不同土壤水分条件下麻栎盆栽实生苗的光合响应
2013-01-02李辉谢会成姜志林李传荣张光灿
李辉,谢会成†,姜志林,李传荣,张光灿
(1.山东农业大学林学院,山东省土壤侵蚀与生态修复重点实验室,271018,山东泰安;2.南京林业大学 资源与环境学院,210037,南京)
土壤水分的主要来源是大气降水和灌溉水。土壤含水量对植物生长和代谢的影响是多方面的,其中对光合作用、蒸腾和气孔导度的影响尤为突出,土壤水分含量不足或过多都会影响植物的光合作用、蒸腾作用和气孔导度[1-2]。干旱经常威胁着人们的生存并制约着植物生产力的提高。植物生长对水分影响很敏感,轻微的水分胁迫就能抑制植物的生长。麻栎(Quercus acutissima Carruth)为壳斗科,栎属,落叶乔木,种子含淀粉和脂肪油,可酿酒、作饲料、制肥皂;壳斗、树皮含鞣质,可提取栲胶,木材坚硬、耐磨,可供机械用材。目前关于麻栎的研究大多集中在麻栎叶片水分、麻栎混交林、不同种源麻栎的差异性等方面[3-6],而涉及到麻栎一年生实生苗在水分胁迫下的净光合速率和羧化效率等生理方面的研究少见报道。笔者采用一年生盆栽麻栎实生苗,研究不同土壤含水量在弱光、强光下麻栎的光合速率,并利用实测值和直角双曲线修正模型对麻栎幼苗的光合作用进行光响应曲线拟合,以探讨不同土壤含水量对麻栎光合特征的影响,为麻栎林带更新及植被重建提供科学依据。
1 研究地概况
实验在南京林业大学树木园进行,该树木园位于江苏省南京市,地理位置E118°47',N32°3',属亚热带季风气候,具有明显的冬冷夏热的四季变化,年降雨量为1 000 mm 左右,年均温15.7 ℃,地貌类型为岗地,土壤为黄棕壤。
2 研究方法
2.1 实验材料与方法
实验采用当年生种子繁殖,种子采集于南京农业大学下蜀林场,设60 个花盆,每个花盆播2 粒种子,让其在适应的环境中生长,1 年后选择生长状况相同的12 盆麻栎小苗用于实验,采用人工控水,单因素对比实验设计。控水实验从2010-07-13开始。土壤含水量控制为4 个水平,分为对照组、处理组1、处理组2、处理组3,每个水平3 盆麻栎小苗,每3 d 浇一次水,每次浇水使对照达到饱和持水量,其他各处理组浇水量依次减为饱和持水量的70%、40%和30%。分别在控水15 d(07-28)后,每盆苗选取不同部位的5 片叶片,用Li-6400 光合作用测定系统测定净光合速率(Pn)和Pn-CO2变化曲线。同时测得平均土壤含水量:对照组为32.0%、处理组1为24.0%、处理组2 为14.5%、处理组3 为11.4%。
2.2 模型拟合
2.2.1 实测值 用Excel 软件得出净光合速率的光响应曲线,并对曲线的初始部分进行线性回归,由于麻栎在弱光条件下就能达到比较高的光合作用,即当光合有效辐射强度到达200 μmol/(m2·s)时,净光合速率就已经随着光照强度的增强而变化缓慢了;因此,在用Excel 软件对光响应曲线的初始部分进行线性回归时,选择光合有效辐射强度PAR<80 μmol/(m2·s),由线性回归的方程求得表观量子效率、光补偿点和暗呼吸速率[7-8]。根据图1 曲线走势估计光饱和点、最大净光合速率作为实测值与模型拟合值进行比较分析。
2.2.2 直角双曲线修正模型 通过前人的研究得出,在拟合C3植物的光响应曲线时,直角双曲线修正模型的计算值比较接近观测值(实测值),具有较高的精确性和适宜性[9];所以,笔者用直角双曲线修正模型对麻栎的光响应曲线进行模拟。通过SPSS17.0 软件,用直角双曲线修正模型对麻栎的净光合速率光响应数据进行非线性回归拟合,直角双曲线修正模型表达式[8,10-11]为:

式中:Pn为净光合速率,μmol/(m2·s);Φ 为观量子效率,mol/mol;PAR为光合有效辐射强度,μmol/(m2·s);Rd为呼吸速率,μmol/(m2·s);β、γ 为模型系数;LSP为光饱和点,μmol/(m2·s);Pmax为最大净光合速率,μmol/(m2·s)。
3 结果与分析
3.1 不同土壤水分条件下的麻栎光合活性
实验是在大棚内进行的,经测定,大棚内的光合有效辐射强度一般维持在160 μmol/(m2·s)左右;所以,在实验过程中,通过人工控光设定光合有效辐射强度为160 μmol/(m2·s),测量不同土壤含水量的麻栎一年生小苗净光合速率、蒸腾速率及其他相关参数。这个光合有效辐射强度也是大部分麻栎林下的光合有效辐射照强度。表1 示出不同土壤水分条件下麻栎幼苗的净光合作用特征。可知,在相同的光合有效辐射条件下,土壤含水量32.0%、24.0%、14.5%、11.4%之间麻栎的净光合速率两两比较,存在显著性差异(P <0.05)。说明麻栎土壤含水量从32%降低到11.4%的3 次降低中,每降低1 次土壤含水量都会对麻栎的净光合作用产生显著影响。在相同的光合辐射强度条件下,除了土壤含水量24.0%和14.5%的蒸腾速率之间的差异水平P=0.66 >0.05 差异不显著之外,其他各组两两之间的土壤含水量的蒸腾速率相比较,都存在显著性差异(P <0.05)。说明在相同光有效辐射条件下,土壤含水量从24.0%降低到14.5%时,对蒸腾速率的影响不大。在对不同土壤含水量的蒸腾速率进行聚类分析(欧式聚离最短距离法)时,土壤含水量24.0%和14.5%的蒸腾速率的距离在0.76(重新标定到0 ~2)时可聚为一类,也说明土壤含水量从24.0%降低到14.5%时对麻栎的蒸腾作用影响不大。气孔导度随着土壤含水量的降低而降低,胞间CO2摩尔分数则随着土壤含水量的降低先下降后上升,说明土壤含水量从32.0%降低到24.0%,净光合速率的下降是由气孔限制因素导致的,而土壤含水量从14.5%降低到11.4%,净光合速率的下降是由非气孔因素限制引起的。与对照组土壤含水量32.0%相比,土壤含水量24.0%、14.5%、11.4%的麻栎净光合速率的下降比例分别为14.2%、40.3%、70.2%,蒸 腾 速 率 的 下 降 比 例 分 别 为27.5%、31.6%、63.0%,在一定程度上说明麻栎的土壤含水量降低到11.4%时,其光合生理已受到严重抑制,长时间生长在此种土壤含水量的情况下,有可能造成麻栎小苗的死亡。
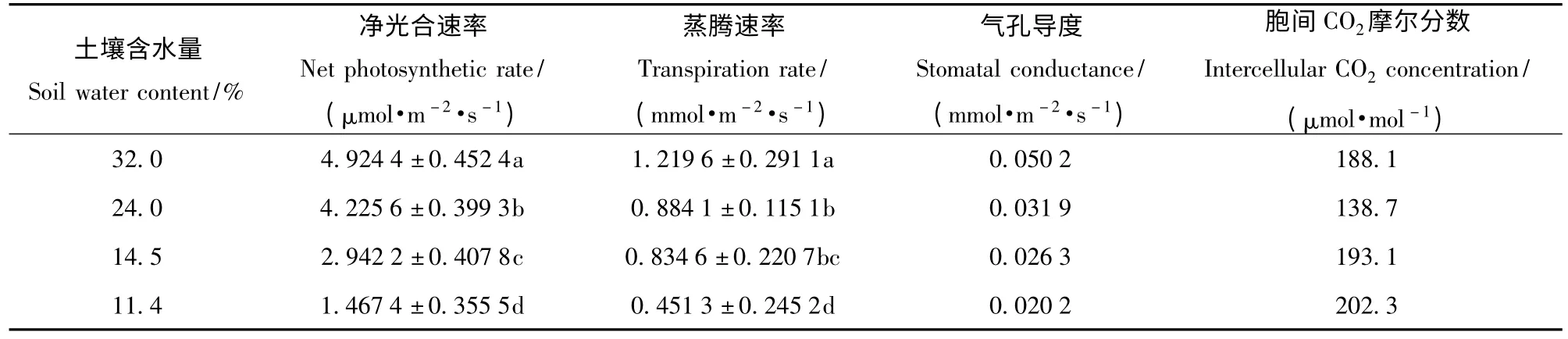
表1 不同土壤水分条件下麻栎幼苗的净光合作用特征Tab.1 Parameters of photosynthetic characteristics of Quercus acutissima seedlings
3.2 麻栎叶片光合作用光响应的模拟
由表2 可知,用直角双曲线修正模型对不同土壤含水量的麻栎光响应曲线进行拟合时,发现模型拟合的决定系数R2都超过0.99。表明此模型可以处理麻栎的光响应曲线问题,能较好地模拟光合响应过程。从直角双曲线修正模型反映的麻栎光合响应参数可以看出,直角双曲线修正模型拟合出的光响应数据除了光饱和点和实测值有较大的差异外,其他拟合的光响应数据相差不大,并随土壤含水量的变化和实测值表现出相同的趋势。这在一定程度上说明直角双曲线修正模型能很好地拟合麻栎的光响应曲线;但直角双曲线修正模型毕竟是光响应模型,决定系数R2不可能达到1,还是存在一定的不足之处。如实验用此模型拟合出的光饱和点就和实测值的光饱和点有一定的差距,因为当光合有效辐射强度大于0 时,净光合速率始终随着光合有效辐射强度的增加而增加。虽然由高等数学知识可计算出式(1)的极值点[12],但模型的模拟不会考虑植物实际在自然界中究竟在什么情况下进行的光合作用;因此,模型趋势线往往不能很好地给出正确的光饱和点。
3.3 麻栎叶片净光合作用的光响应
如图1 所示,当光合有效辐射强度PAR<200 μmol/(m2·s)时,各土壤含水量的净光合速率随着光合有效辐射强度的增强上升的比较快,基本成直线状,当PAR>400 μmol/(m2·s)时,各土壤含水量的净光合速率随着光有效辐射强度的增强上升缓慢,基本上是一条和X 轴(光有效辐射强度PAR)平行的曲线。在相同光照强度条件下,不同土壤含水量的麻栎幼苗的净光合速率都随着土壤含水量的上升而上升。当土壤含水量从32.0%降低到24.0%,再从24.0%降低14.5%时,麻栎的净光合速率成阶梯状下降;而当PAR<100 μmol/(m2·s)时,土壤含水量为14.5%的光响应曲线和土壤含水量为11.4%的光响应曲线基本重合,即当PAR>100 μmol/(m2·s)时,相同光照条件下土壤含水量从14.5%降低到11.4%时,麻栎小苗的净光合速率变化也不大。由此可以说明,弱光条件下,植物对土壤水分胁迫的忍耐性有所提高。
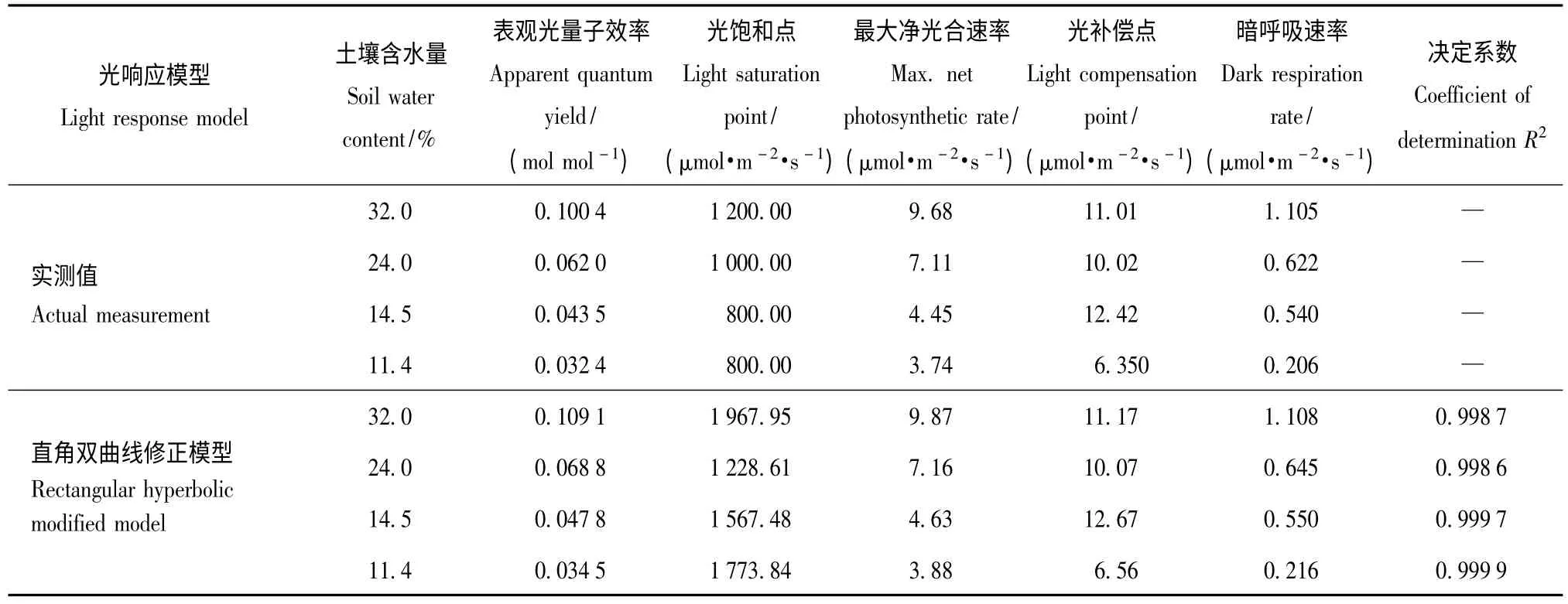
表2 用直角双曲线修正模型模拟的麻栎光合作用响应参数Tab.2 Parameters of photosynthesis-light response of Quercus acutissima fitted by modified rectangular hyperbola model
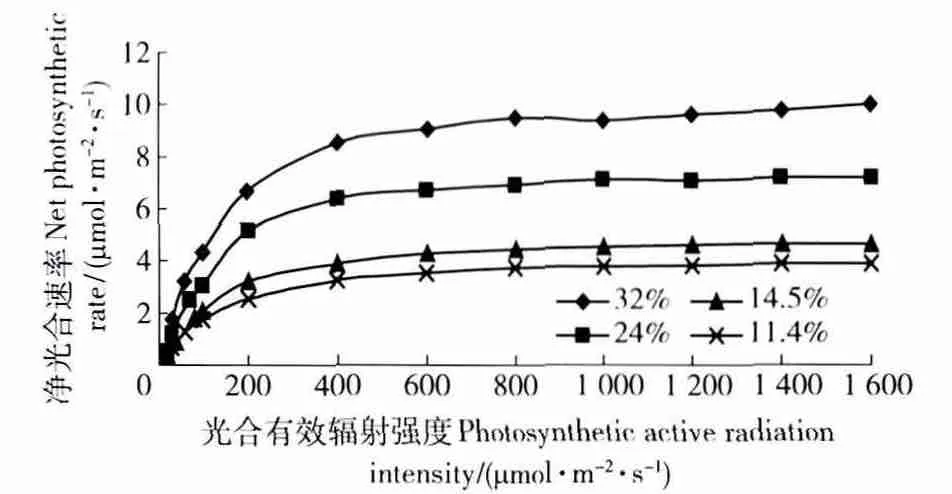
图1 麻栎净光合速率的光响应曲线Fig.1 Light response curve of net photosynthetic rate of Quercus acutissima
3.4 麻栎光合作用对CO2的响应
羧化效率反映了植物在给定条件下对胞间CO2的同化能力,叶片的羧化效率越高,说明光合作用对CO2的利用就越充分。在饱和光照条件下,胞间CO2摩尔分数小于200 μmol/mol 时,植物叶片的净光合速率对胞间CO2摩尔分数升高的响应为近似直线,这条直线的斜率即为叶肉导度或羧化效率[13-14],高光强度下(PAR为1 000 μmol/(m2·s)),叶片的光合羧化速率受羧化部位CO2摩尔分数的限制[15]。如图2 所示,在光有效辐射强度为1 000 μmol/(m2·s)时,不同的土壤含水量,麻栎净光合速率对CO2响应曲线不同,土壤含水量为32.0%、24.0%、14.5%、11.4%时,麻栎的羧化效率分别0.113 7、0.092 1、0.048 8 和0.047 4 mol/(m2·s),CO2补偿点的摩尔分数分别为43.96、40.74、39.24和38.93 μmol/mol,均表现为随着土壤含水量的降低而降低的趋势。这在一定程度上说明了土壤含水量的多少对麻栎幼苗光合作用中固定CO2的1,5-二磷酸核酮糖羧化酶(Rubisco)酶活性的大小产生影响,土壤含水量越高时,1,5-二磷酸核酮糖羧化酶活性越高,固定CO2的能力越强,麻栎的净光合速率越高。

图2 麻栎净光合作用对CO2的响应曲线Fig.2 CO2 response curve of photosynthesis rate of Quercus acutissima
4 结论
1)在相同的光合有效辐射强度160 μmol/(m2·s)下,麻栎小苗的生长受到水分的制约,其净光合速率、蒸腾速率、气孔导度都随着土壤含水量的降低而降低,胞间CO2摩尔分数随着土壤含水量的降低,先降低后升高。
2)直角双曲线修正模型拟合所得到的麻栎幼苗的各个光合作用参数与实际值符合得很好,且确定系数R2都达到0.99 以上,可以用植物光合作用光响应的直角双曲线修正模型来处理麻栎幼苗在相同温度和相同CO2摩尔分数条件下的光响应数据。
3)不同光辐射强度条件下,不同土壤含水量对麻栎的光合作用影响很大。在强光条件下,麻栎光合作用受土壤水分胁迫的影响较大,而在弱光条件下,麻栎对土壤水分胁迫的忍耐性有所提高。
4)在光合有效辐射强度为1 000 μmol/(m2·s)时,麻栎的羧化效率和CO2补偿点的摩尔分数都随着土壤含水量的降低而降低,水分胁迫抑制了羧化酶的活性。
