海南优势海草营养元素的含量及其影响因素研究
2012-12-20蔚枝沁吴可为
蔚枝沁, 邓 泓-3, 吴可为, 杜 璟, 马 敏
(1.华东师范大学 环境科学系,上海 200062;2.华东师范大学 上海市城市化生态过程与生态恢复重点实验室,上海 200062;3.华东师范大学 天童森林生态系统国家野外科学观测研究站,上海 200062)
海南优势海草营养元素的含量及其影响因素研究
蔚枝沁1, 邓 泓1-3, 吴可为1, 杜 璟1, 马 敏1
(1.华东师范大学 环境科学系,上海 200062;2.华东师范大学 上海市城市化生态过程与生态恢复重点实验室,上海 200062;3.华东师范大学 天童森林生态系统国家野外科学观测研究站,上海 200062)
研究了海南新村港、黎安港、文昌和琼海(潭门)等地的优势海草种类海菖蒲(Enhalus acoroides)、海神草(Cymodocea rotundata)及泰来藻(Thalassia hemperichil)中氮、磷及铁的含量和影响因素.研究结果表明,海南潮间带海草对养分的吸收与海水营养水平及沉积物氮、磷、有机碳、铁有关.相对于氮,海草叶片磷含量受海水和沉积物的影响更明显.海水磷、沉积物有机碳和铁均能促进海草组织对磷的吸收.海草叶片的氮磷比为新村港(21.73)>黎安(14.25)>文昌(9.53)≈潭门(10.29),与海水中溶解性总氮和溶解性无机磷的比值呈线性正相关,说明海水的富营养化影响了海草叶片的氮磷平衡.
海草; 氮磷比; 铁; 富营养化
0 引 言
氮磷是植物生长的重要元素,通过分析植物体内的氮磷含量及比例,可以反映环境中养分的供应状况、植物的生长速率和群落的生产力水平.在个体水平上,叶片N∶P比率与植物的生长速率成反比[1].在生态系统水平上,植物组织中氮磷的丰富程度及比例可通过食物链向下一个营养级传递,对生态系统的功能具有重要作用[2].海草(seagrass)是生活于热带和温带海域浅水中的单子叶高等植物,主要分布在河口、海湾潮间带湿地和潮下带浅水中,是许多海洋动物的栖息地、生存场和食物来源[3,4].生长于潮间带湿地的海草,在潮汐的作用下,周期性淹没于海水,不仅通过根系从沉积物吸收养分,还能通过叶片直接摄取海水中的营养物质,是海岸生态系统养分循环的重要推动力[5].受人类活动的影响,全球近海环境中的海草床不断退化[6],面积不断缩小[7],引起了各国对海草床保护和恢复研究的重视.我国的海草种类主要分布在华南沿海地区,除人为挖掘对生境的物理破坏(如围海养虾、海水养殖和拖网挖螺)外,水体富营养化是目前海草床受威胁的最重要因素,主要与浮游藻类的大量繁殖造成溶解氧下降和海草可利用光降低有关[8,9].另外,海水富营养化改变了海草对氮磷等养分的吸收及植物组织中的养分平衡,也可能是海草床退化的原因之一[10,11].因此,研究不同营养条件下海草吸收养分的影响因素对揭示海草床生态系统的物质循环具有重要的意义.
本文研究了海南岛近岸潮间带优势海草种类海菖蒲(Enhalus acoroides)、泰来藻(Thalassia hemperichii)及海神草(Cymodocea rotundata)在不同营养条件下氮、磷、铁等营养物质的积累、分布及化学计量特征,分析影响海草养分含量的环境因素(海水和沉积物),为海草床生态学研究和保护提供参考依据.
1 研究方法
1.1 研究区域概况
根据黄小平[8]等对海南海草床分布的研究以及作者的实地调查,综合考虑人类活动的强度和海水的污染状况,沿海南岛东岸从北向南分别选择文昌、琼海和陵水为研究地点(见图1).其中文昌的经济以农业和工业为主;琼海是海南主要旅游景区之一,以农业和旅游业为主;陵水的经济主要依靠旅游业和海水养殖.在研究地点各选择1~2处海草床进行采样,其中文昌的海草床位于椰林镇附近的海滩,周边有大面积的鱼塘、虾塘和菜地,所调查的海草床面积约为100 m×500 m,优势种类为泰来藻和海神草,另有少量针叶藻和喜盐草,底质类型为砂粒.琼海的采样地点位于潭门港,所调查的海草床面积为250 m×50 m,海水较混浊,海草分布稀疏,呈小斑块状,斑块间距5~10 m,优势种为泰来藻和海菖蒲.陵水县拥有目前海南面积最大、保护最为完整的海草床,即新村港和黎安港海草床,面积分别为200 hm2和320 hm2[8],底质类型为沙—泥质.由于面积较大,两处海草床各选一个采样点,其中新村港样点位于猴山旧码头附近,是海南重要的旅游景点之一,居民、载客游船和餐饮渔排密集,近岸码头水面上漂浮有生活垃圾,浮游藻类数量较多,水体较浑浊.海草床离码头约1 km,海草长势良好,呈连续分布,优势种类为泰来藻和海神草,其次为海菖蒲.调查中还发现新村港海草床有若干排水管,可能是附近居民的污水管道或雨水管.较之新村港,黎安港地理位置较偏僻,经济也欠发达,当地居民以出海捕鱼和海水养殖麒麟菜为主.黎安港采样点位于黎安镇的黎明村西滩,海草优势种为海菖蒲,其次为泰来藻,并混生有少量海神草.该样点受人为干扰少,其附近的居民点离海滩为1~2 km,其间有自然分布的灌丛作为缓冲带,海水清澈,海草长势良好.

图1 海南海草床采样点示意图(图中星号标记)Fig.1 Locations of sampling sites(shown as stars)for seagrass beds in Hainan
1.2 样品的采集和处理
2008年4月对海南海草床进行了初步调查,沉积物以中粗砂粒为主,氧化还原电位为-96~210 mV,均处于还原状态;pH为7.83~9.33,偏碱性,EC为5.7~19 s/cm(平均为14.0s/cm),可溶性盐(TDS)含量为0.407%~0.992%.2008年7月,根据潮汐的涨落情况,选择各样点大潮的低潮位时段(此时海草露出海面,便于采样),采集潮间带的海草、沉积物及海水样品.
由于新村港和黎安港海草床面积大且分布连续,对这两处的海草进行了较为详细的采样.共设置4个采样断面,其中新村港3个(每个断面间隔1 km以上),黎安港由于水体较深,仅在近岸处设置了1个断面.每个断面沿潮水涨落方向依次设置3条采样线,每条样线间距为50 m,按当地潮水的实际情况,分别代表高、中、低潮位(其中低潮位的海草在采样时均淹没于水下),沿每条样线等距离取3个采样点(每个点间隔50 m).每个采样点在5 m×5 m的范围内以多点混合法取一个混合底质样及其范围内的植物样品,每个断面共取9个底质样,植物样品的数量根据采样点中物种是否出现而有差异.潭门和文昌采样地的海草床由于面积较小,因此这些地点根据海草的实际分布随机取3个样点,同上述方法采集植物和相应的底质.底质以内径7.5 cm的PVC管取样,取样深度为0~10 cm(为海草根系分布的平均深度).考虑潮汐对海水的混合作用,每个样点各取1份近岸水样分析海草生长的水质环境.
1.3 样品的处理和化学分析
1.3.1 沉积物
采集的沉积物去除贝壳、大块石砾及动植物残体后,风干.研磨过60目筛,用于分析总氮、总磷及有机质;研磨过100目筛,用于分析Fe.过筛的沉积物经HCl-HNO3-HClO4消解后原子吸收光谱仪(PE-AA800)测定Fe[12];标准凯氏法消解后用Skalar流动注射仪测定氮磷[13];重铬酸钾—硫酸外加热法测定有机质[13].
1.3.2 植物样品
本研究采集了3种优势海草种类,即泰来藻、海神草及海菖蒲.除新村港和黎安外,其余地点仅采集到其中的两种(文昌为海神草和泰来藻;潭门为海菖蒲和泰来藻).采集的海草样品用保温箱低温保存,带回实验室后立刻用自来水冲净泥沙和叶片上附生藻类,并用蒸馏水冲洗干净.植物样品分为地上部分(包括茎叶)及地下部分(包括根和根茎),105℃杀青15 min,然后于80℃烘干至恒重并粉碎过60目筛.经HNO3-HClO4消解后用原子吸收光谱仪(PE-AA800)测定Fe[12];标准凯氏法消解后用Skalar流动注射仪测定氮磷[13].
1.3.3 水样
海水样品用聚乙烯瓶采集后用浓硫酸酸化,保存于低温箱中,运回实验室后经0.45μm醋酸纤维滤膜过滤,用于分析溶解性总氮(DTN)、溶解性总磷(DTP)和溶解性无机磷(P;DIP).DTN采用碱性过硫酸钾氧化法,DTP用过硫酸钾氧化法.氮、磷的测定采用Skalar流动注射仪测定.
1.4.4 数据分析
用SPSS 18.0对所测数据进行统计分析,采用One-Way ANOVA比较植物体内养分含量及叶片氮磷比的差异,用t-test分析沉积物及植物养分含量的季节差异.简单线性回归分析海草养分吸收与沉积物性质的相关性及叶片氮磷比与海水氮磷比的相关性.
2 结果与分析
2.1 海草床海水氮磷含量
采样期间海草床的海水氮磷含量见表1.结果表明黎安在各研究地点中营养水平最低,与当地较低的经济发展程度有关;新村港受旅游和网箱养殖的影响,氮的水平偏高;而文昌和潭门海水中氮磷的含量均最高.其中文昌采样点由于受周边农业用地的影响,海水溶解性无机磷(DIP)已经超过海水质量3类标准(GB3097—1997).水质的这种变化趋势反映了当地的经济发展和人类活动对海水富营养化的影响.
2.2 海草床沉积物化学性质
海草床沉积物的化学性质也显示出受人类活动的影响(见表2).富营养化程度最高的文昌和潭门,其沉积物中氮磷及铁的含量也最高.
mg·L-1
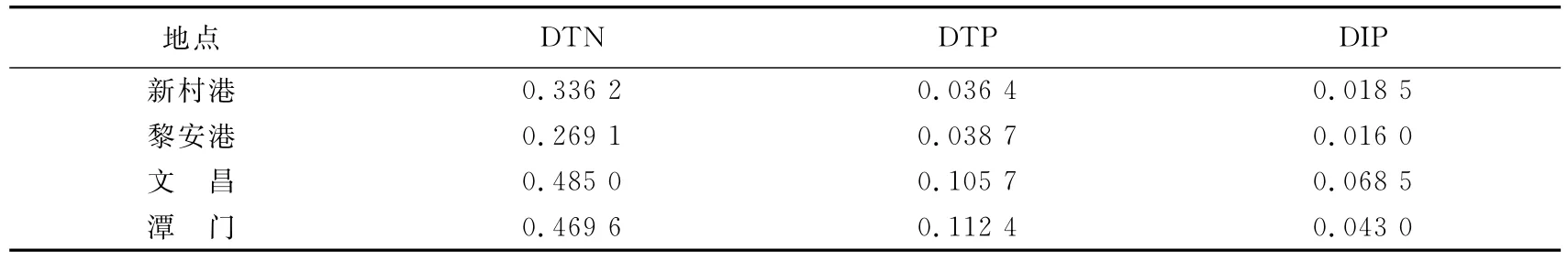
表1 各海草床海水溶解性总氮(DTN)、溶解性总磷(DTP)及溶解性无机磷(DIP)含量Tab.1 Concentrations of dissolved total N (DTN),total P(DTP)and inorganic P(DIP)in seawater
g·kg-1

表2 海南海草床沉积物氮、磷、有机碳及铁含量(Mean±SD)Tab.2 Contents of TN,TP,organic carbon and Fe in sediment of seagrass bed in Hainan
2.3 海草组织的养分含量
海南主要优势海草种类的养分含量见表3.海草对养分的吸收因组织、物种及地点而异.泰来藻、海神草及海菖蒲对氮素和磷素的吸收为地上部分(包括叶和茎)大于地下部分(包括根及根茎),而铁(除海神草外)则主要集中在地下部分.比较物种间的差异发现,海菖蒲地上部分氮和磷的浓度最高,泰来藻最低.同一物种在不同地点养分含量也明显不同.文昌和潭门等地的海草通常具有较高水平的磷或铁,如潭门的海菖蒲地上部分磷含量超过3.0 g/kg,比同时期生长在新村港、黎安港海菖蒲高出230%和36%,潭门和文昌植物体内的铁含量为新村和黎安同种海草的2~3倍.另一方面,新村港和黎安的海草则具有较高浓度的氮,海神草和海菖蒲氮的含量最高可达28~30 g/kg.
海草对养分的富集还与海草的个体大小有关,小型海草如海神草和泰来藻对铁的吸收能力明显高于大型种类海菖蒲.
叶片作为海草的光合器官,其中的氮磷比也具有明显的空间分异(见表4).在各样点中,新村港的海草叶片具有最高的氮磷比(均值为21.73),潭门和文昌最低(均值分别为10.29和9.95),黎安则接近14.
2.4 海水及沉积物对海草养分含量的影响
如前文所述,同种海草在不同地点对养分的吸收存在明显差异,而各样点的海水及沉积物中营养水平也各异,本文进一步分析了影响海草养分积累的环境因素.
2.4.1 海水的影响
通过比较各样点海草组织养分的平均含量,发现海草叶片中磷的含量受海水磷浓度作用明显.文昌和潭门海水中溶解性总磷和无机磷的浓度最高,相应的海草叶片磷的均值为2.17~2.45 g/kg,显著高于新村港和黎安(平均为1.20~1.87 g/kg).尽管文昌海水中氮浓度也最高,海草叶片氮水平(平均20.1 g/kg)却明显低于新村港(平均24.6 g/kg)和黎安(平均26.2 g/kg),说明海水中的氮不是影响叶片氮含量的主要因素.进一步分析发现各样点海草叶片的氮磷比与海水溶解性总氮与溶解性无机磷的比值(DTN∶DIP)有非常显著的相关性(R2=0.750 3)(见图2),说明海水中氮磷营养元素的比例影响了海草光合组织的养分平衡.
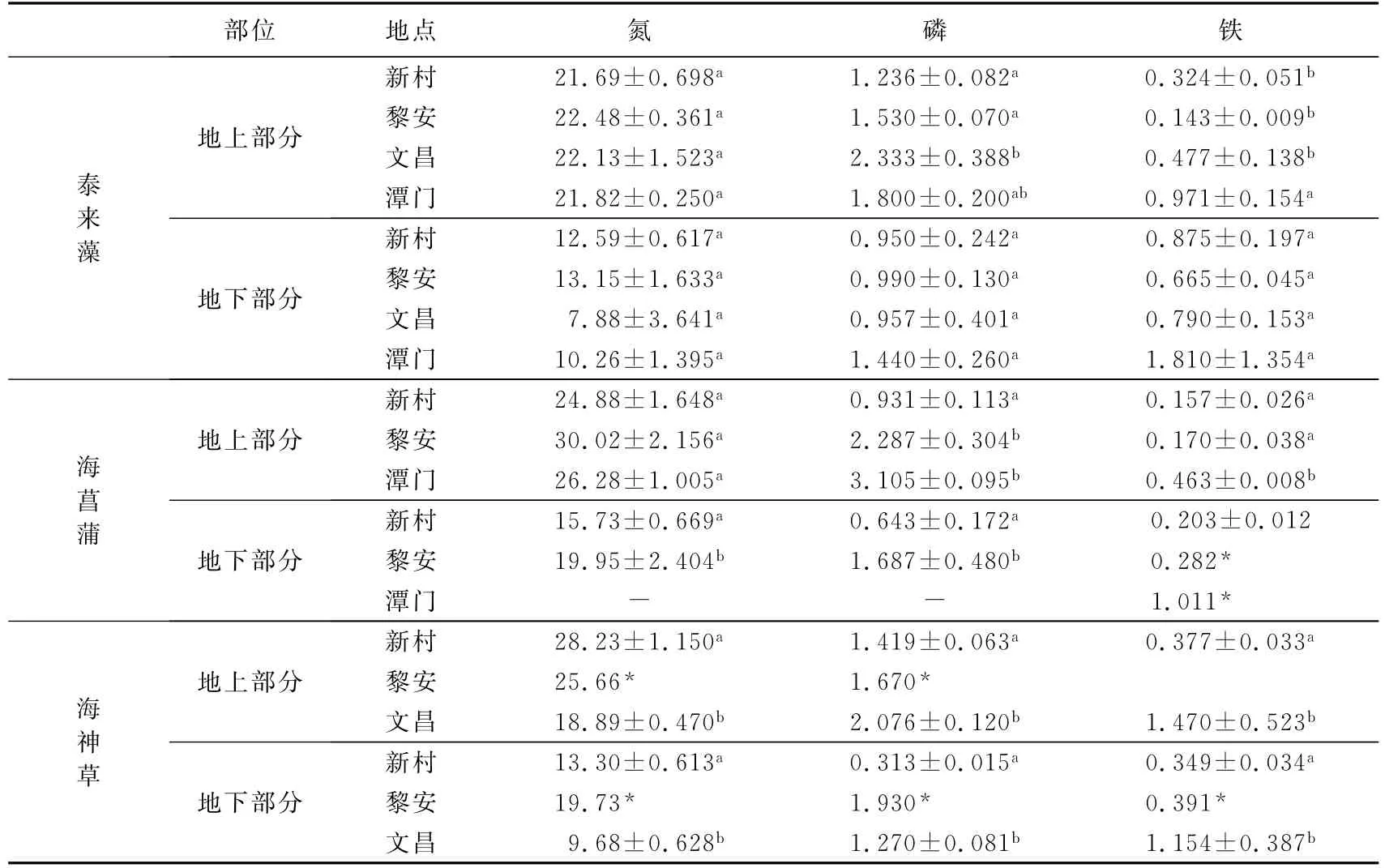
表3 海南优势海草种类体内氮、磷、铁含量(Mean±SD)Tab.3 Nutrient accumulation by dominant seagrass species in Hainan g·kg-1

表4 海南优势海草种类叶片的氮磷比(Mean±SD)Tab.4 Ratio of nitrogen to phosphorus in dominant seagrass species in Hainan
2.4.2 与沉积物的相关性
相关性分析表明,海草对营养元素的吸收不同程度地受到沉积物理化性质的作用(见表5).泰来藻地上部分的氮含量与沉积物中的氮水平显著正相关,但在海菖蒲和海神草中则没有相关性.海草地下部分对氮磷的吸收与沉积物有机碳水平明显正相关.铁的吸收也与沉积物铁具有非常显著的相关性.研究还发现,沉积物中的铁和磷对海草铁、磷素的吸收具有非常强烈的交互作用.例如,沉积物铁促进了泰来藻和海菖蒲地上部分及海神草地下部分对磷素的吸收.另一方面,沉积物中的磷也明显促进了海菖蒲(地上部分和地下部分)及海神草(地上部分)对铁的吸收.沉积物对海草养分吸收的影响还表现在叶片氮磷比方面,研究发现沉积物有机碳和铁均与叶片氮磷比显著负相关,这与两者均对磷素吸收有促进作用有关.沉积物对海草养分吸收的影响程度还与植物大小有关,大型的海菖蒲较小型的泰来藻和海神草受沉积物影响更明显,尤其是地上部分(见表5).这说明海菖蒲对养分的吸收更多地来源于沉积物,而泰来藻和海神草的养分则主要来源于水体,与物种不同的生态策略有关.海菖蒲生长缓慢,其发达的根系有利于对沉积物养分的吸收,而根系浅的泰来藻和海神草则通过叶片对海水养分的吸收维持其快速生长.
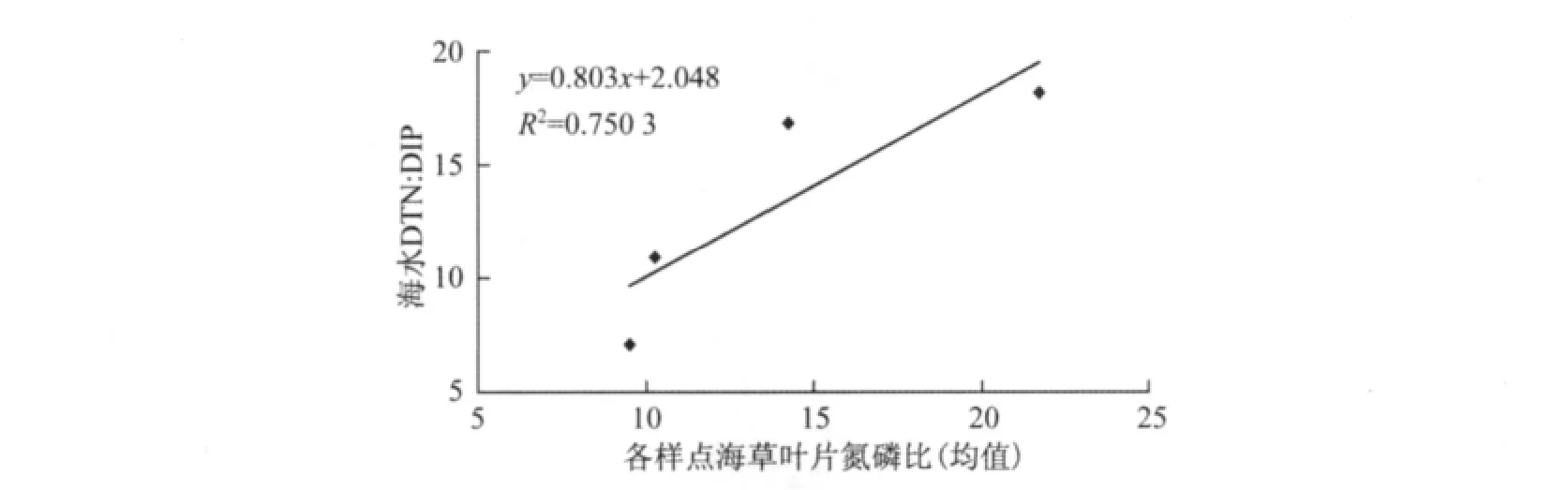
图2 各样点海草叶片氮磷比均值与海水DTN∶DIP的相关性Fig.2 Correlation between average leaf N∶P and DTN∶DIP of seawater in study sites

表5 海南海草优势种体内养分含量与沉积物理化性质的Pearson相关系数Tab.5 Pearson correlation between nutrient uptake by seagrasses and sediment characteristics
3 讨 论
3.1 海草叶片的氮、磷含量及化学计量比
叶片作为主要的光合作用器官,其养分含量是评价植物生长状况的重要依据.7月份是热带海草生长的旺盛季节,本研究中海草叶片氮含量为16.8~33.4 g/kg,平均为23.98 g/kg,磷的含量为0.47~3.2 g/kg,平均为1.62 g/kg,与Florida湾 Thalassia testudinum 比较接近(氮含量为8.8~39.6 g/kg,磷含量为0.48~2.43 g/kg)[17].以往的研究表明,热带和亚热带海草生长通常受氮、磷和铁的限制[6,14-16].Duarte在广泛研究了27种分布于不同地理区域的海草养分含量的基础上得出海草叶片中氮、磷的中位值分别为18和2.0g/kg,并以此判断海草的生长是否受营养限制[18].按照这一标准,本研究中新村港海草磷较缺乏,而文昌的海草则缺氮素.类似的结论也可以通过比较叶片氮磷比得出.叶片氮磷比能反映植物的生境状况,并能了解植物生长受养分的限制情况[1,2].Atkinson和Smith认为养分平衡时,海草叶片N∶P摩尔比为30[19],换算成质量比即为14,高于此值,生长受磷限制,反之则受氮限制.本研究结果表明,海草生长旺盛期(7月)的叶片氮磷比,物种间和各样点间均有显著性差异(见表4).叶片氮磷比的变化规律为新村港(21.73)>黎安(14.25)>潭门(10.29)≈文昌(10.29),说明新村港的海草生长受磷限制,潭门和文昌受氮限制,而黎安的海草则处于养分平衡状态.这一变化规律也反映了各样点的经济发展状况或人类活动强度对环境的影响.黎安港经济发展程度(国内生产总值GDP)最低[20],环境质量的数据也表明黎安是海南水质最为清洁的区域之一[8],本研究中测得的海水氮、磷含量也为最低.新村港是海南重要旅游景点之一,有较为密集的网箱养殖和设在渔排上接待游客的餐饮业,其海水氮、磷相对黎安有增加的趋势,尤其是氮,比黎安高25%(见表1).其他研究也有类似的结果,如新村港网箱养鱼区水体中氮负荷偏高,磷则处于较低水平[10].因此新村港海草对氮的吸收能力提高,叶片N∶P比大于14.文昌和潭门人口密集,经济发展程度较高,近海水体除氮素外,磷的含量也逐年增加(2009年海南省海洋环境状况公报),为黎安的2~4倍,使海草对磷的吸收超过氮.尽管文昌海水及沉积物中氮的水平很高,但海草却仍表现出氮缺乏,叶片N∶P比降低.各样点海草叶片氮磷比的均值与海水中溶解性总氮和溶解性无机磷的比值呈线性相关(见图2),则进一步说明人类活动引起的海水富营养化会最终影响海草对养分的吸收平衡.海草是众多海洋动物的食物来源,海草组织中氮磷比的变化会通过食物链影响下一个营养级对氮磷的吸收和代谢[1,2],最终影响海草床生态系统的物种组成和养分循环.
如前所述,新村港海草生长受磷限制,在各海草种类中又以海菖蒲最为明显(叶片N∶P最高,为28.19±2.09).这可能与两方面的原因有关.首先,海菖蒲是最大型的海草种类,生长缓慢,生产力在所研究的3种海草中最低[8],因此相对于生产力高的泰来藻和海神草,其对养分的竞争能力较弱.其次,本研究开展的时间为7月份,时值海菖蒲的花期,植物开花需要更多的营养物质,因此更容易出现养分缺乏的现象.新村港在地形上属于泻湖,与外海的水体交换能力弱,营养及其他污染物进入水体后滞留时间长,容易累积,若不采取有效措施,水体富营养化程度将继续增加,对海菖蒲的生长将是非常严重的威胁.
3.2 影响海草养分含量的环境因素
海草的生长依赖于环境中养分的供给.由于生活在水下,海草可通过叶片和根系吸收营养物质.在海洋生态系统中,沉积物中的养分水平远远高于海水[6],因此在没有外源营养输入的情况下,沉积物是海草养分的主要来源.在近岸系统中,由于受到人类活动的影响,海水中的氮磷有不同程度的提高,甚至产生富营养化.本研究中,海南近岸潮间带的海草对养分的积累受到海水富营养化的影响.其中磷的增加提高了海草叶片磷的含量,而氮的增加对叶片的氮含量没有明显的促进作用,说明磷相对于氮而言,可能是影响海草生长更为重要的限制因素.另一方面,沉积物主要影响海草地下部分对营养的吸收.除了氮磷的直接作用外,沉积物的有机碳和铁还能间接地影响海草的养分水平.本研究表明沉积物有机碳促进了海草地上/地下组织对氮、磷或铁的吸收.这主要是因为有机碳的增加为根际微生物提供碳源,提高了微生物数量和多样性[22],促进氮、磷、铁的转化和利用.
铁是植物合成叶绿素所必需的元素.在热带海草床,沉积物中Fe的含量一般小于200 mg/kg[21],且大多数被碳酸盐束缚而不能被海草利用,加上缺少陆源铁离子的输入,海草生长很容易受到铁限制[22].被子植物维持生长时叶片所需Fe含量最低应达到100 mg/kg,否则就会导致萎黄病等缺铁病害[23].本文所研究的海草铁含量(均值143.3~1 470 mg/kg)虽然低于一般的淡水植物(3 170 mg/kg)[24],但均高于叶片的缺铁阈值,因此铁未对海南近岸海草生长形成限制.海草组织铁含量与沉积物氮磷及铁水平有关(见表5).沉积物中的铁还能通过间接的作用促进海草的生长.例如铁的添加能缓解硫酸盐还原产生的硫化物对海草的危害,提高叶片碱性磷酸酶的活性,促进对磷素的利用效率[25].本研究则发现沉积物铁促进了泰来藻(地上部分)、海神草(地下部分)及海菖蒲(地上、地下)磷含量的增加(见表5),原因可能是铁影响沉积物中磷素的化学形态[26],并通过与植物的作用如在根表形成铁氧化物膜促进磷素的吸收[27].从沉积物铁含量来看,经济发展程度较高的文昌、潭门明显高于新村港和黎安.这与本文所研究的海草床处于潮间带,受陆源输入影响有关.如人口密集区城市化水平较高,不透水面的增加提高了地表径流,挟带大量富含铁的土壤进入海岸带,使沉积物铁的含量升高.文昌和潭门的海水同时还有较高含量的磷,因此本研究中观察到沉积物铁与叶片氮磷比呈负相关(见表5)的现象.近岸潮间带在人类活动干扰下,富营养化和铁的交互作用对海草床有何影响值得进一步开展研究.
4 结 论
(1)海南潮间带海草对养分的吸收受海水营养水平及沉积物氮、磷、有机碳及铁的影响.相对于氮,海草磷含量受海水和沉积物的作用更为明显.海水中磷、沉积物有机碳和铁对磷均能促进海草组织对磷的吸收.
(2)本文所研究的海草铁含量(均值143.3~1 470 mg/kg)均高于叶片的缺铁阈值,因此铁未对海南近岸海草生长形成限制.沉积物氮磷及铁均能促进海草组织对铁的吸收.
(3)人类活动在不同程度上提高了海水营养盐浓度,影响了海草叶片的氮磷比.海草叶片的氮磷比为新村港(21.73)>黎安(14.25)>文昌(9.53)≈潭门(10.29),与海水中溶解性总氮与溶解性无机磷的比值呈线性正相关,说明海水的富营养化影响了海草叶片的氮磷平衡.
[1] STNER R W,ELSER J J.Ecological Stoichiometry:the Biology of Elements from Molecules to the Biosphere[M].Princeton:Princeton University Press,2002.
[2] ANDERSON T R,BOERSMA M,RAUBENHEIMER D.Stoichiometry:linking elements to biochemicals[J].E-cology,2004,85:1193-1202.
[3] HECK K.L J R,THORMAN T A.The nursery role of seagrass meadows in the upper and lower reaches of the Chesapeake Bay[J].Estuaries,1984(7):70-92.
[4] BOSTROM C,BONSDORFF E.Zoobenthic community establishment and habitat complexity—the importance of seagrass shoot-density,morphology and physical disturbance for faunal recruitment[J].Marine Ecology Progress Series,2000,205:23-138.
[5] JONES C G,LAWTON J H,SHACHAK M.Positive and negative effects of organisms as physical ecosystem engineers.Ecology[J].1997,78:1946-1957.
[6] UDY J W,DENNISON W C,LONG W J L.Responses of seagrass to nutrients in the great barrier reef[J].Australia Marine Ecology Progress Series,1999,185:257-271.
[7] WAYCOTT M,DUARTE C M,CARRUTHERS T J B.Accelerating loss of seagrasses across the globe threatens coastal ecosystems[J].Proceedings of the National Academy of Sciences,2009,106(30):12377-12381.
[8] 黄小平,黄良民,李颖虹.华南沿海主要海草床及其生境威胁[J].科学通报,2006,增刊Ⅱ,51:114-119.
[9] 范航清,彭胜,石雅君,等.广西北部湾沿海海草资源与研究状况[J].广西科学,2007,14(3):289-295.
[10] 黄道建,黄小平,黄良民.海南岛新村湾营养负荷对海菖蒲的影响研究[J].海洋科学进展,2007,25(2):200-207.
[11] 黄道建,黄小平,黄正光.海南新村湾海菖蒲TN和TP含量时空变化及其对营养负荷的响应[J].海洋环境科学,2010,29(1):40-43.
[12] PAGE A L,MILLER R H,KEENEY D R.Methods of Soil Analysis-Chemical and Microbiological Properties[M].Wisconsin:Madison,1982.
[13] 鲍士旦,土壤农化分析[M].北京:中国农业出版社,2005.
[14] LONG M H,MCGLATHERY K J,ZIEMAN J C,et al.The role of organic acid exudates in liberating phosphorus from seagrass-vegetated carbonate sediments[J].Limnology Oceanogry,2008,53:2616-2626.
[15] DUARTE C M,MERINO M,GALLEGOS M.Evidence of iron deficiency in seagrasses growing above carbonate sediments[J].Limnol Oceanogr,1995,40:1153-1158.
[16] HABEEBREHMAN H,PRABHAKARAN M P,JACOB J,et al.Variability in biological responses influenced by upwelling events in the Eastern Arabian Sea[J].Journal of Marine System,2008,74:545-560.
[17] FOURQUREAN J W,ZIEMAN J C.Nutrient content of the seagrass Thalassia testdinumreveals regional patterns of relative availability of nitrogen and phosphrous in the Florida Keys USA[J].Biogeochemistry,2002,61:229-245.
[18] DUARTE C M,CHISCANO C L.Seagrass biomass and production:A reassessment[J].Aquatic Botany,1999,65:159-174.
[19] ATKINSON M J,SMITH S V.C∶N∶P ratios of benthic marine plants[J].Limnology Oceanogry,1983,28:568-574.
[20] 海南省统计局,国家统计局海南调查总队.海南统计年鉴[M].北京:中国统计出版社,2009.
[21] KAREN J M,PETER B,ROXANNE M.Using porewater profiles to assess nutrient availability in seagrass-vegetated carbonate sediments[J].Biogeochemistry,2001,56:239-263.
[22] SERGIO R,STEPHEN A M,JAMES W F.The effects of manipulation of sedimentary iron and organic matter on sediment biogeochemistry and seagrasses in a subtropical carbonate environment[J].Biogeochemistry,2008,87:113-126.
[23] DUARTE C M,MERINO M,GALLEGOS M.Evidence of iron deficiency in seagrasses growing above carbonate sediments[J].Limnol Oceanogr,1995,40:1153-1158.
[24] HUTCHINSON G E.A treatyise on Limnology[M]//Limnological Botany.New York:John Wiley and Sons Press,1975(3).
[25] HOLMER M,DUARTE C M,MARBA N.Iron additions reduce sulfate reduction rates and improve seagrass growth on organic-enriched carbonate sediments[J].Ecosystems,2005,8(6):721-730.
[26] MONBET P,MCKELVIE I D,WORSFOLD P J.Phosphorus speciation,burial and regeneration in coastal lagoon sediments of the Gippsland lakes(Victoria,Australia)[J].Environmental Chemistry,2007(4):334-346.
[27] CROWDER A A,ST-CYR L.Iron oxide plaque on wetland roots[J].Trends in Soil Science,1991(1):315-329.
Nutrient contents of dominant seagrass species and their affecting factors in Hainan Province
YU Zhi-qin1, DENG Hong1-3, WU Ke-wei1, DU Jing1, MA Min
(1.Department of Environment Science,East China Normal University,Shanghai 200062,China;2.Shanghai Key Lab for Urban Ecological Processes and Eco-Restoration,East China Normal University,Shanghai 200062,China;3.Tiantong National Station of Forest Ecosystem,Chinese National Ecosystem Observation and Research Network,East China Normal University,Shanghai 200062,China)
Nutrient(N,P and Fe)contents of Enhalus acoroides,Cymodocea rotundata and Thalassia hemperichi dominated in the seagrass beds in Hainan were studied in this paper.The results indicated that nutrient uptake by inter-tidal seagrass was influenced by both N,P in seawater and chemical characteristics of sediments(N,P,TOC and Fe).Phosphorus concentration of seagrass,which was more sensitive to water and sediment than that of nitrogen,was positively correlated with the level of P in seawater and TOC and total Fe in sediment.Enhanced nutrientlevel induced by human activates has exerted imbalance of N and P uptake in seagrass,with the leaf N:P of Xincungang(21.73)>Li′an(14.25)> Wenchang(9.53)≈ Tanmen(10.29).The fact that leaf N:P had strong correlation with DTN:DIP in seawater has indicated the influence of eutrophication on the imbalance of nutrient uptake in seagrass.
seagrass; ratio of nitrogen to phosphorus; iron; eutrophication
Q948
A
10.3969/j.issn.1000-5641.2012.04.016
1000-5641(2012)04-0131-11
2011-04
科技部863项目子项目(2007AA09Z432);教育部博士点新教师基金(200802691023)
蔚枝沁,女,硕士研究生.
邓泓,女,副教授,主要从事污染生态学和生态恢复的研究及教学工作.E-mail:lilac_deng@yahoo.com.cn.
