导致脂肪组织血流减少,影响脂肪酸转运,影响脂肪动员和利用。综合来看,相关调节机制比较复杂,还需要进一步深入研究。
从表1和图1~图6可知,在相同运动强度(%˙VO2max)下,训练者脂肪氧化率均高于无训练者,而且,随着运动强度的变化训练者脂肪氧化率的变化幅度明显高于无训练者。分析认为,两者出现这样的差异可能与训练者脂肪动员和氧化能力较强有关。而训练导致的脂肪氧化利用能力的增强可能与训练导致的肾上腺素受体敏感度 增 强[18,47]、ANP 活 性 增 强[31,32]、机 体 皮 下 脂 肪 组 织 血 流改善[36]、脂肪酸的转运能力增强、氧化酶活性增强、机体乳酸积累减少等因素有关,也与长期训练导致骨骼肌脂滴包被蛋白(Perilipin)[38]、激素敏感性脂肪酶(HSL)[49]、脂肪甘油三酯酶(ATGL)[6,33]水平上调有关。综合相关理论与已有研究,笔者认为,长期有氧耐力训练提高脂肪分解和氧化供能能力的相关调节机制可能涉及以下几个方面:1)促脂解激素受体敏感度增强与脂肪酶及其相关蛋白表达增加;2)骨骼肌脂肪酸摄入及氧化能力的增强(这可能与相关转运蛋白的表达及线粒体体积和数量增加相关氧化酶活性的增强有关);3)机体心肺功能及摄氧能力的增强;4)脂肪组织血液循环改善,脂肪酸和氧运输增强;5)相同强度运动乳酸生成减少,乳酸对脂解抑制减弱。
3.2 最大脂肪氧化率与最大脂肪氧化强度
本研究结果表明,最大脂肪氧化率训练者高于无训练者(P<0.001),分别为0.74±0.13g/min(13.04±1.82 mg/min/kg)和0.38±0.10g/min(6.71±1.15mg/min/kg),其中,训练者的研究结果与Achten等的研究结果基本一 致[4],但 高 于 其 他人 的 研 究[2,13,34],无 训 练 者 的 研 究 结 果高于 Nordby[34]和 Stisen[48]的研究。另外,最大脂肪氧化强度的研究结果表明,训练者最大脂肪氧化强度高于无训练者,分别为63.95%±3.16%˙VO2max和56.51%±2.50%˙VO2max(P<0.001),其中,训练者的研究结果与 Achten等的研究结果基本一致[3],但高于其他人的研究[2,13],无训练者的研究结果与 Venables等[50]和Stisen[48]等基本一致,但高于Nordby的研究[34]。总的来看,在相同条件下,训练者的最大脂肪氧化率和最大脂肪氧化强度均高于无训练者,这与已有的研究结果一致[8,48]。分析认为,这些研究结果的差异可能与研究对象的不同(男女、年龄、、训练背景等)有关。因为已有研究已经证明,研究对象的性别、年龄以及训练状态会对最大脂肪氧化率和最大脂肪氧化强度 产 生 影 响[8,12,17,21,39,43,48],认 为 女 性 脂 肪 氧 化 利 用 能 力 强于男性,年轻人脂肪氧化利用能力强于老年人,长期耐力训练者脂肪氧化利用能力强于无训练者。另外,与测试时饮食状态(是否空腹)以及研究过程采用的运动方式的不同(自行车、跑台)也有一定关系。因为空腹状态运动可导致脂肪氧化利用增加,跑台运动脂肪氧化利用能力强于自行 车 运 动[13,25]。
另外,本研究发现,无训练者在48.21%~65.41%˙VO2max强度范围,脂肪氧化率即可达到最大脂肪氧化率的95%以上,训练者在57.14%~69.86%˙VO2max强度范围,脂肪氧化率即可达到最大脂肪氧化率的95%以上。提示,在跑台运动过程中,若要达到最大脂肪氧化率的95%以上,无训练者可采用的强度范围较大,但平均运动强度较低。这与脂肪氧化利用能力较低有关。
3.3 最大脂肪氧化率和最大脂肪氧化强度与最大耗氧量的关系
为进一步分析脂肪氧化利用与机体耗氧能力的关系,本研究对机体最大脂肪氧化率和最大脂肪氧化强度与最大耗氧量进行了相关分析。结果发现,最大脂肪氧化率和最大脂肪氧化强度与最大耗氧量存在正相关,这与已有的研究结果一致[25,50]。说明机体耗氧能力越强有助于脂肪的氧化利用。因为至少存在这样的事实,在相同相对强度下,最大耗氧量越高绝对耗氧量越高,如果在呼吸商相同情况下,脂肪氧化率就越高。
本研究作为横向比较研究在解释运动对脂肪氧化利用的影响方面可能还有一定的局限性,今后有必要进行长期运动训练干预下的纵向研究。
4 结论
1.相同强度(%˙VO2max)运动,训练者脂肪氧化率高于无训练者。训练者最大脂肪氧化强度和最大脂肪氧化率也高于无训练者,提示,长期耐力训练可增强机体脂肪动员和氧化利用能力。
2.从实践应用的角度考虑,跑台运动时,无训练者强度在50%~65%˙VO2max范围,有训练者强度在60%~70%˙VO2max范围均可促使机体脂肪氧化率达到最大脂肪氧化率的95%以上。
3.最大脂肪氧化强度、最大脂肪氧化率、最大耗氧量之间存在一定正相关,提示,最大脂肪氧化率和最大脂肪氧化强度与机体耗氧能力和利用氧的能力有一定关系。
[1]张勇,李之俊.运动与脂肪动员研究进展[J].中国运动医学杂志,2012,31(1):69-75.
[2]ACHTEN J,GLEESONl M,JEUKENDRUP AE.Determination of the exercise intensity that elicits maximal fat oxidation[J].Med Sci Sports Exe,2002,34(1):92-97.
[3]ACHTEN J,JEUKENDRUP A E.Maximal fat oxidation during exercise in trained men[J].Int J Sports Med,2003,24(8):603-608.
[4]ACHTEN J,VENABLES M C,JEUKENDRUP A E.Fat oxidation rates are higher during running compared with cycling over a wide range of intensities[J].Metabolism,2003,52(6):747-752.
[5]AHEMD K,TUNARU S,TANG C,et al.An autocrine lactate loop mediates insulin-dependent inhibition of lipolysis through GPR81[J].Cell Metab,2010,11(4):311-319.
[6]ALSTED T J,NYBO L,SCHWEIGER M,et al.Adipose triglyceride lipase in human skeletal muscle is upregulated by exercise training[J].Am J Physiol Endocrinol Metab,2009,296(3):E445-453.
[7]AUCOUTURIER J,BAKER J S,DUCHE P.Fat and Carbohydrate Metabolism during Submaximal Exercise in Children[J].Sports Med,2008,38(3):213-238
[8]BASSAMI M,AHMADIZAD S,DORAN D,et al.Effects of exercise intensity and duration on fat metabolism in trained and untrained older males[J].Eur J Appl Physiol,2007,101(4):525-532.
[9]BEZAIRE V,HEIGENHAUSER G J,SPRIET L L.Regulation of CPT I activity in intermyofibrillar and subsarcolemmal mitochondria from human and rat skeletal muscle[J].Am J Physiol Endocrinol Metab,2004,286(1):E85-91.
[10]BEZAIRE V,LANGIN D.Regulation of adipose tissue lipolysis revisited[J].Proc Nutr Soc,2009,68(4):350-360
[11]BRADLEY N S,SNOOK L A,JAIN S S,et al.Acute endurance exercise increases plasma membrane fatty acid transport proteins in rat and human skeletal muscle[J].Am J Physiol Endocrinol Metab,2012,302(2):E183-189.
[12]CHENEVIERE X,BORRANI F,SANGSUE D,et al.Gender differences in whole-body fat oxidation kinetics during exercise[J].Appl Physiol Nutr Metab,2011,36(1):88-95.
[13]CHENEVIERE X,MALATESTA D,GOJANOVIC B,et al.Differences in whole-body fat oxidation kinetics between cycling and running[J].Eur J Appl Physiol,2010,109(6):1037-1045.
[14]CHENEVIERE X,MALATESTA D,PETERS E M,et al.A mathematical model to describe fat oxidation kinetics during graded exercise[J].Med Sci Sports Exe,2009,41(8):1615-1625.
[15]CRISP N A,FOURNIER P A,LICARI M K,et al.Adding sprints to continuous exercise at the intensity that maximises fat oxidation:implications for acute energy balance and enjoyment[J].Metabolism,2012,61(9):1280-1288.
[16]CRISP N A,GUELFI K J,LICARI M K,et al.Does exercise duration affect Fatmax in overweight boys?[J].Eur J Appl Physiol,2012,112(7):2557-2564.
[17]DASILVA S G,GUIDETTI L,BUZZACHERA C F,et al.Gender-based differences in substrate use during exercise at a self-selected pace[J].J Strength Cond Res,2011,25(9):2544-2551.
[18]DE GLISEZINSKI I,CRAMPES F,HARANT I,et al.Endurance training changes in lipolytic responsiveness of obese adipose tissue[J].Am J Physiol,1998,275(6Pt1):E951-956.
[19]FRAYN KN.Fat as a fuel:emerging understanding of the adipose tissue-skeletal muscle axis[J].Acta Physiol(Oxf),2010,199(4):509-518.
[20]HABETS D D,COUMANS W A,VOSHOL P J,et al.AMPK-mediated increase in myocardial long-chain fatty acid uptake critically depends on sarcolemmal CD36[J].Biochem Biophys Res Commun,2007,355(1):204-210.
[21]HAUFE S,ENGELI S,BUDZIAREK P,et al.Determinants of exercise-induced fat oxidation in obese women and men[J].Horm Metab Res,2010,42(3):215-221.
[22]HOLLOWAY G P,BEZAIRE V,HEIGENHAUSER G J,et al.Mitochondrial long chain fatty acid oxidation,fatty acid translocase/CD36content and carnitine palmitoyltransferase I activity in human skeletal muscle during aerobic exercise[J].J Physiol,2006,571(Pt1):201-210.
[23]JEUKENDRUP A E,ACHTEN J.Fatmax:A new concept to optimize Fat oxidation during exercise?[J].Eur J Sport Sci,2001,1(5):1-5.
[24]KIENS B,ALSTED T J,JEPPESEN J.Factors regulating fat oxidation in human skeletal muscle[J].Obes Rev,2011,12(10):852-858.
[25]KNECHTLE B,MULLER G,WILLMANN F,et al.Fat oxidation in men and women endurance athletes in running and cycling[J].Int J Sports Med,2004,25(1):38-44.
[26]KOPPO K,LARROUY D,MARQUES M A,et al.Lipid mobilization in subcutaneous adipose tissue during exercise in lean and obese humans.Roles of insulin and natriuretic peptides[J].Am J Physiol Endocrinol Metab,2010,299(2):E258-265.
[27]LAFONTAN M,MORO C,BERLAN M,et al.Control of lipol-ysis by natriuretic peptides and cyclic GMP[J].Trends Endocrinol Metab,2008,19(4):130-137.
[28]LIU C,WU J,ZhU J,et al.Lactate inhibits lipolysis in fat cells through activation of an orphan G-protein-coupled receptor,GPR81[J].J Biol Chem,2009,284(5):2811-2822.
[29]LUIKEN J J,COORT S L,WILLEMS J,et al.Contraction-induced fatty acid translocase/CD36translocation in rat cardiac myocytes is mediated through AMP-activated protein kinase signaling[J].Diabetes,2003,52(7):1627-1634.
[30]MORO C,PILLARD F,DE GLISEZINSKI I,et al.Sex differences in lipolysis-regulating mechanisms in overweight subjects:effect of exercise intensity[J].Obesity,2007,15(9):2245-2255.
[31]MORO C,PILLARD F,DE GLISEZINSKI I,et al.Training enhances ANP lipid-mobilizing action in adipose tissue of overweight men[J].Med Sci Sports Exe,2005,37(7):1126-1132.
[32]MORO C,POLAK J,HEJNOVA J,et al.Atrial natriuretic peptide stimulates lipid mobilization during repeated bouts of endurance exercise[J].Am J Physiol Endocrinol Metab,2006,290(5):E864-869.
[33]NIELSEN T S,VENDELBO M H,JESSEN N,et al.Fasting,but not exercise,increases adipose triglyceride lipase(ATGL)protein and reduces G(0)/G(1)switch gene 2(G0S2)protein and mRNA content in human adipose tissue[J].J Clin Endocrinol Metab,2011,96(8):E1293-1297.
[34]NORDBY P,SALTIN B,HELGE J W.Whole-body fat oxidation determined by graded exercise and indirect calorimetry:a role for muscle oxidative capacity?[J].Scand J Med Sci Sports,2006,16(3):209-214.
[35]PELSERS M M,STELLINGWERFF T,VAN LOON L J.The role of membrane fatty-acid transporters in regulating skeletal muscle substrate use during exercise[J].Sports Med,2008,38(5):387-399.
[36]PEREZ-MATUTE P,NEVILLE M J,TAN G D,et al.Transcriptional control of human adipose tissue blood flow[J].Obesity,2009,17(4):681-688.
[37]PERONNET F,MASSICOTTE D.Table of nonprotein respiratory quotient:an update[J].Can J Sport Sci,1991,16(1):23-29.
[38]PETRIDOU A,TSALOUHIDOU S,TSALIS G,et al.Longterm exercise increases the DNA binding activity of peroxisome proliferator-activated receptor gamma in rat adipose tissue[J].Metabolism,2007,56(8):1029-1036.
[39]RIDDELL M C,JAMNIK V K,ISCOE K E,et al.Fat oxidation rate and the exercise intensity that elicits maximal fat oxidation decreases with pubertal status in young male subjects[J].J Appl Physiol,2008,105(2):742-748.
[40]ROMIJN J A,COYLE E F,SIDOSSIS L S,et al.Substrate metabolism during different exercise intensities in endurancetrained women[J].J Appl Physiol,2000,88(5):1707-1714.
[41]ROONEY K,TRAYHURN P.Lactate and the GPR81receptor in metabolic regulation:implications for adipose tissue function and fatty acid utilisation by muscle during exercise[J].Br J Nutr,2011,106(9):1310-1316.
[42]SAMRA J S,SIMPSON E J,CLARK M Let al.Effects of epinephrine infusion on adipose tissue:interactions between blood flow and lipid metabolism[J].Am J Physiol,1996,271:E834-E839.
[43]SCHARHAG-ROSENBERGER F,MEYER T,WALITZEK S,et al.Effects of one year aerobic endurance training on resting metabolic rate and exercise fat oxidation in previously untrained men and women.Metabolic endurance training adaptations[J].Int J Sports Med,2010,31(7):498-504.
[44]SCHENK S,HOROWITZ J F.Coimmunoprecipitation of FAT/CD36and CPT I in skeletal muscle increases proportionally with fat oxidation after endurance exercise training[J].Am J Physiol Endocrinol Metab,2006,291(2):E254-E260.
[45]SCHOISWOHL G,SCHWEIGER M,SCHREIBER R,et al.Adipose triglyceride lipase plays a key role in the supply of the working muscle with fatty acids[J].J Lipid Res,2010,51(3):490-499.
[46]STARRITT E C,HOWLETT R A,HEIGENHAUSER G J,et al.Sensitivity of CPT I to malonyl-CoA in trained and untrained human skeletal muscle[J].Am J Physiol Endocrinol Metab,2000,278(3):E462-468.
[47]STICH V,DE GLISEZINSKI I,GALITZKY J,et al.Endurance training increases the beta-adrenergic lipolytic response in subcutaneous adipose tissue in obese subjects[J].Int J Obes,1999,23(4):374-381.
[48]STISEN A B,STOUGAARD O,LANGFORT J,et al.Maximal fat oxidation rates in endurance trained and untrained women[J].Eur J Appl Physiol,2006,98(5):497-506.
[49]TALANIAN J L,GALLOWAY S D,HEIGENHAUSER G J,et al.Two weeks of high-intensity aerobic interval training increases the capacity for fat oxidation during exercise in women[J].J Appl Physiol,2007,102(4):1439-1447.
[50]VENABLES M C,ACHTEN J,JEUKENDRUP A E.Determinants of fat oxidation during exercise in healthy men and women:a cross-sectional study[J].J Appl Physiol,2005,98(1):160-167.
[51]ZAKRZEWSKI J K,TOLFREY K.Acute effect of Fatmax exercise on the metabolism in overweight and nonoverweight girls[J].Med Sci Sports Exe,2012,44(9):1698-1705.
[52]ZAKRZEWSKI J K,TOLFREY K.Exercise protocols to estimate Fatmax and maximal fat oxidation in children[J].Pediatr Exerc Sci,2011,23(1):122-135.
[53]ZECHNER R,KIENESBERGER P C,HAEMMERLE G,et al.Adipose triglyceride lipase and the lipolytic catabolism of cellular fat stores[J].J Lipid Res,2009,50(1):3-21.
