高原鼠兔洞系特征及功能研究
2013-04-10卫万荣张灵菲杨国荣徐吉伟范旭东张卫国
卫万荣,张灵菲,杨国荣,徐吉伟,范旭东,张卫国
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730020)
高原鼠兔(Ochotonacurzoniae)的活动空间包括地上和地下2个部分,洞口既是这2个特定空间的界面,也是使之得以贯通的门户。洞口是洞系乃至领地的重要指征,因而,洞口的基本特征(如数量、位置、朝向、格局等)与高原鼠兔的生境适合度,特别是洞道建筑特点存在必然的逻辑关系,隐示着高原鼠兔对栖居环境及洞道建筑的一种适应性取向,包含着大量生态学和生物学信息[1-3]。基于自然制约力控制草原鼠类数量和分布的生态调控法,是当今草原鼠害防治发展的主流方向,也是鼠害防控研究的热点领域[4]。因此,将研究的视角聚焦于洞系特征与鼠类生境适合度和洞道建筑的关联性,并将其与高原鼠兔的生态防控实践相联系,不仅可深化对高原鼠兔生境选择取向的认识,而且也是补充和完善草原鼠害生态防控理论,实施生态防控的重要依据。
高原鼠兔是高寒草原生态系统中最主要的啮齿动物之一,对维持草原生态系统的平衡与稳定具有重要的不可替代的作用[5]。高原鼠兔同时也是高寒牧区草原畜牧业和生态环境最重要的危害因素之一,常可在过度放牧的诱导下对草原植被造成严重破坏[6]。因此,如何将高原鼠兔的种群数量控制在对生态环境有利、对牧业生产无害的水平上,始终是科研工作者和生产实践者努力探讨的一个重要命题[7]。种群数量的生态调控是近年来极具活力的热点领域,也是未来实施鼠害治理和生态保护的根本途径。实施生态调控的前提是全面掌握高原鼠兔的生态习性,特别是对生活环境的适应性取向。目前,对于高原鼠兔生态习性的研究已较为深入,特别是栖境适合度与植被性状的相关性研究尤显全面而精细[8-9],此外,栖境与天敌、栖境与气候等领域的研究也被给予了应有的关注[9-11]。然而,作为对栖境适合度具有较高指示意义的洞口特征的相关研究迄今仍极少有人涉及,现有的研究大多仅将其视为洞系结构的一个组成部分,至于其“所以然”的生态学意义则少有问津,而这一点正是对生态调控策略最具现实指导意义的环节。有鉴于此,本研究将在全面研究高原鼠兔洞口特征的基础上,着力探讨其与各主要环境因素间的相关关系,为高原鼠兔的生态防控实践提供理论依据。
1 材料与方法
1.1 研究地概况
研究地设于玛曲县阿孜畜牧实验站场部东南向约2km处的公路南侧约80hm2的区域,核心区的地理坐标为33°40′40″N,101°53′20″E,海拔3540m。地貌属山间河谷滩地,地势平坦开阔,土层50~70cm。年均温1.2℃,最高平均气温10.8℃,最低平均气温-9.4℃;年均降水量643.9mm,最高年份821.9mm,最低年份472.1mm;冬春季多西北风,1年中出现8级以上大风日平均在37d以上。植被为典型亚高山草地,长期作冷季草场使用,因近十余年来,放牧期延长、载牧量上升、气候转暖、地下水位下降等因素的综合影响,草地已发生严重退化。主要植被成分除了莎草科、禾本科的物种仍占有一定比重外,毛茛科的丝叶毛茛(Ranunculustanguticus)、金莲花(Trolliuschinensis)、草玉梅(Anemonerivularis),菊科的黄帚橐吾(Ligulariavirgaurea)、风毛菊(Saussureagossypiphora)、火绒草(Leontopodiumalpinum),蔷薇科的鹅绒委陵菜(Potentillaanserina)、莓叶委陵菜(Potentillafragarioidis),龙胆科的秦艽(Gentianamacrophylla),玄生科的马先蒿(Pedicularisresupinata),豆科的披针叶黄华(Thermopsislanceolata)等物种也占有较大比重[12-13]。至1990年代起,此地即为高原鼠兔典型分布地,2009年进入高密度周期,鼠群密度在330~780个有效洞口/hm2之间。
1.2 样地设置
在研究地分别选3个高原鼠兔的自然分布斑块设为样地,各样地内需保证至少有300个以上的高原鼠兔洞口。3个样地分别用于2011年5,8月和2012年1月的观测。
1.3 观测项目及方法
洞口朝向:分东(east,E)、南(south,S)、西(west,W)、北(north,N)、东南(southeast,ES)、东北(northeast,EN)、西南(southwest,WS)、西北(northwest,WN)8个方向。E向指正东偏S、偏N各15°的区域,S向指正南偏E、偏 W 各15°的区域,W、N向依次类推;ES向指E偏S 30°与S偏E 30°之间的区域,WN向指 W 偏N 30°与N偏W 30°的区域,EN、WS向依次类推(图1)。洞口朝向用罗盘仪逐一确定。无显著洞口朝向的洞口(no direction,ND)指洞口面与地平面基本处于同一平面的洞口。
洞口斜度:指洞口下缘与洞口上缘连线与水平线的角度,用坡度仪测定(图2)。
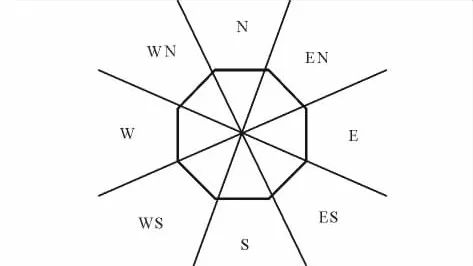
图1 洞口朝向的划定Fig.1 Demarcation of portal orientation
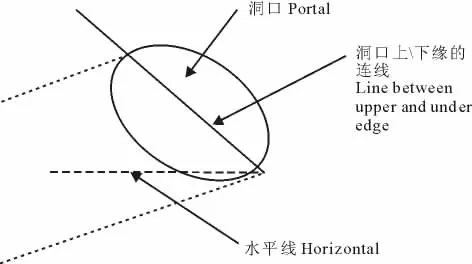
图2 洞口斜度示意Fig.2 Hint of portal slope
洞口温度:指洞口内10cm处的温度,用探针式电子温度计测定。测定时以1个洞系为单位,各朝向所有洞口于同一时刻进行(误差≤1min),以将探针置入洞口3 min后的数据为准。共测定10个洞系,在晴朗天气条件下于10:00-16:00时测定,取各朝向测定值的平均值用于比较分析。
洞道结构:于各样地分别解剖一个洞系,测定洞道直径、深度、拐点、窝巢等结构特征,并用计算纸现场绘制洞系结构图。
1.4 统计分析
采用Excel 2003版、SPSS 11.5版统计软件进行图表制作和统计分析。
2 结果与分析
2.1 洞口朝向的空间配置
3个样地共900个(3×300)洞口朝向的空间配置如图3、图4所示。总体而言,洞口朝向以正西(W)、东南(ES)、东北(EN)和西南(WS)4个方向较为集中,其洞口数依次为(42.3±4.3),(44.3±2.0),(45.7±3.0)和(42.3±1.5)个,分别占统计洞口总数的14.1%,14.8%,15.2%和14.1%,4个方向的洞口数无显著性差异(P>0.05);其次为正南(S)和西北(WN)2个方向,洞口数分别为(37.3±3.3)和(39.7±2.0)个,占统计洞口数的12.4%和13.2%,二者间无显著性差异(P>0.05),但均显著低于前4个方向(P<0.05);正东(E)、正北(N)方向的洞口数较少,分别为(27.7±6.5)和(13.7±2.5)个,占洞口数的9.2%和4.6%,与以上6个方向的洞口数差异均极显著(P<0.01),而且二者间也存在极显著差异(P<0.01);无明显洞口朝向(ND)的洞口数最低,平均为(7.0±1.0)个,仅占洞口数的2.3%,与8个方向的洞口数差异均极显著(P<0.01)。
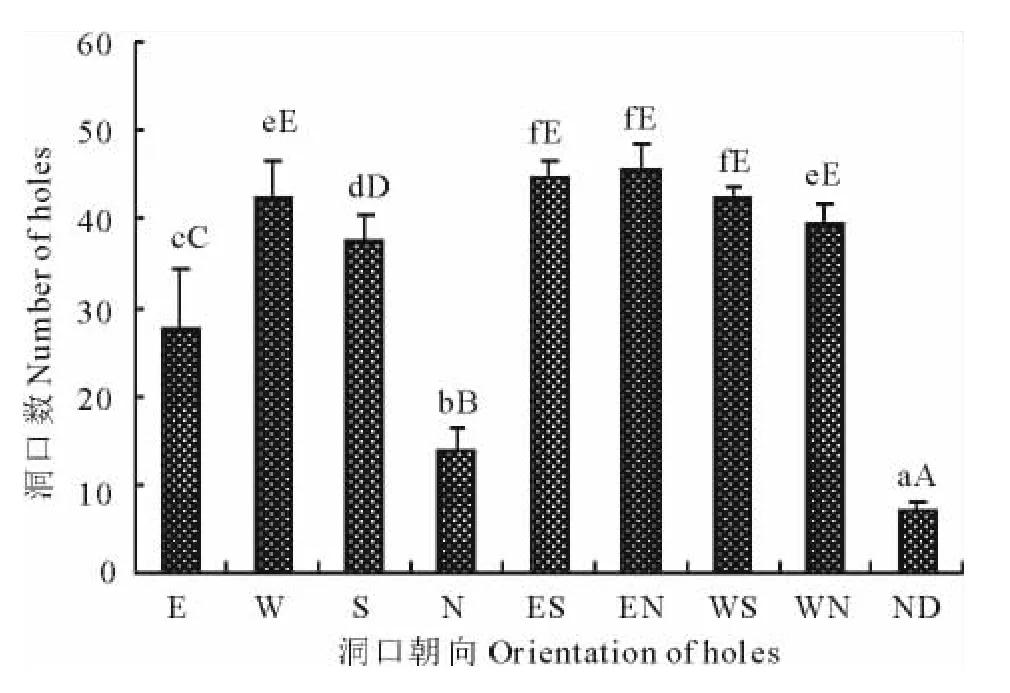
图3 3个样地洞口朝向配置格局Fig.3 Distribution pattern of portal orientation in 3sites
若将洞口朝向划分为全阴(N、WN、EN)、半阴(W、E)和全阳(S、WS、ES)3个类型,其洞口数平均依次为(99.1±10.8),(70.0±5.0)和(123.9±6.8)个,分别占洞口总数的33.0%,23.3%和41.3%,三者间均存在极显著差异(P<0.01)。
2.2 洞口直径与斜度
由图5可见,洞口直径和斜度决定着洞口面积,斜度越大表明其与外界因素(如光照、风等)的作用面积越大,反之则越小。洞口直径在8个方向上均未表现出显著性差异(表1,P>0.05),表明洞口直径与洞口朝向无相关性。洞口斜度≤55°的洞口主要集中在SW、SE和S 3个方向,占洞口总数的48.6%;在E、W方向的占23.4%;而在NW、N和NE三个方向洞口数仅为洞口总数的27.8%。N向和偏N向、正E和正W向斜度≤55°的洞口数极显著低于S向和偏S向(P<0.01),而N向和偏N向与正E和正 W向的洞口数无显著性差异(P>0.05)。洞口斜度>55°的洞口主要集中在NW、NE和N三个方向,占洞口总数的52.1%;E、W方向的洞口占23.6%;SW、SE和S方向的洞口占24.3%。E向、ES向、正 W和正S向斜度>55°的洞口数无显著性差异(P>0.05),而N向与EN、WN向与正 W向的洞口数间亦无显著性差异(P>0.05),但前4个方向与后4个方向间的差异极显著(P<0.01)(表1)。这一结果说明,洞口斜度的设置与太阳角的高度的季节差异密切相关,斜度≤55°的洞口数多,特别是其中S向和偏S向的洞口所占比例较大,意味着有利于冬季对太阳辐射热量的获取,提升洞内温度。
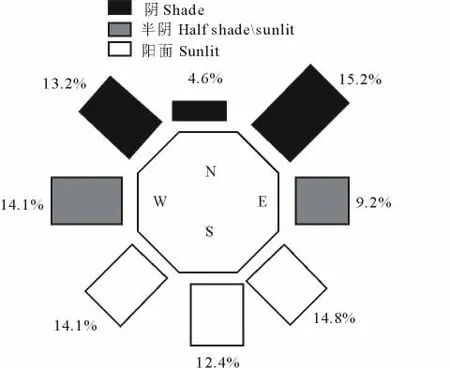
图4 洞口朝向的空间配置Fig.4 Spacial distribution of portal orientation
2.3 不同洞口朝向的温度差异
如表1所示,洞口温度因洞口朝向不同而存在明显差异。温度从高到低的洞口朝向排序是:S>WS>W>ES>ND>E>WN>N>EN,其中,最高温度(21.3℃)与最低温度(13.2℃)的差值达8.1℃(P<0.01),除N与EN和WN,E与ND以及ES与W间无显著差异(P>0.05)外,其余朝向间均存在显著(P<0.05)或极显著差异(P<0.01)。但总体上表现为S和偏S的洞口温度显著高于N和偏N的洞口,W和偏W洞口普遍高于E和偏E洞口的基本态势。
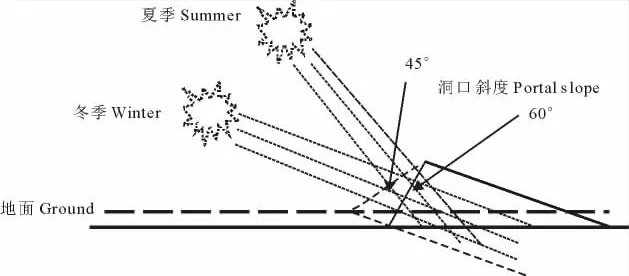
图5 洞口斜度与太阳入射角高度的关系Fig.5 Relationship between portal slope and sun altitude
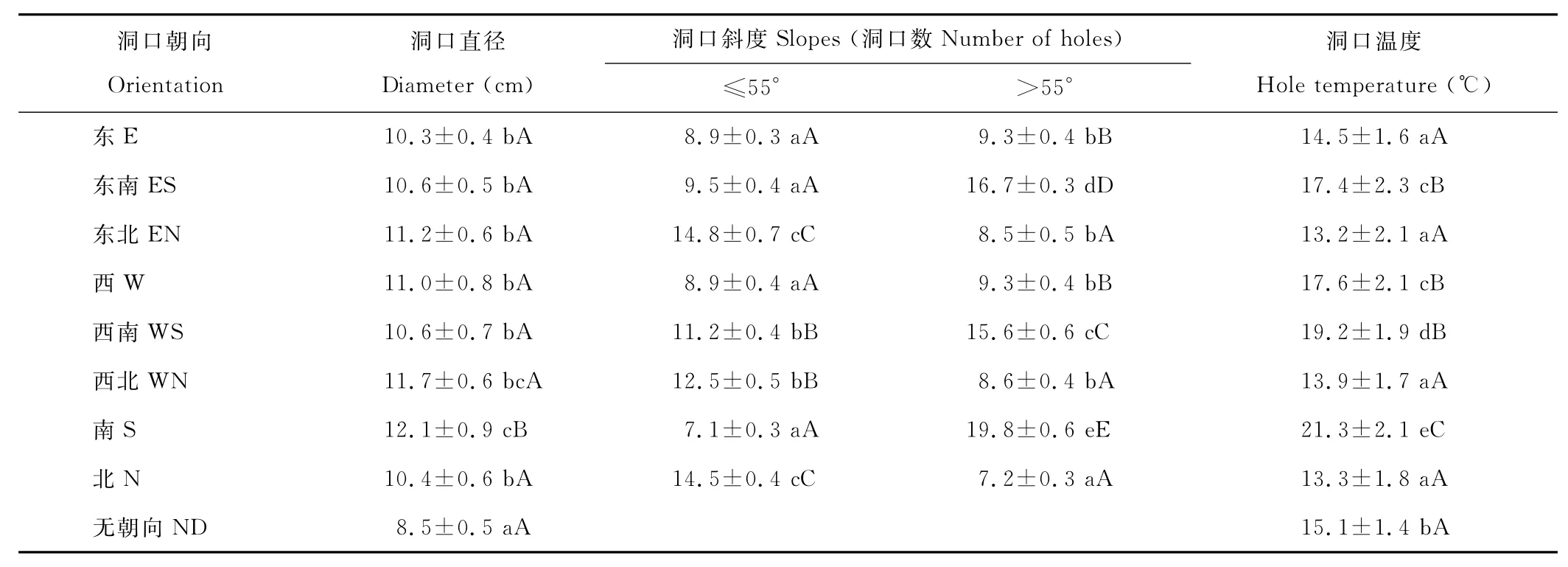
表1 不同洞口朝向下的洞口直径、斜度及温度差异Table 1 The diameter,slope and temperature of holes with different orientation
2.4 洞道结构
对3个独立洞系的解剖结果表明,开放洞口数最多为6个,最少为4个,平均4.7个。其中,朝向为N偏E的洞口数平均为2.3个,正S 1.0个,S偏 W 1.4个,EN向与S和 WS向的洞口数基本相等,分别为2.3和2.4个。3个洞系的洞道总长度平均为6.37m,洞道平均直径7.1cm,距地面最大距离47.2cm,3个洞系35个测点(拐点)距地面的平均距离为30.1cm。洞道以窝巢为中心呈放射状分布(图6),但通往N或偏N向的主干道平均为1.3条,而通往S或偏S向的主干道为2.3条,二者差异极显著(P<0.01)。在S或偏S向通道中,其中1条较为简单,距离相对较短,拐点少且弯曲度小,第1个拐点距洞口相对较远;而另外1条则较为复杂,距离明显较长,拐点多,弯曲度大,中段常可出现90°的转角,第1个拐点距洞口也相对较近。N或偏N向通道的显著特征是在主干道中段的拐点弯曲度较大,特别是洞口入口段向下延伸的坡度可达70°以上,在水平延伸40cm左右后猛然以接近直角的方式导入主干道。
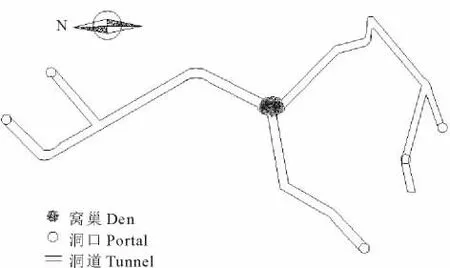
图6 洞系结构平面示意图Fig.6 The sketch ichnography of the tunnel structure of pika
3 讨论
草原鼠类主要营穴居生活,地下洞穴不仅为其提供了栖居、繁殖和活动的场所,同时也是其规避捕食和伤害风险、应对恶劣天气条件所依赖的基本设施。洞系结构、布局和规格除须满足上述功能的基本要求外,保持较高的舒适性也是洞系建造中必须兼顾的一个因素。对于高原鼠兔而言,由于其分布地具有冬季漫长(7个月以上)、寒冷多风、太阳辐射强的气候特点,因而,如何既能保持洞内的适宜温度又可满足通风换气的需要,便成了高原鼠兔在建造洞系时无法回避的问题。在有关高原鼠兔洞系构造现有的研究中,人们关注的视角主要集中于洞系本身的构造特征[14]及其在降低能耗[15]、防范捕食风险[16]和自然灾害[17]等方面的功能性对策,虽然也有极少数研究者注意到了保温与通风对洞系结构的特殊要求,但这类研究似乎只是对洞口朝向和入口处坡度与太阳角和风向的季节性变化给予了更多的关注[18-21],而对于不同朝向洞口的空间配置、温度差异、洞口斜度以及洞系内部结构与温度和空气流动的关系等相关因素则少有提及[22-23]。此外,研究所涉及的生态类型和具体鼠种也与高寒草甸和高原鼠兔相差甚远,这也在一定程度上限制了其研究成果的可借鉴性。本研究结果表明,高原鼠兔洞系的复杂性取决于其对功能要求的多样性,其中,包括洞口朝向、数量、配置,洞道走向、长度、拐点数量和分布、曲折度和起伏度等特征因素。除与常规性功能需求密切相关而外,也是保持洞系栖居适合度,特别是调节洞系温度和空气流通性的重要考量。
就洞口朝向的数量配置而言,在本研究所调查的900个洞口中,朝阳(S、SW、SE)的洞口数最多,占到了洞口总数的41.3%,其次为朝阴(N、NW、NE)的洞口,占33.0%,半阴半阳的洞口数最少,占23.3%。同时,在洞口斜度≤55°的洞口中,朝阳的洞口数(48.6%)显著高于(P<0.01)朝阴的洞口数(27.8%),而在斜度>55°的洞口中则表现为朝阴的洞口数(52.1%)显著高于(P<0.01)朝阳的洞口数(24.3%)。由此可见,如果只考虑温度因素,高原鼠兔所应采取的策略是尽可能多地增加朝阳洞口的数量,减少朝阴洞口的数量,同时,增加斜度≤55°洞口的比重,减少斜度>55°洞口的比重,因为,只有如此方可在寒冷的冬季有效提升洞内的温度。尽管实际结果总体上符合这一原则,但显然仍存在一定的增减空间。这一方面说明,提高洞内温度是高原鼠兔选择洞系构造特征的一个主导性因素,但同时也说明,在决定高原鼠兔洞系舒适度的诸多因素中,温度并非唯一的主导性因素,在考虑温度因素的同时,显然也兼顾了与其他因素的权衡。
本研究中有关洞口温度的结果也同时印证了上述推论。洞口温度随洞口朝向的不同而存在明显差异,特别是S向和偏S向的洞口温度大都极显著高于N向和偏N向,其差值最高可达13.2℃;此外,W向和偏W向与E向和偏E向洞口的温度差异也较为显著,差值平均也在1.8~6.0℃。从洞系结构来看,洞道的基本走向大都以S—N向为主线,而且洞口也主要集中于S、N两向,另外,N向的洞口与主洞道的垂直距离普遍大于S向。可见,由于S、N向洞口间显著温差的存在,加之洞系特殊结构的共同作用,必然导致洞内空气对流的形成,而确保洞内空气的流动与更新,极有可能是决定高原鼠兔洞系构造特征的另1个重要因素。值得注意的是,在洞口朝向为全阴的3个方向上,EN向的洞口数占15.2%,明显高于WN向(13.2%),而正N向的洞口数极低,仅为4.6%。这样一种配置格局除了与温度和空气流动有关而外,很可能是对当地冬春季风向主要为N风和WN风,极少有EN风这一气候特点的一种适应性选择。
4 结论
高原鼠兔洞口朝向为全阴、半阴和全阳的洞口数分别占洞口总数的33.0%,23.3%和41.3%。
洞口直径与洞口朝向无相关性。洞口斜度≤55°的洞口主要集中在SW、SE和S3个方向,占洞口总数的48.6%;在E、W方向的占23.4%;而在NW、N和NE 3个方向洞口数仅为洞口总数的27.8%。洞口斜度>55°的洞口主要集中在NW、NE和N 3个方向,占洞口总数的52.1%;E、W方向的洞口占23.6%;SW、SE和S方向的洞口占24.3%。
S向和偏S向的洞口温度显著高于N向和偏N向的洞口,W向和偏W向洞口温度普遍高于E和偏E洞口,前者最高温差为13.2℃,后者最高温差为6.0℃。
高原鼠兔的洞系建筑特点是对高原环境的一种适应性选择,除了常规意义上的栖居和避险等基本功能而外,还兼顾了洞内温度、对流和抵御寒风的利弊权衡。
[1] Smith A T,Foggin J M.The plateau pika is a keystone species for biodiversity on the Tibetan Plateau[J].Animal Conservation,1999,2:235-240.
[2] Begall S,Gallardo M H.Spalacopuscyanus(Rodentia:Octodontidae):an extremist in tunnel constructing and food storing among subterranean mammals[J].Journal of Zoology,2000,251:53-60.
[3] Vleck D.Burrow structure and foraging costs in the fossorial rodent,Thomomys bottae[J].Oecologia,1981,49:391-396.
[4] Saundersa G,Cooke B,McColl K,etal.Modern approaches for the biological control of vertebrate pests:An Australian perspective[J].Biological Control,2010,52(3):288-295.
[5] Lai C H,Smith A T.Keystone status of plateau pikas(Ochotonacurzoniae):effect of control on biodiversity of native birds[J].Biodiversity and Conservation,2003,12:1901-1912.
[6] 施大钊,钟文勤.2000年我国草原鼠害发生状况及防治对策[J].草地学报,2001,9(4):248-252.
[7] 王兴堂,花立民,苏军虎,等.高原鼠兔的经济损害水平及防治指标研究[J].草业学报,2009,18(6):198-203.
[8] 施银柱.草场植被影响高原鼠兔密度的探讨[J].兽类学报,1983,3(2):181-187.
[9] 张卫国,丁连生,韩天虎.降水对高原鼠兔种群消长的影响[J].草业科学,1999,16(6):20-22.
[10] 张卫国,刘蓉,江小雷.风险性声讯信号对高原鼠兔行为模式的影响[J].草地学报,2010,(1):115-120.
[11] 边疆辉,刘季科.相关风险因子对高原鼠兔摄食行为的影响[J].兽类学报,2001,21(3):41-52.
[12] 张永超,牛得草,韩潼,等.补播对高寒草甸生产力和植物多样性的影响[J].草业学报,2012,21(2):305-309.
[13] 文淑均,张世挺,李伟,等.青藏高原东缘典型草甸与沼泽化草甸中种子雨的差异[J].草业学报,2012,21(2):256-263.
[14] Luna F C,Antinuchi C D.Effect of tunnel inclination on digging energetics in the tuco-tuco,Ctenomystalarum(Rodentia:Ctenomyidae)[J].Naturwissenschaften,2007,94:100-106.
[15] Lovegrove B G.The cost of burrowing by the social mole rats(Bathyergidae)CryptomysdamarensisandHeterocephalus glaber:The role of soil moisture[J].Physiological Zoology,1989,62(2):449-469.
[16] Vibe-Petersen S,Leirs H,de Bruyn L.Effects of predation and dispersal onMastomysnatalensispopulation dynamics in Tanzanian maize fields[J].Journal of Animal Ecology,2006,75:213-220.
[17] Le Comber S C,Eabloom E W,Romanach S S.Burrow fractal dimension and foraging success in subterranean rodents:a simulation[J].Behavioral Ecology,2005,11:188-194.
[18] Wang J M,Zhang Y M,Wang D H.Seasonal thermogenesis and body mass regulation in plateau pikas(Ochotonacurzoniae)[J].Oecologia,2006,149:373-382.
[19] Roci’O Torres M,Borghi C E,Giannonii S M.etal.Portal orientation and architecture of burrows in tympanoctomys barrerae(Rodentia,Octodontidae)[J].Journal of Mammalogy,2003,84(2):541-546.
[21] Thomas H G,Bateman P W,Scantlebury M,etal.Season but not sex influences burrow length and complexity in the nonsexually dimorphic solitary Cape mole-rat(Rodentia:Bathyergidae)[J].Journal of Zoology,2012,288(3):1-8.
[22] Rosi M I,Cona M I,Videla F,etal.Architecture ofCtenomysmendocinus(Rodentia)burrows from two habitats differing in abundance and complexity of vegetation[J].Acta Theriologica,2000,45(4):491-505.
[23] Edelman A J.Kangaroo rats remodel burrows in response to seasonal changes in environmental conditions[J].Ethology,2011,117(5):430-439.
