体外产气法评定植酸酶对稻糠、玉米麸和麦麸瘤胃发酵特性的影响
2012-12-20沈莎莎杨红建任清长白萨茹拉张晓明
沈莎莎 杨红建 任清长 白萨茹拉 张晓明
(中国农业大学动物科学技术学院,动物营养学国家重点实验室,北京 100193)
植酸酶(phytase)即六磷酸肌醇水解酶(myoinositol hexakisphosphate phosphohydrolase),是通过水解植酸的磷酸单酯键生成低级磷酸肌醇和无机磷的一类酶的总称[1]。目前,植酸酶作为一种新型的饲料添加剂,通常添加到单胃动物或鱼类的饲粮中用来降解植物性饲料中的抗营养因子——植酸,并释放出无机磷以及与植酸结合的蛋白质、微量元素等,进而提高饲料磷利用效率和动物生长性能。与单胃动物不同,反刍动物能很大程度上依靠瘤胃微生物分泌的内源性植酸酶来消化利用饲料中的植酸[1]。由于在反刍动物特别是高产奶牛饲粮中富含植酸磷的谷物饲料所占比例很大,盲目额外添加磷酸氢钙等无机磷所带来的奶牛养殖磷排放环境污染问题备受关注。有研究认为在饲粮中额外添加少量的植酸酶可以提高反刍动物对饲料植酸磷的利用率[2]。但也有文献研究认为,瘤胃微生物在降解利用富含纤维饲料的同时可以依靠自身分泌的植酸酶来降解利用饲料中的植酸磷,从而可能无需在饲粮中额外添加植酸酶[3]。因此,作者采用短期人工瘤胃发酵试验结合动态产气实时记录技术,分析了添加植酸酶对瘤胃微生物降解和利用稻糠、玉米麸和麦麸的功效,以期为今后通过开展饲养试验开发和研究植酸酶作为反刍动物饲料添加剂提供依据。
1 材料与方法
1.1 发酵底物
本试验采集不同产地的糠麸饲料作为试验材料,对稻糠分别编号为RB1、RB2、RB3,玉米麸分别编号为 MB1、MB2、MB3,麦麸分别编号为WB1、WB2、WB3。饲料经电热鼓风干燥箱65℃烘干48 h后粉碎过40目筛。并按实验室常规方法[4]对试验饲料进行粗蛋白质(CP)、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)和植酸磷含量的测定。
1.2 植酸酶处理方法
植酸酶样品由中国农业科学院饲料研究所赠送,活性为5 000 U/g(1 U代表该植酸酶在反应条件下作用于植酸钠溶液,每分钟可释放出1μmol无机磷)。参照Engelen等[5]的酶处理方法,称取0.5 g植酸酶溶解于100 mL乙酸缓冲液(称取1.76 g乙酸、30.02 g 三水乙酸钠、0.147 g 二水氯化钙溶解于900 mL蒸馏水中,用乙酸调节pH至5.50,并用蒸馏水定容至1 000 mL),常温下在磁力搅拌器上高速搅拌30 min,制成25 U/mL植酸酶工作液。
1.3 体外发酵
使用本实验室自主开发和设计的64通路AGRS-Ⅲ型体外发酵产气自动记录装置和软件系统[6]进行实时测定累积产气量。
试验前,根据Menke等[7]的配方配制缓冲液,主要包括微量元素溶液、碳酸缓冲溶液、磷酸缓冲溶液、硫化钠(Na2S)还原溶液以及刃天青指示剂。配制好的缓冲液混匀后持续通入CO230 min以上直至溶液pH达到6.8,置于39℃恒温水浴锅中备用。
称取500 mg待测饲料于150 mL厌氧发酵瓶中,每种饲料设置对照组和加酶组。对照组设置3个平行,加酶组设置4个平行。于晨饲前1 h内采集3头安装有永久瘤胃瘘管、平均日产奶量为20 kg的泌乳期成年荷斯坦奶牛的瘤胃液,等体积混匀后经4层纱布过滤,滤液置于39℃恒温水浴锅中,持续通入CO2备用。
用移液器准确移取50 mL缓冲液和25 mL瘤胃液于各发酵瓶中,并用微量移液器将100μL配制好的植酸酶工作液添加至加酶组发酵瓶中,通入CO2数秒,驱除液面上方空气后盖上瓶塞,立即与AGRS-Ⅲ型体外发酵产气自动记录装置各气路通道相连,并于39℃条件下连续培养72 h,期间每12 h轻轻晃动培养基。
1.4 样品收集与处理
连续发酵72 h后,关闭发酵产气记录系统,打开发酵瓶测定pH。将瓶内发酵液倒入经烘干称重的尼龙袋(孔径200目)中过滤,采集滤液样品。尼龙袋连同发酵剩余底物用蒸馏水漂洗后,在65℃下烘干至少48 h后称重,根据发酵前饲料样中的干物质含量,利用差减法计算待测饲料体外发酵干物质消失率(dry matter disappearance in vitro,IVDMD)。取1.0 mL发酵滤液在4℃条件下3 000×g离心5 min,收集上清液。取500μL上清液加入150μL 25%(质量体积比)偏磷酸,混匀,4℃静置30 min后在10 000×g离心15 min,取上清液待分析。
1.5 产气动力学模型分析
根据AGRS-Ⅲ装置自动记录到的各发酵瓶的产气时间和对应的累积产气量,参照Ørskov等[8]提出的指数函数模型对不同饲料累积产气量数据进行非线性拟合,得出:
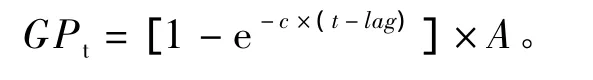
式中:GPt为累积产气量(mL/g,干物质基础),c为产气速率(mL/h),t为产气时间(h),lag为产气延滞时间(h),A为发酵底物在该产气速率下的理论最大产气量(mL)。
根据下式计算达到最大产气量1/2时的产气速率:

式中:AGPR为达到最大产气量1/2时的产气速率(mL/h)。
1.6 氨态氮和挥发性脂肪酸(VFA)的测定
发酵液中氨态氮浓度的测定采用靛酚比色法[9]。VFA 含量的测定参照杨红建等[10]的方法。非糖异生类短链脂肪酸(乙酸、丁酸)与糖异生类短链脂肪酸(丙酸、戊酸等)的比率按下式计算:非糖与糖异生的类短链脂肪酸比率(NGR)=(乙酸 +2×丁酸 +戊酸)/(丙酸 +戊酸)[11]。
1.7 统计分析
试验数据采用Excel 2003整理后,采用SAS 9.1.3统计软件中GLM 过程进行方差分析,采用Duncan氏法和MEANS语句统计组间差异的显著性,并利用LSMENAS语句统计各测定指标最小二乘法均值的标准误(SEM)。差异性显著水平为P <0.050 0,差异极显著水平为 P <0.000 1。
2 结果
2.1 试验发酵底物的常规营养成分
由表1可知,不同来源的同种糠麸饲料营养成分存在较大差异,其中,稻糠中RB1的CP、植酸磷的含量均高于RB2、RB3,其 NDF和ADF的含量最低;麦麸中WB1也存在同样的特性;玉米麸中MB1的CP和植酸磷的含量最高,NDF含量最低,ADF含量在3中饲料中居中。此外,稻糠、玉米麸和麦麸的营养成分也不同。稻糠的CP含量最低,但其NDF和ADF的含量均高于玉米麸和麦麸。
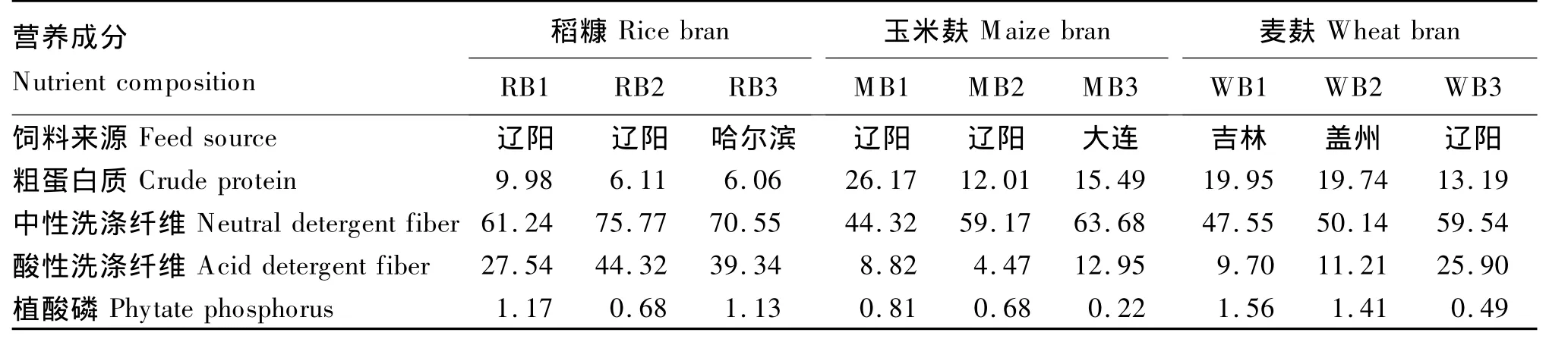
表1 试验发酵底物的常规营养成分(干物质基础)Table 1 Common nutrient composition of the experimental fermentation substrates(DM basis) %
2.2 添加植酸酶对3种糠麸饲料体外发酵参数的影响
由表2可知,玉米麸对照组的IVDMD显著高于加酶组(P<0.050 0),而稻糠和麦麸对照组的IVDMD和加酶组差异均不显著(P>0.050 0)。饲料种类对IVDMD有极显著影响(P<0.000 1),比较3种糠麸饲料发现,它们的IVDMD表现为玉米麸>麦麸>稻糠。而就植酸酶添加效果而言,添加5 U/g的植酸酶并不能显著改变饲料IVDMD(P >0.050 0)。
除稻糠加酶组产气速率显著低于对照组外(P<0.050 0),同一饲料各发酵动力学参数,对照组和加酶组的间均无显著差异(P>0.050 0)。比较稻糠、玉米麸和麦麸发现,饲料种类不同,72 h累积产气量、理论最大产气量及达到最大产气量1/2时的产气速率存在极显著差异(P<0.000 1),产气速率和产气延滞时间存在显著差异(P<0.050 0)。此外,比较植酸酶的添加对饲料发酵动力学参数的影响发现,添加5 U/g植酸酶显著降低了饲料产气速率及达到最大产气量1/2时的产气速率(P<0.050 0)。不同饲料及酶处理的累积产气量动态有明显的不同,总体上保持玉米麸>稻糠>麦麸(图1)。
2.3 添加植酸酶对3种糠麸体外瘤胃发酵特性的影响
由表3可知,对同一饲料而言,对照组和加酶组pH、NH3-N浓度、VFA含量(除异丁酸外)和NGR均未表现出显著差异(P>0.050 0)。植酸酶显著提高了总挥发性脂肪酸(TVFA)浓度(P<0.050 0),且TVFA浓度增幅由高至低依次为麦麸(4.2%)、稻糠(3.3%)、玉米麸(1.7%);从VFA含量来看,植酸酶显著提高了异丁酸含量(P<0.050 0),异戊酸和戊酸的含量有提高的趋势,但差异不显著(P>0.050 0)。此外,添加植酸酶也不能显著改变NGR(P>0.050 0)。
比较稻糠、玉米麸和麦麸3种饲料发现,各种VFA的含量不同饲料间存在较大的差异。丁酸和异戊酸含量饲料间不存在显著差异(P>0.050 0),饲料种类不同,异丁酸含量存在显著差异(P<0.050 0),乙酸、丙酸和戊酸含量均存在极显著差异(P<0.000 1)。乙酸含量稻糠>麦麸>玉米麸,丙酸和戊酸的含量均为玉米麸>麦麸>稻糠。
3 讨论
饲料有机物的消化率与体外培养的产气量之间存在较高的相关性,即产气量越高,饲料在瘤胃内的发酵程度越高[12]。本试验以稻糠、玉米麸和麦麸为发酵底物,采用短期人工瘤胃发酵试验结合动态产气实时记录技术,发现3种糠麸饲料微生物消化率表现为玉米麸 >麦麸 >稻糠,符合Nsahlai等[13]研究指出的理论最大产气量与饲料本身NDF含量呈显著负相关,与CP含量呈正相关的规律。但额外添加植酸酶除可显著降低产气速率外,并未显著影响饲料体外发酵累积产气量和理论最大产气量。目前,国内外有关额外添加植酸酶对体外发酵产气量影响未见报道,研究多集中添加植酸酶提高饲料植酸磷的降解利用效率上。Kincaid等[2]发现在奶牛全混合日粮中额外添加427 U/kg植酸酶能明显提高饲料植酸磷的降解和总磷的利用率,但奶牛的干物质采食量和产奶效率均不受植酸酶的影响。Bravo等[14]在泌乳山羊和干奶期奶牛饲粮中额外添加真菌植酸酶(2 000 IU),并利用尼龙袋技术研究豆粕和菜籽粕的干物质降解率和磷在瘤胃中的降解率时指出,添加真菌植酸酶有降低豆粕干物质降解率的趋势,但对菜籽粕的干物质降解率没有影响。本试验通过比较对照组和加酶组IVDMD发现,在稻糠、玉米麸和麦麸中额外添加5 U/g植酸酶有降低其IVDMD的趋势,且玉米麸IVDMD的变化最显著,与 Bravo 等[14]结果相似。

表2 植酸酶对糠麸饲料体外发酵72 h干物质消失率、产气量及发酵动力学参数的影响Table 2 Effects of phytase on IVDMD,gas production and fermentation kinetic parameters of grain brans after fermented for 72 h

图1 体外发酵累积产气量动态曲线Fig.1 The dynamic curve of accumulative gas production of fermentation in vitro
饲料在瘤胃微生物的作用下可产生大量的乙酸、丙酸及丁酸等VFA,它们是反刍动物的主要能量来源,可提供反刍动物总能量需要量的70%~80%[15]。本试验通过测定发酵液中的各种 VFA含量发现,虽然与对照组相比,额外添加植酸酶并未显著影响3种糠麸饲料发酵产生的乙酸、丙酸和丁酸含量,但从整体来看,乙酸的含量较高,为66.42% ~71.10%;丙酸含量偏低,为 12.26% ~16.35%;丁酸含量也在10%左右。额外添加植酸酶却显著增加了异丁酸的含量,同时也显著提高了饲料发酵后TVFA浓度。该结果与Bravo等[14]的结果稍有不同,该研究通过测定乙酸、丙酸和丁酸3种主要的短链VFA含量指出,在泌乳山羊饲粮中额外添加真菌植酸酶并不能显著影响乙酸含量和乙酸/丙酸,却显著提高了丙酸的含量(从23.4%提高至28.4%)以及显著降低了丁酸的含量(从16.3%降低至12.4%),不过这种 VFA含量的变化可能是因为在泌乳山羊饲粮中添加了70%的精料。戊酸、异丁酸、异戊酸分别是饲料的蛋白质中缬氨酸、亮氨酸、异亮氨酸的微生物发酵产物[16],在本试验中这3种VFA在添加植酸酶后都有不同程度的提高,尤其是异丁酸含量显著升高,从一定程度上说明添加植酸酶促进了瘤胃微生物对饲料中上述3种氨基酸的代谢,在后续的研究中应注意瘤胃微生物蛋白质合成量的测定。此外,本试验根据测得的乙酸、丙酸、丁酸及戊酸含量计算得出的NGR与Zhang等[17]用羊草∶玉米为4∶1的混合饲料底物进行体外瘤胃发酵48 h后测得的未添加任何甲烷抑制剂的对照组NGR(3.83)相比结果偏高,但与 Yang等[18]利用不同碳源和氮源组合的培养基研究新丽鞭毛菌(Neocallimastix sp.YQ1)以苜蓿作为木质纤维性底物时的纤维降解活性试验中测得的 NGR(4.9~6.5)相近。本试验结果表明糠麸饲料在瘤胃微生物发酵作用下生糖前体物——丙酸含量较低,而非生糖前体物或乳脂前体物——乙酸和丁酸含量较高。

表3 植酸酶对糠麸饲料体外瘤胃发酵特性的影响Table 3 Effects of phytase on rumen fermentation characteristics of grain brans in vitro
4 结论
稻糠、玉米麸和麦麸间体外瘤胃发酵特性存在差异,添加5 U/g植酸酶可在一定程度上促进瘤胃微生物发酵释放出更多的VFA,提高瘤胃微生物对饲料中能量物质的发酵利用效率。
[1] 沈莎莎,杨红建,张晓明.瘤胃微生物植酸酶的分类与酶学性质研究进展[J].畜牧兽医学报,2011,42(12):1655-1660.
[2] KINCAID R L,GARIKIPATI D K,NENNICH T D,et al.Effect of grain source and exogenous phytase on phosphorus digestibility in dairy cows[J].Journal of Dairy Science,2005,88(8):2893-2902.
[3] RAUN A,CHENG E,BURROUGHS W,et al.Ruminant nutrition:phytate phosphorus hydrolysis and availability to rumen microorganisms[J].Journal of Agricultural and Food Chemistry,1956,4(10):869-871.
[4] 张丽英.饲料分析及饲料质量检测技术[M].3版.北京:中国农业大学出版社,2007.
[5] ENGELEN A J,VANDERHEEFT F C,RANDSDORP PH G,et al.Simple and rapid determination of phytase activity[J].Journal of AOAC International,1994,77(3):760-764.
[6] 杨红建,宋正河,祝仕平,等.一种发酵微量气体产生量数据自动采集存储装置及方法:中国,ZL200610011301.X[P].2007-12-19.
[7] MENKE K H,STEINGASSH.Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid[J].Animal Research Development,1988,28:55.
[8] ØRSKOV E R,MCDONALD I.The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage[J].Journal of Agricultural Science,1979,92:499-503.
[9] VERDOUW H,VAN ECHTELD C JA,DEKKERSE M J.Ammonia determination based on indophenol formation with sodium salicylate[J].Water Research,1978,12(6):399-402.
[10] 杨红建,黎大洪,谢春元,等.阿魏酸酯酶处理对羊草、玉米秸、稻秸及麦秸瘤胃体外发酵特性的影响[J].动物营养学报,2009,21(6):207-211.
[11] ØRSKOV E R.Manipulation of rumen fermentation for maximum food utilization[J].World Review of Nutrition Dietetics,1975,22:152-182.
[12] MENKE K H,RAAB L,SALEWAKI A,et al.The estimation of the digestibility and metabolizable energy content of ruminant feedstuffs from the gas production when they are incubated with rumen liquor in vitro[J].Journal of Agricultural Science,1979,93:217-222.
[13] NSAHLAI I V,SIAW D E K A,OSUJI P O.The relationships between gas-production and chemical-composition of 23 browses of the genus Sesbania[J].Journal of the Science of Food and Agriculture,1994,65(1):13-20.
[14] BRAVO D,MESCHY F,BOGAERT C,et al.Effects of fungal phytase addition,formaldehyde treatment and dietary concentrate content on ruminal phosphorus availability[J].Animal Feed Science and Technology,2002,99(1/2/3/4):73-95.
[15] BERGMAN E N.Glucose metabolism in ruminants as related to hypoglycemia and ketosis[J].Cornell Veterinarian,1973,215:865-873.
[16] ALLISON M J,BRYANT M P.Metabolic function of branched-chain volatile fatty acids,growth factors for Ruminocoocci[J].Journal of Dairy Science,1961,43:184-198.
[17] ZHANG D F,YANG H J.In vitro ruminal methanogenesis of a hay-rich substrate in response to different combination supplements of nitrocompounds;pyromellitic diimide and 2-bromoethanesulphonate[J].Animal Feed Science and Technology,2011,163:20-32.
[18] YANG H J,YUE Q.Effect of glucose levels and N sources in defined media on fibrolytic activity profiles of Neocallimastix sp.YQ1 grown on Chinese wildrye grass hay or alfalfa hay[J].Asian-Australasian Journal of Animal Sciences,2011,24(3):379-385.
