西双版纳食果鸟对植物果实颜色的取食选择
2012-12-25权锐昌
段 琼, 权锐昌
(1. 中国科学院西双版纳热带植物园, 云南 西双版纳 666303; 2. 中国科学院研究生院,北京100049)
西双版纳食果鸟对植物果实颜色的取食选择
段 琼1,2, 权锐昌1,*
(1. 中国科学院西双版纳热带植物园, 云南 西双版纳 666303; 2. 中国科学院研究生院,北京100049)
黑色和红色是最主要的果实颜色。虽然食果鸟的取食选择常被假设为果实颜色特征形成的主要进化动力, 但能直接支持该假说的研究案例极少。该研究选择西双版纳常见的5种食果鸟, 利用试验当季常见的黑、红、黄、绿4种果实进行室内和野外环境(开阔林地、林冠下层)果实颜色选择实验。结果表明, 室内试验时5种食果鸟最喜欢的果实颜色是黑色和/或红色, 而野外环境下, 虽然开阔林地果实被取食总量远大于林冠下层果实被取食量, 但黑色和红色仍均为该两类生境中被取食最多的果实颜色。即大部分食果鸟均喜好自然界最常见的果实颜色,其喜好程度能反映研究地果实颜色组成分布格局, 提示果实颜色的进化或部分来自于食果鸟取食选择的压力。
果实颜色; 林缘; 林下; 动植物关系; 果实颜色比例格局
大部分陆生植物依赖食果动物为其传播种子和花粉, 如植物通过产生鲜艳的花或果实来吸引传粉者和种子散布者。因此, 颜色常被认为是增大被传播媒介发现几率的适应结果 (Willson & Whelan,1990; Schmidt et al, 2004; Lomáscolo et al, 2010)。自然界最常见的果实颜色是与自然背景颜色对比度最大的黑色和红色, 占所有成熟果实的 50%~70%(Wheelwright & Janson, 1985; Willson & Whelan,1990; Lee et al, 1994; Schmidt et al, 2004)。若果实颜色在吸引传播者中起信号作用, 那么黑色和红色被食果动物发现和捕食的几率也相对较大。但是, 到目前为止, 果实颜色与食果动物取食选择之间的相互关系仍不清楚。
为了探究种子散布者在果实颜色演化中的意义, 很多研究利用野外捕获的食果鸟来进行实验,结果各异。虽然部分研究发现食果鸟有特定的颜色偏好 (McPherson, 1988; Willson & Comet, 1993;Willson, 1994; Puckey et al, 1996; Siitari et al, 1999;Hartley et al, 2000), 但还有一些研究表明食果鸟取食选择与果实颜色无关或关系很弱, 且该选择在种内和种间存在很大差异 (Willson & Whelan, 1990;Willson & Comet, 1993)。然而这些研究多来自美洲或欧洲, 几乎没有来自热带亚洲的相关报道, 尽管热带亚洲拥有全球最丰富的食果鸟种类 (Corlett,1998), 且主要依靠鸟类传播种子而不同于热带美洲依靠蝙蝠传播种子 (Ingle, 2003; Muscarella &Fleming, 2007)。因此, 热带亚洲是探讨食果鸟取食选择与果实颜色相互关系的理想之地。
除颜色外, 影响食果鸟选择食物的因素仍很多,如果实大小、软硬度、可食性、营养成分和果实产量等 (Gautier-Hion et al, 1985; Whelan & Willson,1994; Schaefer et al, 2007)。由于鸟类主要依靠视觉搜索食物, 颜色无疑是影响食果鸟选择食物的主要因素之一 (Schaefer, 2011)。Lomáscolo et al (2010)在分子水平分析植物果实的性状演化历史, 发现颜色不同于其他性状, 其产生与演化无法用系统进化来解释, 而与食果鸟的选择等环境因素关系更为密切。食果鸟具有高度发达的四色视觉系统, 可见光范围为 300~700 nm(人类为 400~700 nm )(Vorobyev & Osorio, 1998)。果实颜色作为第一视觉信号无疑是食果鸟选择食物最重要的影响因素 (Schaefer, 2011)。一些研究采用人工果实的方法来单一讨论颜色与取食的关系 (Levey & Grajal,1991; Whelan & Willson, 1994; Alves‐Costa &Lopes, 2001), 但自然情况更为复杂, 如果直接利用自然果实试验也能得出相似的结论或许能更好解释食果鸟对果实颜色的取食选择。
此外, 生境对食果鸟的食物选择也有很大影响。在热带雨林中, 林冠与林下的光照、湿度和温度等较大环境因素差异(Molofsky & Augspurger,1992; Becerra et al, 2004; Ayres et al, 2009)导致了林冠与林下的不同动、植物的组成及其生活习性(Munn & McDaniel, 1985; Canham, 1988)。林冠光照环境较稳定, 植物果实林冠更为丰富, 因此, 大部分食果鸟均活动于林冠或光线充足的开阔地, 且较少在垂直方向来回活动。而在林下分布较多的食虫或杂食性鸟通常采取机会式的取食果实方式(Malmborg & Willson, 1988; Loiselle & Blake,1991)。因此, 不同生境下食果鸟的果实颜色选择可能不同。
西双版纳位于热带亚洲北缘, 已经调查的该地626种高等植物果实颜色组成比例表明, ~40%为黑色, 19%为红色, 13%为棕色, 13%为黄色, 8%为双色, 4%为绿色, 2%为白色以及1%为蓝色 (Chen et al, 2004)。如果食果鸟取食影响果实颜色进化, 我们首先假设食果鸟对果实某些颜色表现强烈偏好, 自然界高比例的果实颜色应该被更多的鸟类所选择。其次, 如果颜色是影响取食选择的主要因素, 那么不同的光照环境对取食选择影响则相对较小。
1 材料与方法
1.1 实验地点与材料
选择西双版纳热带植物园为野外实验地点。沟谷雨林林冠连续, 林下光照弱, 而野菜园林冠开阔,林下光照强。两者分别代表不同光照条件的生境试验材料包括室内试验所需的食果鸟和植物果实。食果鸟为由野外迷网捕获的10只红耳鹎, 8只黑冠黄鹎, 4只蓝喉拟啄木鸟, 2只灰眼短脚鹎, 和2只褐脸雀鹛。为避开繁殖影响, 所有试验用鸟在2011年1月—2月获得并单独饲养于30 cm×30 cm×40 cm的鸟笼内, 自由饮水, 试验前主要食物为苹果、梨、芭蕉、小鸡饲料和少量面包虫。
植物果实选取试验当季大量成熟, 在野外观察到有食果鸟采食, 并能反映西双版纳果实颜色不同组成比例格局的4种果实, 颜色分别为黑色、红色、黄色和绿色。黑果采自潺槁木姜子 (Litsea glutinosa), 乔木, 成熟果实为黑色; 红果采自催吐萝芙木 (Rauvolfia vomitoria), 乔木, 果实从最初的绿色转为橙黄色,最后成熟时为红色; 黄果采自假连翘 (Duranta erecta), 小乔木, 四季挂果, 成熟果实为黄色, 观察到的食果鸟拜访次数较少; 绿果采自布渣叶 (Microcos nervosa), 绿色, 未成熟果实为绿色, 果期很长, 观察到食果鸟取食, 成熟果实为黑褐色, 成熟期很短。所有试验用果大小、形状和软硬度等均相似, 直径6~8 mm, 圆球形, 较硬。
1.2 实验设计
1.2.1 果实颜色测定 鸟类与人的视觉系统差异大, 利用仪器测量各颜色光谱更能准确描述果实颜色。为避免肉眼对颜色分辨造成的误差, 我们使用海洋光学公司生产的 USB4000光谱仪测量各颜色光谱, 标准光源为海洋光学公司生产的氚卤DT-MINI-2-GS光源。各个被测物的反射率通过标准白光对照 (Top Sensor Systems WS-1-SS) 校准。为避免环境光干扰, 光纤探头 (QR400-2-UV-BX,Ocean Optics)固定于黑色塑料管中进行测量, 测量角度为45°。每个颜色的植物果实各测量6个, 取平均值作为各颜色植物果实的反射率。最后将光谱测量值以 20 nm的间隔平均值用 SigmaPlot软件作300~700 nm波长范围的光谱图。
1.2.2 室内果实颜色选择 试验集中在 2011年7—8月, 每天的试验时间为9:00—11:00和13:00—15:00。试验前一天下午采集植物果实, 4 ℃保存。所有试验用鸟在试验前1~1.5 h不喂食, 以确保试验时每个个体都有相似的取食意向。试验时每只试验鸟提供32粒果实(4种颜色×8粒)以确保提供的食物总量远大于实际取食量, 15 min后取出摆放食物的培养皿计数剩余的果实数量, 从而确定每次实验时各颜色果实的取食数量。每个试验重复 8次,持续3~5 d。
1.2.3 野外果实颜色选择 试验时间为 2011年7—8月。试验开始前, 在沟谷雨林和野菜园分别选取能反映光照强、弱的16株高度为2~3 m的灌木或小乔木用于人工挂果。为了避免树间隔太近所导致的数据不独立, 树间距≥5 m。挂果时, 选取每棵树高度在1.5 m 处的2个枝条, 用面糊在该枝条中部固定 20个同一颜色的果子。每个颜色果实挂 4棵树。每天7:00开始试验, 17:00 结束试验并检查记录被取食数量, 持续10天/重复10次。每天试验开始前更换所有果实以确保每次重复试验时果实新鲜程度一致。为避免空间位置影响鸟类的果实选择, 每棵树每天随机更换不同颜色的果实。
1.2.4 数据分析x2-检验及 Mann-Whitney U-tests检测食果鸟对果实颜色的取食偏好, 所有数据分析在SPSS 16.0 中完成。
2 结 果
2.1 果实颜色光谱值
黑、红、黄、绿4种颜色的植物果实光谱分布如图 1, 每个颜色的光谱最高反射率都与各颜色标准波长范围一致 (黑色光反射率几乎为零; 红色为630~750 nm; 黄色为~600 nm; 绿色为~550 nm),能真实代表这4种选定的果实颜色。
2.2 室内果实颜色选择
5种实验用鸟均对4种果实颜色表现强烈偏好(红耳鹎:x2=127.6,df=3,P=0.000,n=80; 黑冠黄鹎:x2=29.3,df=3,P=0.000,n=64; 褐脸雀鹛:x2=52.7,df=3,P=0.000,n=16; 灰眼短脚鹎:x2=12.7,df=3,P=0.005,n=16; 蓝喉拟啄木鸟:x2=53.1,df=3,P=0.000,n=32)。红耳鹎(10只)偏好红色果实; 黑冠黄鹎 (8只), 灰眼短脚鹎(2只)和蓝喉拟啄木鸟(4只)黑色和红色果实好; 而褐脸雀鹛(2只)则偏好黑色果实(图 2)。
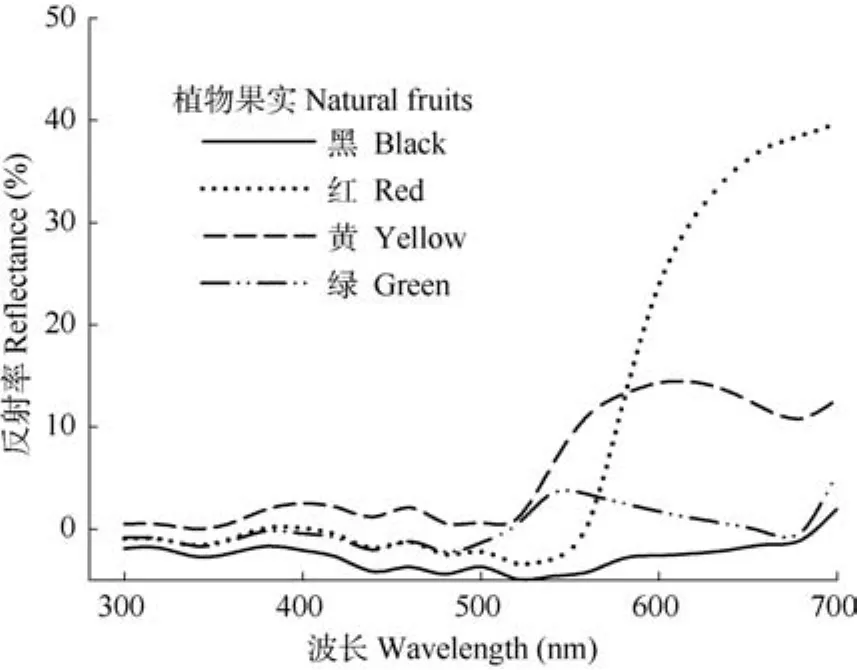
图1 植物果实 (黑、红、黄、绿)在300~700 nm范围内的光谱图Fig. 1 Spectra of natural fruits from 300−700 nm illustrating true colory
2.3 野外果实颜色选择
无论在光线较弱的沟谷雨林林下还是在光线充足的开阔地野菜园, 食果鸟都对果实颜色有明显的选择偏好(沟谷林林下:x2=9.7,df=3,P=0.022; 野菜园开阔地:x2=84.1,df=3,P=0.000)。红色和黑色均为这两类生境中食果鸟最喜好的果实颜色, 二者差异不显著 (x2= 0.001,df=1,P>0.05)。但在开阔地野菜园被消耗的果实总量要远远高于沟谷林林下的消耗量 (x2=73.4,df=1,P=0.000) (图3)。
3 讨 论
通过选择代表自然界果实颜色组成分布比例从多到少的 4种果实(黑、红、黄、绿)来进行食果鸟取食颜色选择试验, 我们发现无论是室内还是野外试验, 食果鸟均对果实颜色表现强烈的取食偏好(图2, 3)。该结果不同于已有的来自亚洲以外的很多相关研究, 此类研究大多表明食果鸟对果实颜色没有选择或呈现弱选择, 且这些选择有些只是暂时的,会随环境而改变 (Willson & Whelan, 1990; Willson& Comet, 1993; Schmidt et al, 2004)。这些结论上的差异或许归因于热带亚洲所拥有的全球最多的食果鸟种类 (Corlett, 1998), 且该地区森林更新也主要依赖鸟类传播种子而不同于热带美洲主要依靠蝙蝠散布种子 (Ingle, 2003; Muscarella & Fleming,2007)。鸟类主要依靠视觉搜寻食物, 因此在热带亚洲为代表的旧大陆地区, 食果鸟与果实颜色的相互关系要比在热带美洲等新大陆地区更加紧密。
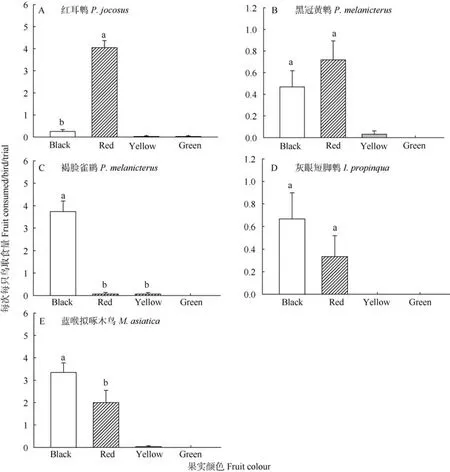
图2 室内试验用鸟对果实颜色所表现的强烈取食偏好Fig. 2 Each species showed a strong preference for certain color

图3 野外试验时不同颜色果实被取食情况Fig. 3 Consummation of different color fruits in open field experiment
试验中, 所有鸟类均对颜色表现强烈的选择偏好。其中, 红耳鹎显著偏好红色果实; 褐脸雀鹛则更加集中取食黑色果实, 且这两种鸟都几乎只选择一种颜色的果实。黑冠黄鹎、灰眼短脚鹎和蓝喉拟啄木鸟均取食两种颜色的植物果实(红色和黑色),对果实颜色的选择范围相对较宽。虽然营养是影响取食的重要因素之一 (Schaefer et al, 2008b;Wellmann & Downs, 2009), 但我们的研究结果更支持颜色的主导影响地位。实验时所有的鸟类均给予相同的 4种植物果实, 若营养决定最终取食选择,那么所有的鸟类或许都会集中取食某一种营养最高的果实, 但本试验结果难以支持该推理, 如红耳鹎和褐脸雀鹛的选择则完全不同。同时, 我们也注意到颜色对比度也会在一定程度上影响取食选择,红色和黑色与自然界常见的绿色背景间的颜色对比度大, 易于被鸟类发现并取食 (Cazetta et al,2007; Schaefer et al, 2008a), 这或许也是红色和黑色被取食最多的原因之一。当然, 食果鸟对黑色和红色的喜好可能还是由于该颜色是某些特殊营养的指示, 如花青素等抗氧化合物在红色和黑色果实中含量较多, 而有些研究支持鸟类能够识别并选择含有较高浓度花青素的食物 (Schaefer et al, 2008a;Schaefer, 2011)。但本试验所选取的红色和黑色果实是否也含有较高浓度的花青素还需要进一步验证。
由于不同实验材料和研究地点间鸟类组成的差异, 食果鸟颜色选择的野外试验结果差异很大。如Galetti et al (2003) 应用粘土制成的红、黑、白3种人工果实来研究野外环境下不同颜色果实的被取食情况, 发现黑色和红色是野外环境下被取食最多的果实颜色。而 Willson & Melampy (1983)和Traveset & Willson (1998) 的研究结果则表明野外环境下无颜色选择差异。本研究的野外试验则得到了类似Galetti et al (2003)的结果, 在光照较弱的沟谷雨林林下和光照较强的野菜园, 黑色和红色果实的被取食量均为最大值。野菜园被取食的红色和黑色果实总量远大于其在沟谷雨林林下被取食的数量, 这或许是由于野菜园的食果鸟种类和数量多于沟谷林林下(段琼, 未发表数据)。同时, 野菜园视野宽阔, 黑色和红色因其与背景颜色间的较高对比度易于被远距离发现, 这也会导致野菜园的果实被取食量相对较多。
本研究是在热带亚洲完成的有关果实颜色与食果鸟食物选择相互关系的少数研究之一。结果表明常见的食果鸟均表现强烈取食颜色偏好, 且主要为黑色和红色。但同时, 这些食果鸟也取食少量的其他颜色果实。因此, 我们认为食果鸟对果实颜色的选择格局在一定程度反映了自然界果实颜色比例分布格局, 如:颜色比例最高的红色和黑色是被取食最多的果实颜色, 而比例较低的黄色和绿色被取食的比例也相对较低。
致谢:感谢西南林业大学的刘锋和李焕对野外工作的支持与帮助; 感谢李梦帮助捕获及饲喂试验用鸟。湖北大学的甘文瑾博士研究生在颜色光谱测量上给予了无私帮助, 两位评委对文章的初稿提出了宝贵意见及建议, 在此一并致谢。
A lves-Costa CP, Lopes AV. 2001. Using artificial fruits to evaluate fruit selection by birds in the field [J].Biotropica,33(4): 713-717.
Ayres E, Steltzer H, Simmons BL, Simpson RT, Steinweg JM, Wallenstein MD, Mellor N, Parton WJ, Moore JC, Wall DH. 2009. Home-field advantage accelerates leaf litter decomposition in forests [J].Soil Biol Biochem,41(3): 606-610.
Becerra PI, Celis‐ Diez JL, Bustamante RO. 2004. Effects of leaf litter and precipitation on germination and seedling survival of the endangered tree Beilschmiedia miersii [J].Appl Veg Sci,7(2): 253-257.
Canham CD. 1988. Growth and canopy architecture of shade-tolerant trees:response to canopy gaps [J].Ecology,69(3): 786-795.
Cazetta E, Schaefer HM, Galetti M. 2007. Why are fruits colorful? The relative importance of achromatic and chromatic contrasts for detection by birds [J].Evol Ecol,23(2): 233-244.
Chen J, Fleming TH, Zhang L, Wang H, Liu Y. 2004. Patterns of fruit traits in a tropical rainforest in Xishuangbanna, SW China [J].Acta Oecol,26(2): 157-164.
Corlett RT. 1998. Frugivory and seed dispersal by vertebrates in the Oriental (Indomalayan) Region [J].Biol Rev,73(4): 413-448.
Galetti M, Alves-Costa CP, Cazetta E. 2003. Effects of forest fragmentation,anthropogenic edges and fruit colour on the consumption of ornithocoric fruits [J].Biol Conserv,111(2): 269-273.
Gautier-Hion A, Duplantier JM, Quris R, Feer F, Sourd C, Decoux JP,Dubost G, Emmons L, Erard C, Hecketsweiler P. 1985. Fruit characters as a basis of fruit choice and seed dispersal in a tropical forest vertebrate community [J].Oecologia,65(3): 324-337.
Hartley L, Waas J, O'Connor C, Matthews L. 2000. Colour preferences and coloured bait consumption by weka Gallirallus australis, an endemic New Zealand rail [J].Biol Conserv,93(2): 255-263.
Ingle NR. 2003. Seed dispersal by wind, birds, and bats between Philippine montane rainforest and successional vegetation [J].Oecologia,134(2):251-261.
Lee WG, Weatherall I, Wilson JB. 1994. Fruit conspicuousness in some New Zealand Coprosma (Rubiaceae) species [J].Oikos,69(1): 87-94.
Levey DJ, Grajal A. 1991. Evolutionary implications of fruit-processing limitations in cedar waxwings[J].Am Nat,138(1): 171-189.
Loiselle BA, Blake JG. 1991. Temporal variation in birds and fruits along an elevational gradient in Costa Rica [J].Ecology,72(1): 180-193.
Lomáscolo SB, Levey DJ, Kimball RT, Bolker BM, Alborn HT. 2010.Dispersers shape fruit diversity in Ficus (Moraceae) [J].Proc Natl Acad Sci USA,107(33): 14668-14672.
Malmborg PK, Willson MF. 1988. Foraging ecology of avian frugivores and some consequences for seed dispersal in an Illinois woodlot [J].Condor,90(1): 173-186.
McPherson JM. 1988. Preferences of cedar waxwings in the laboratory for fruit species, colour and size: a comparison with field observations[J].Anim Behav,36(4): 961-969.
Molofsky J, Augspurger CK. 1992. The effect of leaf litter on early seedling establishment in a tropical forest [J].Ecology,73(1): 68-77.
Munn L, McDaniel P. 1985. Effect of temperature on organic carbon-texture relationships in Mollisols and Aridisols [J].Soil Sci Soc Am J,49(6):1486-1489.
Muscarella R, Fleming TH. 2007. The role of frugivorous bats in tropical forest succession [J].Bioll Rev,82(4): 573-590.
Puckey HL, Lill A, O'Dowd DJ. 1996. Fruit color choices of captive silvereyes (Zosterops lateralis) [J].Condor,98(4): 780-790.
Schaefer HM. 2011. Why fruits go to the dark side [J].Acta Oecol,37(6):604-610.
Schaefer HM, McGraw K, Catoni C. 2008a. Birds use fruit colour as honest signal of dietary antioxidant rewards [J].Funct Ecol,22(2): 303-310.
Schaefer HM, Schaefer V, Vorobyev M. 2007. Are fruit colors adapted to consumer vision and birds equally efficient in detecting colorful signals?[J].Am Nat,169(S1): S159-S169.
Schaefer HM, Spitzer K, Bairlein F. 2008b. Long-term effects of previous experience determine nutrient discrimination abilities in birds [J].Front Zool,5(1): 4.
Schmidt V, Martin Schaefer H, Winkler H. 2004. Conspicuousness, not colour as foraging cue in plant–animal signalling [J].Oikos,106(3):551-557.
Siitari H, Honkavaara J, Viitala J. 1999. Ultraviolet reflection of berries attracts foraging birds. A laboratory study with redwings (Turdus iliacus) and bilberries (Vaccinium myrtillus) [J].P Roy Soc B- Biol Sci,266(1433): 2125-2129.
Traveset A, Willson MF. 1998. Ecology of the fruit-colour polymorphism in Rubus spectabilis [J].Evol Ecol,12(3): 331-345.
Vorobyev M, Osorio D. 1998. Receptor noise as a determinant of colour thresholds[J].P Roy Soc B-Biol Sci,265(1394): 351-358.
Wellmann A, Downs C. 2009. Sugar preferences and digestion by Cape white-eyes, Zosterops virens, fed artificial fruit diets [J].Afr Zool,44(1): 106-116.
Wheelwright NT, Janson CH. 1985. Colors of fruit displays of bird-dispersed plants in two tropical forests [J].Am Nat,126(6):777-799.
Whelan CJ, Willson MF. 1994. Fruit choice in migrating North American birds: field and aviary experiments [J].Oikos,71(1): 137-151.
Willson MF. 1994. Fruit choices by captive American robins [J].Condor,96(2): 494-502.
Willson MF, Comet TCA. 1993. Food choices by northwestern crows:experiments with captive, free-ranging and hand-raised birds [J].Condor,95(3): 596-615.
Willson MF, Melampy MN. 1983. The effect of bicolored fruit displays on fruit removal by avian frugivores [J].Oikos,41(1): 27-31.
Willson MF, Whelan CJ. 1990. The evolution of fruit color in fleshy-fruited plants [J].Am Nat,136(6): 790-809.
Natural fruit colour selection by frugivorous birds in Xishuangbanna
DUAN Qiong1,2, QUAN Rui-Chang1,*
(1.Xishuangbanna Tropical Botanical Garden, the Chinese Academy of Sciences, Xishuangbanna Yunnan666303,China;2.Graduate School of the Chinese Academy of Sciences, Beijing100049,China)
Black and red are the most common colors of fruit, but the reason behind this has been subject to debate.Food preferences of avian frugivores for certain colors of food have been proposed as a selection mechanism that explains these traits, but there is little evidence supporting this hypothesis. Here, we conducted a lab experiment using four colors of natural fruit to evaluate color preferences of five avian species, and we also conducted this experiment in open area and understory habitats. Our results showed that red and black fruits were selected most often in lab experiment;in field experiment, red and black fruits were also the most preferred food, but the total amount of consumed fruits differed significantly between open areas and understory habitats. Our study suggested that differences in color preferences among frugivores may potentially reflect the diversity of fruit color and frequency in Xishuangbanna.
Fruit color; Forest edge; Understory; Plant-animal interaction; Fruit color distribution pattern
Q958.12
A
0254-5853-(2012)05-0427-06
10.3724/SP.J.1141.2012.05427
2012-05-10;接受日期:2012-06-01
中国科学西部博士基金和中科院青年科技人员方向项目(KSCX2-EW-Q-17)∗
Corresponding author),权锐昌, E-mail: quanrc@xtbg.ac.cn
段琼(1987−), 女, 云南丽江人, 硕士研究生, 主要从事动植物关系研究
