变温层积过程中圆齿野鸦椿种子内含抑制物的生理活性变化
2013-04-10游双红涂淑萍
游双红,钟 诚,涂淑萍
(江西农业大学 林学院,江西 南昌 330045)
圆齿野鸦椿Euscaphis konishii属省沽油科Staphyleaceae 野鸦椿属Euscaphis常绿小乔木,是我国特有树种[1]。树高3~8 m,春夏之际(4~5月),黄白色小花集中生于枝顶,满树银花,十分美观;秋、冬、春三季(10月至翌年4月初),果实布满枝头,成熟后果荚裂开,果皮反卷,露出鲜红色的内果皮,黑色的种子粘挂在内果皮上,犹如满树红花上点缀着颗颗黑珍珠,非常艳丽,令人赏心悦目,是优良的观果树种。可孤植、列植或群植于草坪或疏林内,也可用于庭院、公园等的布景,或作矮化盆栽,室内观果[2]。目前,国外尚未见有关圆齿野鸦椿的研究报道,国内学者对圆齿野鸦椿的研究主要集中在树种的生物学特性及繁育技术方面[3-11],而较少见到有关其种子休眠方面的研究报道。引起种子休眠的原因主要有三个方面,即种皮、种胚和抑制物[12]。覃佳佳等人[13]对圆齿野鸦椿种子内含物的提取、分离以及生物测定进行了研究,结果表明,圆齿野鸦椿种子外种皮、内种皮及胚中抑制萌发物质的存在是引起其休眠的原因之一。本文主要研究在变温层积过程中圆齿野鸦椿种子内含抑制物的生理活性变化情况,旨在为打破圆齿野鸦椿种子休眠、促进种子萌发提供科学依据。
1 材料与方法
1.1 试验材料
圆齿野鸦椿种子购自福建省建瓯县万木林省级自然保护区。2011年1月,将种子与湿沙按1∶3的体积比混合均匀后再进行 9 个月的变温层积处理:先置于冰箱中于4 ℃下冷温层积处理3 个月(即冷温层积阶段Ⅰ);然后转入恒温箱中于25 ℃下暖温层积 3 个月,再置于冰箱中于4 ℃下冷温层积3 个月(即冷温层积阶段Ⅱ)。在各层积处理阶段的前、后分别取适量种子用于抑制物生理活性的测定。在变温层积过程中,定期洒水以保证沙的湿润度。
发芽试验采用白菜Brassica campestris种子,购自南昌市赣农蔬菜种子经营部,种子纯度≥95.00%,种子净度≥98.00%,发芽率≥85.00%,水分含量≤7.00%。使用前先用蒸馏水浸泡1 h。
1.2 试验方法
1.2.1 种仁、种壳的甲醇和蒸馏水浸提液的制备
分别以甲醇和蒸馏水为浸提剂制备圆齿野鸦椿种仁、种壳浸提液[14-16]。具体方法是:准确称取种仁或种壳5.00 g置于研钵中,加入少许甲醇,冰浴研磨成浆,以甲醇定溶至50 mL,转移至100 mL的锥形瓶中,于4 ℃下浸提24 h后,上离心机以500 r·min-1的转速离心15 min,取上清液,重复3 次,然后合并全部浸提液,用旋转蒸发仪于56 ℃下将甲醇挥发至干,加入25 mL的蒸馏水溶解残渣,得到质量浓度为0.20 g·mL-1的甲醇浸提液,取适量的该溶液加蒸馏水分别稀释成质量浓度为0.04、0.08、0.12、0.16 g·mL-1的圆齿野鸦椿种仁和种壳甲醇浸提液,与质量浓度为0.20 g·mL-1的原液一起置于4 ℃下保存以备用。
蒸馏水浸提液的制备方法同上,只要将甲醇替换成蒸馏水即可。
1.2.2 各种浸提液抑制活性的测定
用上述制备好的各种浸提液分别浸泡白菜种子24 h,以蒸馏水作为对照,每个处理30 粒种子。在培养皿中放置滤纸,分别加入各质量浓度的浸提液3.00 mL,然后将浸泡处理后的 30 粒白菜种子置于培养皿中,在 25 ℃的恒温箱中进行发芽,72 h后统计各处理的发芽率及幼苗生长状况(以胚轴弯曲、胚根伸长≥4.0 mm 作为萌发标准),每个处理重复4 次,并按下列公式计算相对发芽率、相对茎长和相对根长:
相对发芽率=(各处理的发芽率/对照处理发芽率)×100%;
相对茎长=(各处理的茎长/对照处理茎长)×100%;
相对根长=(各处理的根长/对照处理根长)×100%。
1.3 数据统计分析
采用Excel2003 和 SPSS17.0软件进行数据的统计与分析。
2 结果与分析
2.1 种子层积处理前浸提液抑制物的生理活性
2.1.1 圆齿野鸦椿种子浸提液对白菜种子发芽的影响
圆齿野鸦椿种仁和种壳的甲醇和蒸馏水浸提液对白菜种子发芽的影响情况如图1。从图1中可以看出,随着浸提液质量浓度的增加,其抑制作用显著增强;当质量浓度为0.20 g·mL-1时,以种仁浸提液处理的白菜种子其相对发芽率极显著低于种壳浸提液处理;种仁甲醇浸提液处理的白菜种子其相对发芽率为23.33%,显著低于种仁蒸馏水浸提液处理的白菜种子的相对发芽率(28.33%);而种壳甲醇浸提液处理的白菜种子其相对发芽率为58.33%,与种壳蒸馏水浸提液处理的白菜种子相对发芽率(61.67%)的差异不显著。这说明圆齿野鸦椿种仁和种壳中均含有发芽抑制物,但种仁的抑制活性极显著高于种壳;种仁中的发芽抑制物以有机相为主,同时也含有一定量的水溶性成分,而种壳中的有机相与水溶性成分二者抑制活性间的差异并不显著。
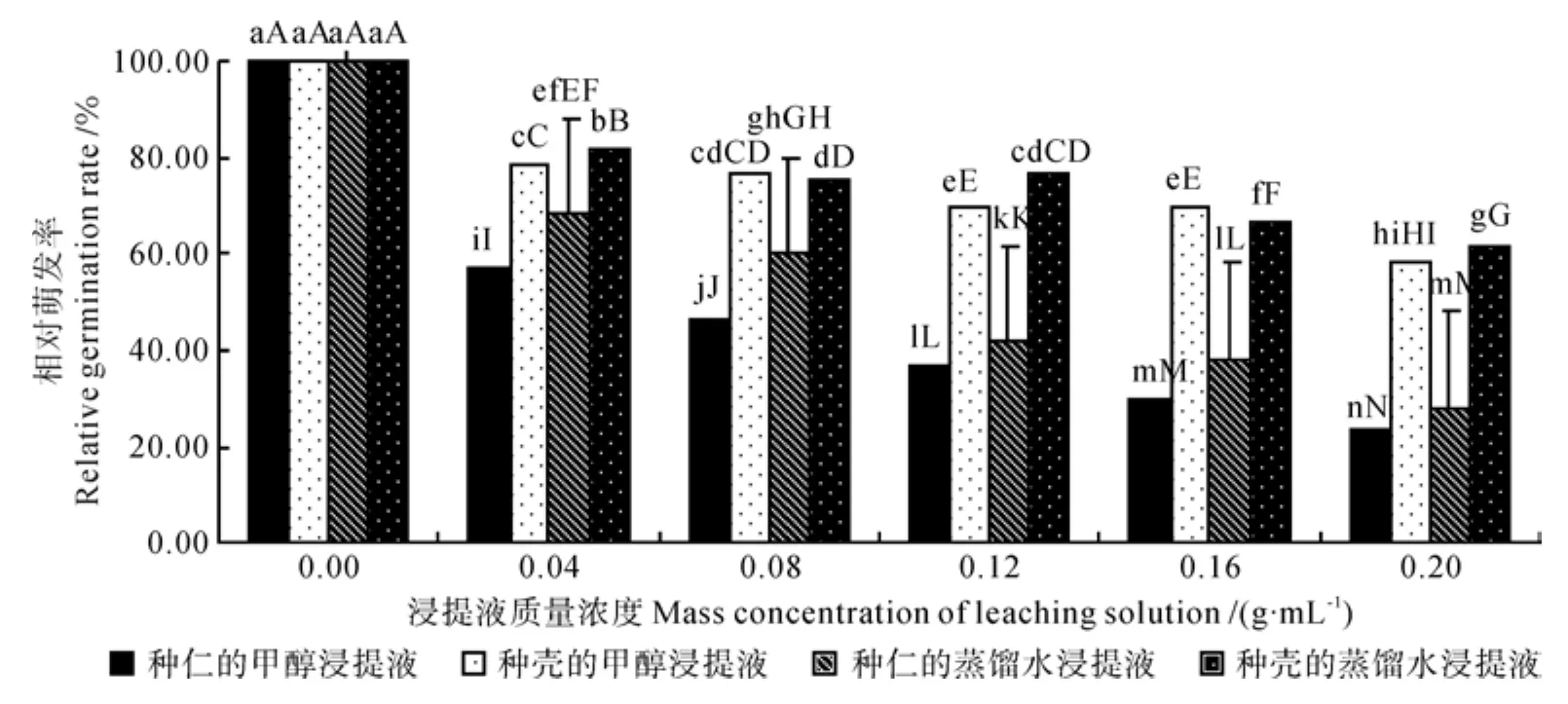
图1 不同质量浓度浸提液处理的白菜种子的相对萌发率Fig.1 Relative germination rate of Brassica campestris seed treated by different mass concentrations of leaching solution
2.1.2 种子浸提液对白菜幼苗生长的影响
圆齿野鸦椿种仁和种壳的甲醇和蒸馏水浸提液对白菜幼苗生长的影响情况如表1。由表1可知,种仁和种壳的甲醇及蒸馏水浸提液对白菜幼苗的根及茎生长均有极显著的抑制作用,且随着浸提液质量浓度的增加,其抑制作用显著增强。以0.20 g·mL-1的种仁甲醇浸提液处理的白菜幼苗其根最短,说明该浸提液对白菜幼苗根长生长的抑制作用最强。将以0.20 g·mL-1的种仁甲醇浸提液与以相同浓度的种仁蒸馏水浸提液及种壳甲醇浸提液处理的相比,幼苗根长的差异均达显著水平,与种壳蒸馏水浸提液处理的相比,幼苗根长的差异达极显著水平;种壳的甲醇浸提液处理与种壳的蒸馏水浸提液处理相比,幼苗根长的差异亦达显著水平。以0.20 g·mL-1的种仁甲醇浸提液处理白菜幼苗的茎最短,说明该浸提液对白菜幼苗茎长生长的抑制作用最强。它与相同浓度的其他三个浸提液处理相比,幼苗茎长的差异均达极显著水平;种壳的甲醇浸提液处理与种仁的蒸馏水浸提液处理相比,幼苗茎长的差异达显著水平,与种壳的蒸馏水浸提液处理相比,幼苗茎长的差异达极显著水平;种仁的蒸馏水浸提液处理与种壳的蒸馏水浸提液相比,幼苗茎长的差异亦达极显著水平。由此可见,对白菜幼苗生长的抑制作用,甲醇浸提液处理显著大于蒸馏水浸提液处理,种仁浸提液处理显著大于种壳浸提液处理。各处理对白菜幼苗茎生长的抑制作用大于对根生长的抑制作用。
2.2 变温层积过程中种子浸提液抑制活性的变化
2.2.1 变温层积过程中种子浸提液对白菜种子发芽的影响
变温层积过程中圆齿野鸦椿种子浸提液对白菜种子发芽率的影响情况如表2所示。从表2中可以看出,以0.20 g·mL-1种仁甲醇浸提液处理的白菜种子其相对发芽率,层积前仅为23.33%,经3个月的4 ℃冷温层积阶段Ⅰ后达28.34%,再经3个月的25 ℃暖温层积处理后的为46.67%,差异均达极显著水平;又经3个月的4 ℃冷温层积阶段Ⅱ后,白菜种子的相对发芽率仍然保持46.67%。但当浸提液质量浓度下降至0.12 g·mL-1及其以下时,圆齿野鸦椿种子在变温层积处理的各阶段其发芽抑制物活性逐渐降低,差异达极显著水平。种子经过9 个月的变温层积处理后,以0.20 g·mL-1的种仁甲醇浸提液处理的白菜种子其相对发芽率为46.67%,白菜幼苗的相对茎长为34.27%、相对根长为41.71%。试验结果与实际情况相符,即该种子于采种后的第2年秋季播种,发芽率仍然很低,说明圆齿野鸦椿种子具有深度休眠的特性。
2.2.2 变温层积过程中种子浸提液对白菜幼苗生长的影响
变温层积过程中圆齿野鸦椿种子浸提液对白菜幼苗茎长、根长的影响情况如表3,圆齿野鸦椿种仁甲醇浸提液处理后白菜幼苗生长量的多重比较结果如表4。
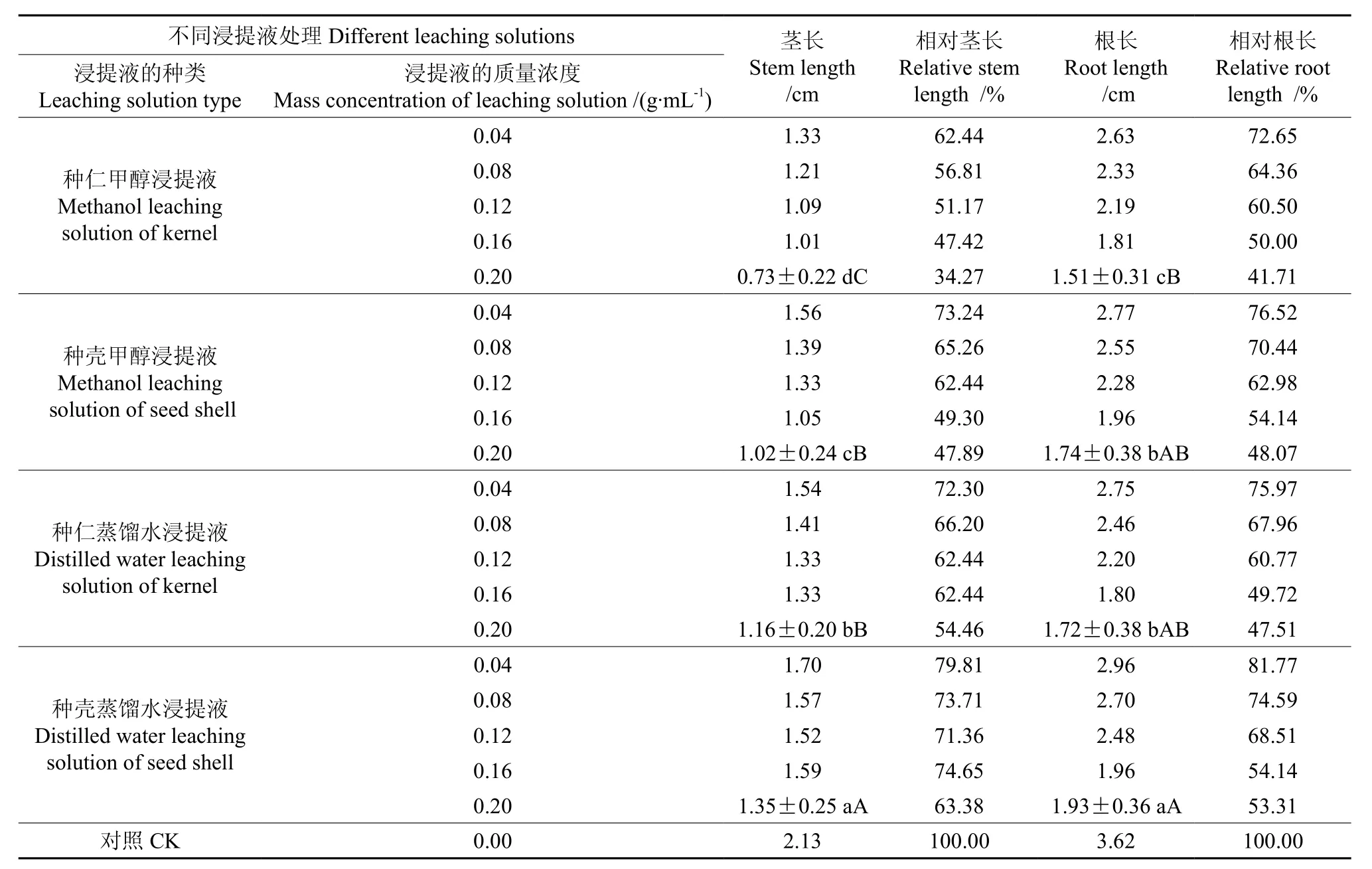
表1 各种浸提液对白菜幼苗生长的影响†Table 1 Effect of different leaching solutions on growth of Brassica campestris seedling
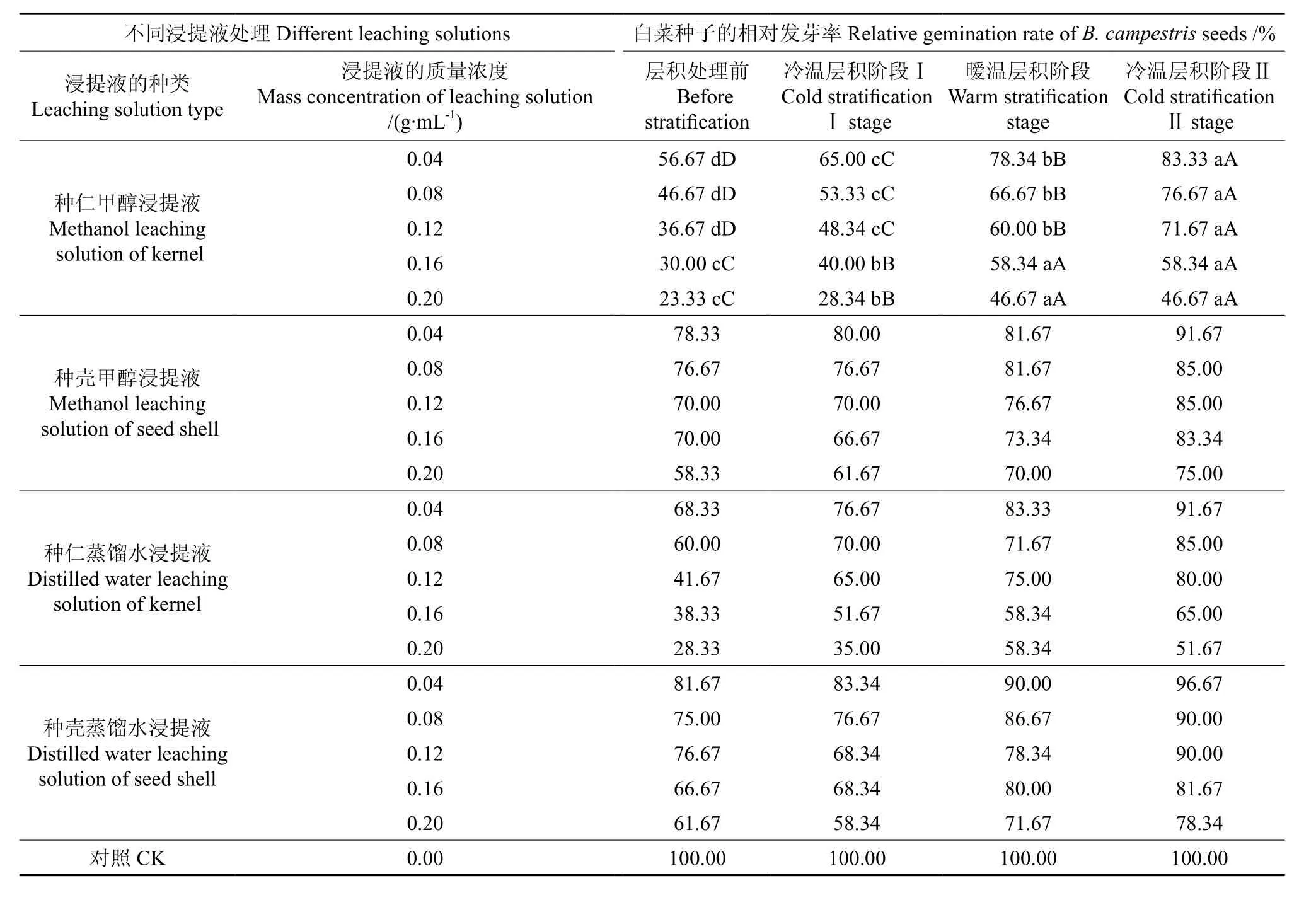
表2 变温层积各阶段种子浸提液对白菜种子发芽率的影响Table 2 Effect of seed leaching solution at each stage of variable temperature stratification on germination rate of Brassica campestris seeds
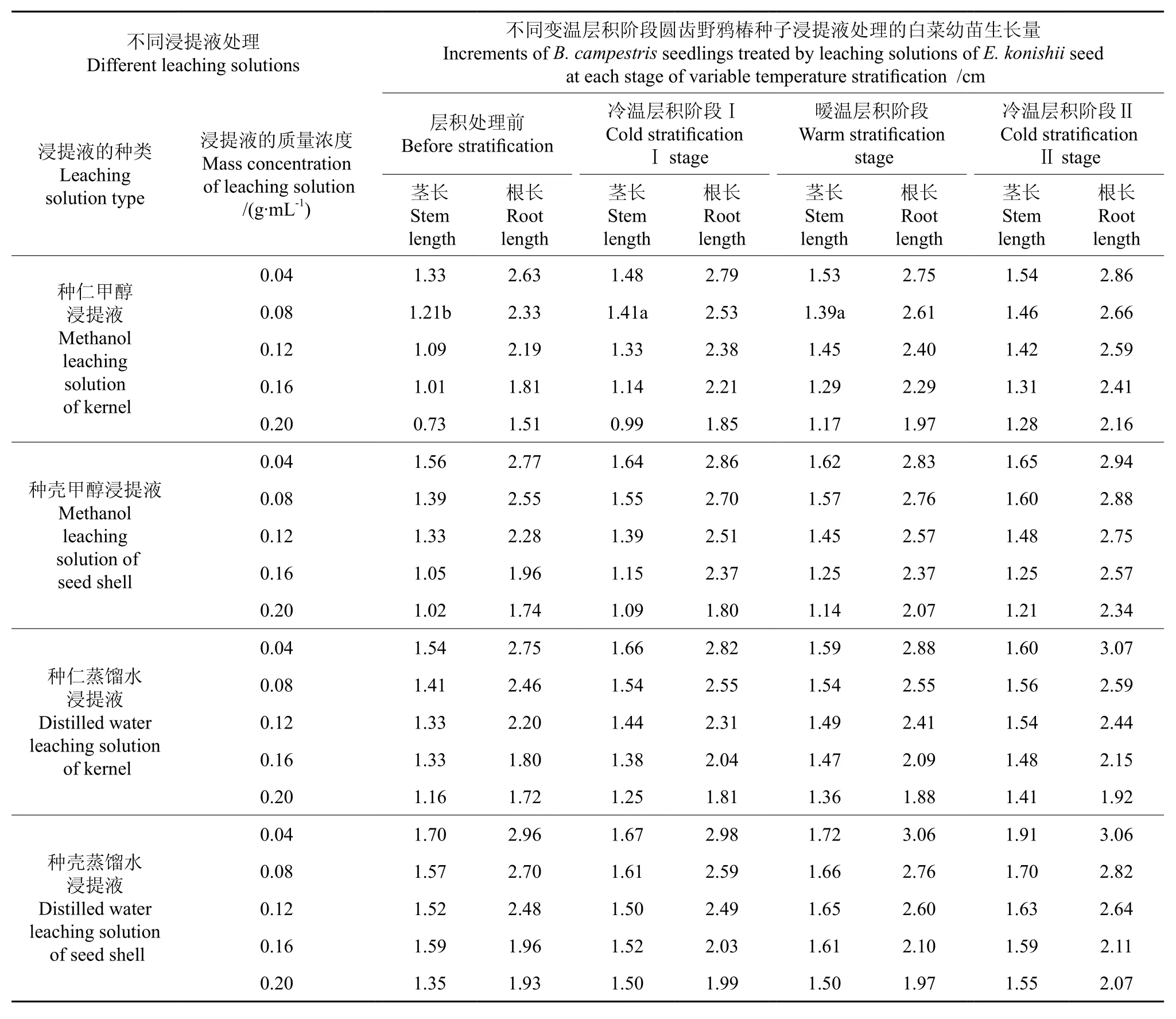
表3 变温层积圆齿野鸦椿种子浸提物对白菜幼苗生长的影响Table 3 Effect of leaching solutions of Euscaphis konishii seeds at each stage of variable temperature stratification on growth of Brassica campestris seedlings
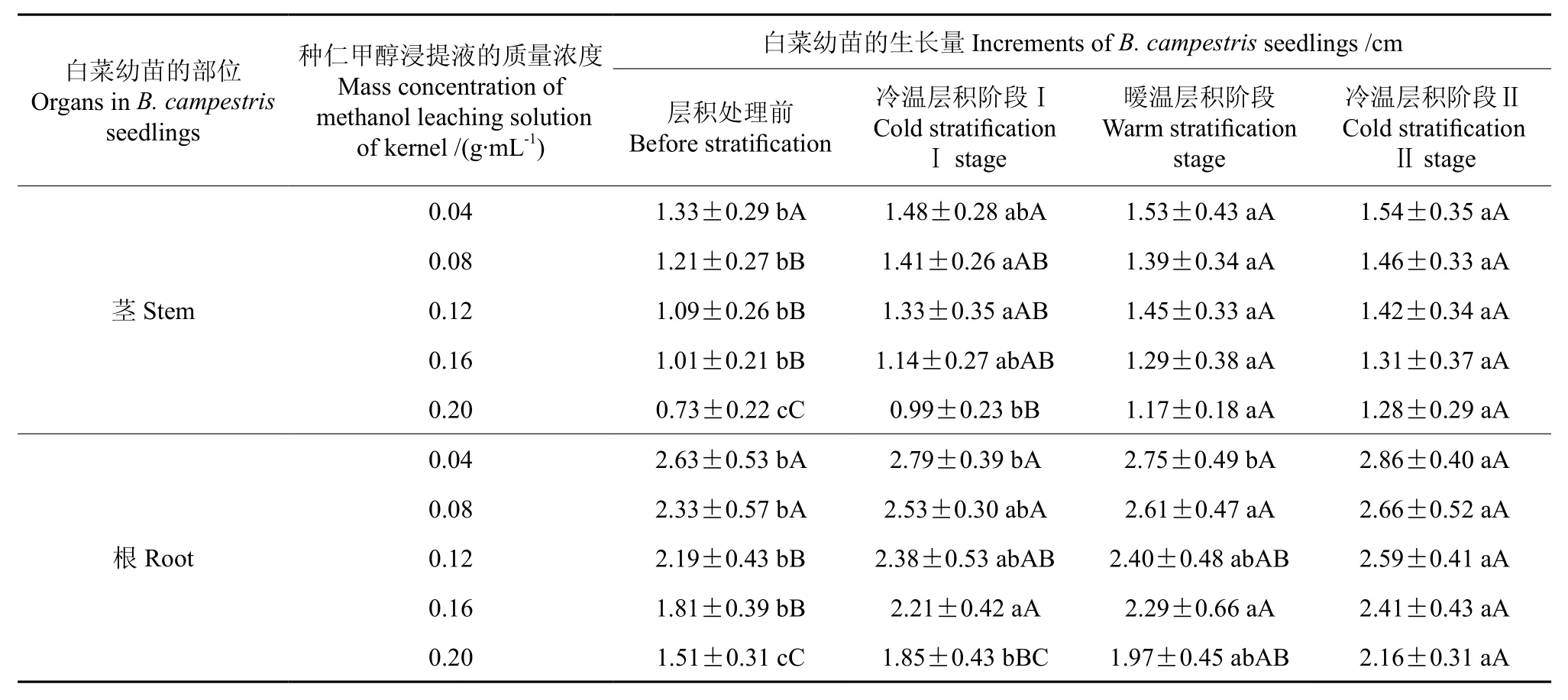
表4 不同质量浓度的种仁甲醇浸提液处理后白菜幼苗生长量的多重比较结果Table 4 Result of multiple comparison of increments of B. campestris seedlings treated by different mass concentrations of methanol leaching solutions of kernel
从表3中可以看出,随着变温层积处理的进行,圆齿野鸦椿种子的抑制活性逐渐降低,表现为白菜种子的相对发芽率逐渐提高,白菜幼苗的茎长与根长逐渐增加。
从表4中可以看出,以0.20 g·mL-1圆齿野鸦椿种仁甲醇浸提液处理的白菜幼苗的茎长,层积前为0.73 cm,经3 个月的4 ℃冷温层积处理Ⅰ后为0.99 cm,再经3 个月25 ℃的暖温层积(变温层积6个月)后为1.17 cm,以上各阶段幼苗茎长的差异达极显著水平,再经3 个月的冷温层积处理Ⅱ(变温层积9个月)后幼苗茎长为1.28 cm,与经暖温层积(变温层积6个月)后的相比,二者差异不显著。白菜幼苗的根长,层积前为1.51 cm,与经3 个月的4 ℃冷温层积处理Ⅰ后的根长(1.85 cm)相比,其差异达显著水平;与再经3 个月的25 ℃暖温层积(变温层积6 个月)后的根长(1.97 cm)及再经3 个月的4 ℃冷温层积处理Ⅱ(变温层积9 个月)后的根长(2.16 cm)相比,二者差异均达极显著水平,但将变温层积6 个月与变温层积9 个月的种子浸提液处理的白菜根长相比,二者间的差异不显著。这与杨晓玲等人[17]研究的山楂在层积过程中抑制物的变化结果基本一致。
3 结论与讨论
3.1 抑制物在种子中的分布位置
自然界中有许多植物的种子中存在发芽抑制物,但抑制物在种子中的分布位置有所不同。如西洋参主要分布于果皮和果肉中[18],青钱柳主要分布于外种皮上[19],日本女贞主要分布于胚乳中,向日葵[20]和三角槭[21]分布于种胚中,珊瑚树[22]分布于果皮和种胚中,而天女木兰种子的果皮、假种皮、中种皮和胚乳中均有抑制物质存在[23]。本试验结果表明,圆齿野鸦椿种子的种仁和种壳中均有发芽抑制物和生长抑制物存在,但种仁的抑制活性极显著高于种壳。
3.2 抑制物以有机相的极性化合物为主,亦有一定量的水溶性成分
试验结果表明,圆齿野鸦椿种子所含抑制物以有机相的极性化合物为主,亦含有一定量的水溶性成分,这与四川牡丹等植物种子类似[24-26]。因此,热水浸种可以提高种子的发芽率,这可从欧斌等人的研究结果[5](即用65~68 ℃的热水浸泡圆齿野鸦椿种子,可以提高种子发芽率)中得到证实,此外,李彩琴等人[27]的研究结果也表明,以45 ℃温水短时间浸提宽叶羌种子,可有效除去大部分内源抑制物。
3.3 圆齿野鸦椿种子具有深度休眠特性
圆齿野鸦椿种子经变温层积处理9 个月后,以0.20 g·mL-1的种仁甲醇浸提液处理的白菜种子其相对发芽率仅为46.67%,相对茎长为34.27%,相对根长为41.71%。这一试验结果表明,此时种子内仍含有一定量的发芽抑物和生长抑制物,即圆齿野鸦椿种子具有深休眠特性,短时间内难于解除休眠。生产实践证明,圆齿野鸦椿种子层积处理时间需达一年以上方可解除休眠。
3.4 种子内含发芽抑制物是圆齿野鸦椿种子休眠的主要原因之一
层积处理是解除圆齿野鸦椿种子休眠的主要措施,而变温层积处理可通过温度的变化,降低萌发抑制物的生理活性[28-31],加快种胚的后熟,从而促进种子萌发。在本试验中,种子在层积处理9 个月后,种胚已经发育成熟,但仍存在一定的抑制活性,播种后发芽率仍很低,这说明种子内含发芽抑制物是圆齿野鸦椿种子休眠的主要原因之一。
参考文献:
[1]欧 斌.圆齿野鸦椿(Euscaphis konishiiHayata)[J].江西林业科技,2006(5):61.
[2]曹人智.观果新品种——圆齿野鸦椿[J].林业实用技术,2006,(8):46.
[3]覃嘉佳,聂海兵,龙云英,等.圆齿野鸦椿种子发芽过程及其酶活性变化的研究[J].江西林业科技,2007,(6):7-9.
[4]覃嘉佳,龙云英.圆齿野鸦椿扦插繁殖技术[J].林业科技开发,2007,21(3):71-73.
[5]欧 斌,李远章.圆齿野鸦椿种子预处理和苗木生长规律及育苗技术研究[J].江西林业科技,2006(3):16-18.
[6]曹 蕾,涂淑萍.圆齿野鸦椿愈伤组织诱导研究[G].国观赏园艺研究进展.北京:中国林业出版社,2008:268-270.
[7]涂淑萍,曹 蕾.圆齿野鸦椿芽继代增殖的影响因素[J].安徽农业科学,2009,37(28):13486-13487,13413.
[8]杨燕凌.打破圆齿野鸦椿种子休眠及外植体选择诱导实验研究[D],福建:福建农林大学,2008:24-31.
[9]支丽燕,吴田兵,龙云英,等.干旱胁迫下圆齿野鸦椿苗期叶片的生理特性[J],福建林学院学报,2008,28(2):190-192.
[10]支丽燕,胡松竹,余 林,等.涝渍胁迫对圆齿野鸦椿苗期生长及其叶片生理的影响[J],江西农业大学学报,2008,30(2):279- 282.
[11]许方宏,张倩媚,王 俊,等.圆齿野鸦椿的生态生物学特性,生态环境学报,2009,18(1):306-309.
[12]黄 丹,许岳香,胡海波.植物种子休眠原因与机理的研究进展[J].亚热带植物科学,2010,39(2):78-83.
[13]覃嘉佳,胡 滨,黄焱辉.圆齿野鸦椿种子内含物的提取·分离以及生物测定[J].安徽农业科学,2011,39(32):19693-19694,19697.
[14]赵 敏.防风种子中内源抑制物质活性的研究[J].中草药,2004, 35(4):441-444.
[15]张艳杰,高捍东,鲁顺保.南方红豆杉种子中发芽抑制物的研究[J].南京林业大学学报(自然科学版),2007,31(4):51-56.
[16]罗夫来,郭巧生.百蕊草种子内源抑制物质的初步研究[J].中国中药杂志,2007,32(17):1737-1 739.
[17]杨晓玲,郭守华,关学敏.山楂种子层积过程中抑制物质的变化[J].经济林研究,2009,27(2):44-48.
[18]韩 东,黄耀阁,李向高,等.西洋参果实中发芽抑制物质——二苯胺的分离鉴定[J].吉林农业大学学报, 2001,23(4): 60- 63,68.
[19]杨万霞,方升佐.青钱柳种皮甲醇浸提液的生物测定[J].植物资源与环境学报,2005,14(4):l1-14.
[20]孔祥海.抑制物质和种子休眠[J].龙岩师专学报,2002, 20(6):50-52.
[21]肖至诚.三角槭种子休眠机理及解除方法的研究[D].南京:南京林业大学,2006:18.
[22]吴小娴,高捍东.珊瑚朴种子生物学特性研究[J].林业科技开发,2008,22(05):55-57.
[23]李 澎,陆秀君,姚 飞,等天女木兰种子休眠原因的初步探讨[J].种子,2006,25(3):36-39.
[24]宋会兴,刘立光,高素萍,等.四川牡丹种子浸提液内源抑制物活性初探[J].园艺学报,2012,39(02):370-374.
[25]胡曼玲,裴明玉.轮叶党参种子萌发抑制物质活性研究[J].人参研究,2005,(4):15-17.
[26]张秋菊,秦佳梅,徐克章,等.不同溶剂提取对返魂草种子中抑制物质活性的影响[J].植物生理学通讯, 2005, 41(5):629-630.
[27]李彩琴,陈 垣,郭凤霞,等.宽叶羌活种子浸提液对白菜种子萌发及幼苗生长的抑制活性[J].甘肃农业大学学报,2008,(5): 84-86.
[28]吴正军,朱再标,郭巧生.老鸦瓣种子生理及其萌发特性研究[J].中国中药杂志,2012,37(5):575-579.
[29]黄儒珠,郭详泉,方兴添.变温层积处理对南方红豆杉种子生理生化特性的研究[J].福建师范大学学报,2006,22(2):95-98.
[30]杨晓玲,郭守华,关学敏,等.山楂种子层积过程中抑制物的变化[J].经济林研究,2009,27(2):44-48.
[31]范文丽,宁 伟,李天来.变温层积条件下GA3对长白惚木种子萌发的影响[J].沈阳农业大学学报,2008,39(1):80-82.
