核桃优良品种枝条抗寒性生理生化指标分析
2012-12-23宋艳波乔永胜石美娟武彦霞刘朝红樊新萍
王 勇,宋艳波,乔永胜,石美娟,武彦霞,刘朝红,田 鑫,樊新萍,安 栋
(1.山西省农业科学院果树研究所,山西太谷030815;2.山西农业大学生命科学学院,山西太谷030801)
核桃(Juglans regia L.)属核桃科落叶乔木[1],在山西分布广泛,南北均有栽培,尤其以太行、吕梁等丘陵地区分布最多[2]。我国北方地区核桃幼树普遍存在着不同程度的低温伤害,主要包括冬季低温冻害和早春霜害[3]。核桃幼树遭受低温冻害后,部分主枝、多年生大枝及1 年生枝会死亡,当年大量减产,甚至绝收。国内外许多学者已在果树抗寒机理、抗寒性鉴定、抗寒资源评价及抗寒性遗传育种等领域进行了大量研究,并取得了良好的进展[4-9]。王勇等[10]对核桃枝条电解质渗出率与核桃树的抗寒性关系进行了研究,结果表明,品种间和不同处理温度间的电解质渗出率均存在极显著差异,核桃枝条电解质渗出率随处理温度的降低而逐渐升高;不同树龄间的电解质渗出率存在显著差异,核桃成龄树1 年生枝条抗寒性略高于幼龄树。王勇等[10]还对核桃的1 年生枝条的抗寒性指标进行了测定分析,结果表明,元丰、辽核4 号、辽核1 号比阿7、扎343 品种比较抗寒;对抗寒性指标进行分析比较,品种间抗寒性差异较大时可用电导率法测定,此法既简便又节省费用;抗寒性差异小时可用总酚、黄酮类物质、可溶性糖和淀粉含量进行抗寒性综合鉴定。
本试验在前人研究基础上[11-16],以礼品1 号、礼品2 号、中林1 号、中林5 号、辽核1 号、辽核3号和晋龙1 号7 个山西主推的核桃品种为试材,对其生育后期和休眠期的新梢和1 年生枝条的抗寒性生理生化指标,包括可溶性糖、可溶性蛋白、脯氨酸含量及超氧化物歧化酶(SOD)、过氧化物酶(POD)活性等进行了分析测定,旨在明确各指标的变化规律,分析其与核桃抗寒性之间的关系,并对各个品种的抗寒性进行比较,为核桃品种的选择及合理栽培管理提供理论依据。
1 材料和方法
1.1 试验材料
试验在山西省农业科学院果树研究所进行,供试品种为礼品1 号、礼品2 号、中林1 号、中林5 号、辽核1 号、辽核3 号和晋龙1 号,树龄5 年生,每个品种选取生长状况基本相同的植株6 株,2010 年9 月底至2011 年2 月底,每隔1 个月采样一次,从每株树树体中上部四周随机采取整齐均匀、粗细相近的新梢和1 年生枝条1 个组成混合样,用保鲜袋装好带回实验室。用自来水和去离子水冲洗多次,然后用洁净的纱布擦干,最后用保鲜袋分装备用。每个处理3 次重复。
主要试剂有:磷酸、愈创木酚、甲基氨酸(met)、氮蓝四唑(nbt)、核黄素、EDTA、考马斯亮蓝G250、蒽酮- 乙酸乙酯、90%的乙醇、茚三酮、冰醋酸、磺基水杨酸。试验器材有:UV-2450 分光光度计、PRZMOR 多功能离心机、BT 244S 电子天平。
1.2 试验方法
1.2.1 可溶性糖的测定 参照乔富廉[17]的方法测定各样品可溶性糖含量。
1.2.2 可溶性蛋白含量测定 采用考马斯亮蓝G250 法测定各样品可溶性蛋白含量[18]。
1.2.3 脯氨酸含量测定 参照乔富廉[17]的方法测定各样品脯氨酸含量。
1.2.4 酶活性的测定 参照李合生等[19]的方法测定各样品酶活性。
2 结果与分析
2.1 可溶性糖含量的变化
从图1 可以看出,随着核桃生育进程的继续以及气温的变化,除中林5 号和辽核3 号核桃枝条的可溶性糖含量一直呈上升趋势外,其余5 个品种枝条的可溶性糖含量整体呈先升高后降低的明显变化。在12 月多数品种可溶性糖含量大幅增加,在最低温之前均达到最大,这是由于初期的低温锻炼,激发了植物体内可溶性糖的积累,以此来提高植物的抗寒性。之后除中林5 号、辽核3 号枝条的可溶性糖含量继续上升外,其余5 个品种均开始出现小幅下降趋势,可能是由于这5 个品种正在逐步解除自然休眠,而中林5号、辽核3 号可能仍处在自然休眠状态。

不同品种间可溶性糖含量比较如表1 所示。

表1 不同品种间可溶性糖含量比较μg/g
从表1 可以看出,各品种在越冬过程中枝条可溶性糖含量存在明显差异,依其枝条可溶性糖含量的高低,推断其抗寒性强弱顺序依次为:辽核3 号>礼品1 号>礼品2 号>中林1 号>中林5 号>晋龙1 号>辽核1 号。
2.2 可溶性蛋白含量的变化
植物体内可溶性蛋白含量是一个重要的生理生化指标,当遭受低温胁迫时,体内蛋白质含量就会有所积累,以调节植物抗寒作用。

由图2 可知,在整个越冬过程中供试核桃枝条的可溶性蛋白含量变化比较一致,呈降—升—降—升的“W”趋势。随着气温下降,7 个品种可溶性蛋白质含量大幅增加,到11 月上旬,可溶性蛋白质含量最高。从整体上看,以晋龙1 号增加幅度最大,含量最高,可见其对寒冷的调节作用最强;礼品1 号增幅最小,对寒冷的调节作用最弱。
从表2 可以看出,各品种在越冬过程中枝条可溶性蛋白含量存在差异,辽核3 号和礼品1 号之间的可溶性蛋白含量差异显著,其他各品种间差异不显著。依各品种枝条可溶性蛋白含量的高低,推断其抗寒性强弱顺序依次为:辽核3 号>晋龙1 号>中林1 号>礼品2 号>中林5 号>辽核1 号>礼品1 号。

表2 不同品种间可溶性蛋白含量比较μg/g
2.3 脯氨酸含量的变化

由图3 可知,2010 年9—12 月,温度逐渐降低的过程中,各品种脯氨酸含量在逐渐升高。有6 个核桃品种在越冬过程中枝条脯氨酸含量基本呈先升高后降低的状态,而中林5 号枝条中脯氨酸含量一直呈升高的状态。但是在整个过程中,脯氨酸的平均含量仍然是辽核3 号最高,辽核1 号最低。然后随着核桃生育进程的继续以及气温的变化,供试品种除中林5 号的脯氨酸含量继续上升外,其他品种都有不同幅度的下降。这可能是由于中林5 号仍然处于自然休眠状态,而其他品种已经解除自然休眠的原因。
由表3 可知,辽核3 号与礼品1 号、晋龙1 号、辽核1 号间差异显著;辽核3 号、中林5 号与辽核1 号间差异极显著;其他各品种间差异不显著。依7 个品种枝条脯氨酸含量的高低,推断其抗寒性顺序依次为:辽核3 号>中林5 号>中林1 号>礼品2 号>礼品1 号>晋龙1 号>辽核1 号。
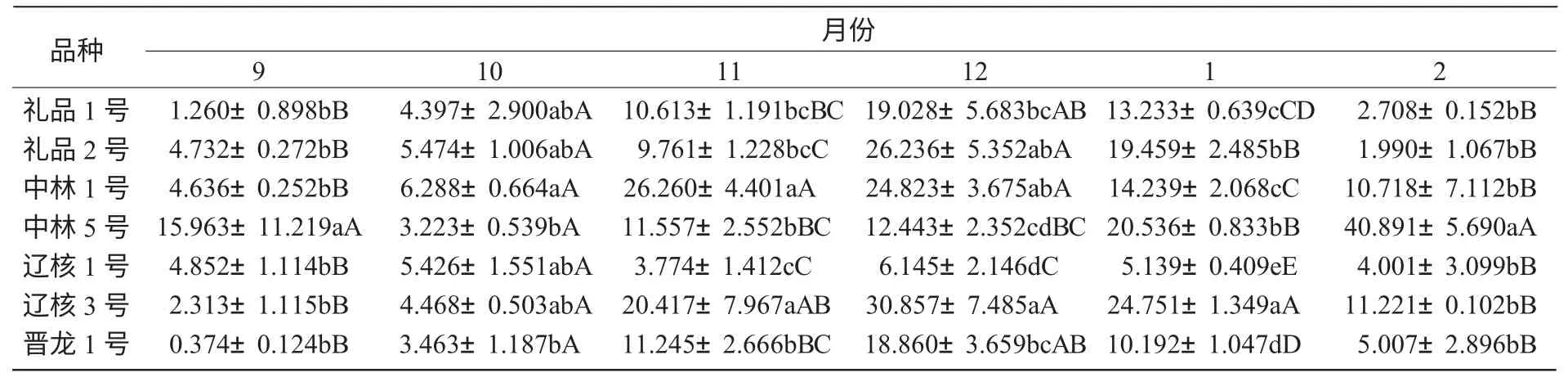
表3 不同品种间脯氨酸含量比较μg/g
2.4 保护酶活性的变化
2.4.1 越冬过程中POD 酶活性变化 由图4 可知,各个核桃品种枝条POD 酶活性随着越冬过程中气温变化及生理发育过程,总体呈初期有小幅下降然后逐渐走高的趋势。在温度较低时,各品种枝条所含的POD 酶活性的变化幅度不大,但随着温度的升高,POD 酶的活性迅速升高。而且到测定后期,各个品种的POD 酶活性都有较大幅度增加。这可能是各个品种在测定后期都已经解除了自然休眠,导致POD 酶活性的大幅变化。气温最低时,中林1 号、礼品2 号、礼品1 号的POD 酶活性较高,而辽核3 号的POD 酶活性相对较低。不同品种间POD 酶活性如表4 所示。


表4 不同品种间POD 酶活性比较U/(g·min)
由表4 可知,各品种枝条POD 酶活性存在显著差异,中林1 号与礼品2 号、晋龙1 号、辽核3 号、中林5 号之间差异极显著。依其枝条POD酶活性的高低,推断其抗寒性强弱顺序依次为:中林1 号>辽核3 号>礼品1 号>礼品2 号>晋龙1 号>辽核1 号>中林5 号。
2.4.2 越冬过程中SOD 酶活性变化 从图5 可以看出,礼品1 号、礼品2 号、中林1 号、辽核3 号、晋龙1 号在整个越冬过程中呈降—升—降的变化趋势,而中林5 号和辽核1 号呈升—降—升—降的变化趋势。在降温初期,各品种SOD 酶活性均有小幅下降,经过初期的低温锻炼后,进入12 月底时,各品种又有不同程度的升高,其中以辽核3 号的活性最大。到1 月份随着气温回升,SOD 酶活性表现为降低的趋势。在整个越冬过程中,辽核3 号的SOD 活性平均值最高,为243.55 U/(g·min)。

从表5 可以看出,各品种枝条SOD 酶活性存在差异,依其枝条SOD 酶活性的高低,推断其抗寒性强弱顺序依次为:礼品2 号>晋龙1 号>礼品1 号>中林1 号>辽核3 号>中林5 号>辽核1 号。

表5 不同品种间SOD 酶活性比较U/(g·min)
3 结论
本试验结果表明,随着核桃生育进程的继续以及气温的变化,可溶性糖、可溶性蛋白、脯氨酸含量、POD 酶活性在核桃枝条内含量越高,抗寒性越强,而SOD 酶活性越高,其抗寒性越弱。
综合比较各品种核桃枝条抗寒性生理生化指标,依各指标与抗寒性可能的相关性,推测7 个品种的抗寒性强弱可能为:辽核3 号、晋龙1号抗寒性较强,中林1 号、礼品2 号和礼品1 号的抗寒性次之,而中林5 号和辽核1 号的抗寒性较差。
由于条件和时间的限制,本试验只对山西省农业科学院果树研究所核桃园7 个品种的部分核桃枝条抗寒性生理生化指标进行了测定分析,而核桃生产实践中栽培应用的品种和影响果树抗寒性的因素及相关指标非常多,还需进行更深入的研究。
[1]潘瑞炽,董愚得.植物生理学[M].北京:高等教育出版社,1995.
[2]Kataoka,K Sumitomo,T Fudano,et al.Changes in sugar content of Phalaenopsis leaves before floral transition[J].Scientia Horticulturae,2004,102:121-132.
[3]刘祖祺,张石诚.植物抗性生理学[M].北京:中国农业出版社,1994.
[4]陈新华,郭宝林,赵静,等.休眠期内甜樱桃不同品种枝条的抗寒性[J].河北农业大学学报,2009,3(6):37-40.
[5]胡春霞,王丽,汤杰.低温对南果梨的生理生化指标的影响[J].沈阳农业大学学报,2009,40(3):349-352.
[6]周蕴薇.翠南报春抗寒生理生态学研究[D].儋州:华南热带农业大学,2003.
[7]黄义江,王宗清.苹果属果树抗寒性的细胞学鉴定[J].园艺学报,1982,9(3):23-30.
[8]邹喻苹.葡萄抗寒生理特性的分析[J].植物学集刊,1989,1(1):64-68.
[9]周碧燕,梁立峰,黄辉白,等.低温和多效唑对香蕉及大蕉超氧化物歧化酶和脱落酸的影响[J].园艺学报,1995,22(4):331-335.
[10]王勇,田建保,程恩明,等.核桃枝条电解质渗出率与抗寒性的关系[J].江西农业大学报,2007(2):230-233.
[11]王丽雪,李荣富,张富仁,等.葡萄枝条中蛋白质、过氧化物酶活性变化与抗寒性的关系[J].内蒙古农牧学院学报,1996,17(1):45-50.
[12]石雪晖,陈祖玉,杨会卿.低温胁迫对柑橘离体叶片中SOD及其同工酶活性的影响 [J]. 园艺学报,1996,23(4):384-386.
[13]王华,王飞,陈登文,等.低温胁迫对杏花SOD 活性和膜脂过氧化的影响[J].果树科学,2000,17(3):197-201.
[14]吴经柔.应用过氧化物酶同Ⅰ酶谱测定苹果的抗寒性[J].果树科学,1990,7(1):41-44.
[15]沙广利,郭长城,睢薇,等.梨抗寒性遗传的研究[J].果树科学,1996,13(3):167-170.
[16]金万梅,董静,尹淑萍,等.冷诱导转录因子CBF1 转化草莓及其抗寒性鉴定 [J]. 西北植物学报,2007,27(2):223-227.
[17]乔富廉.植物生理学实验分析测定技术[M].北京:高等教育出版社,2000.
[18]邹崎.植物生理学实验指导 [M].北京:中国农业出版社,2000.
[19]李合生,孙群,赵世杰.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
