崇明东滩湿地芦苇和互花米草种群的分布格局及其与生境的相关性
2012-12-14李德志1b1c1b高锦瑾吕媛媛
潘 宇,李德志,1b,1c,①,袁 月,1b,徐 洁,2,高锦瑾,吕媛媛
(1.华东师范大学:a.环境科学系,b.上海市城市化生态过程与生态恢复重点实验室,c.浙江天童森林生态系统国家野外科学观测研究站,上海200062;2.上海海事大学应用数学系,上海201306)
芦苇〔Phragmites australis(Cav.)Trin.ex Steud.〕为本土植物,在中国很多滨岸湿地生态系统中常作为优势种存在,并在生态系统结构和功能的维护过程中发挥了主要作用。互花米草(Spartina alterniflora Loisel.)系多年生草本植物,原产于大西洋西海岸及墨西哥湾,主要生长于平均海平面至平均高潮位之间的广阔滩面以及河口湾[1],由于其具有显著的保滩护岸和促淤造陆的作用而被许多国家引种栽培[2]。1979年12月互花米草被引入中国[3],目前已广泛分布于辽宁、天津、山东、江苏、上海和广西等10个省市区的沿海河口盐沼地带。然而,由于引入后迅速扩散,互花米草在短期内给当地自然环境造成了较为严重的生态影响,与本土植物芦苇产生了不同程度的生态竞争(在某些生境甚至发生比较严重的排挤效应),同时也造成滩涂养殖等产业的直接经济损失。虽然对于互花米草引入后长期生态效应的科学评估和定论还为时尚早,但长期、深入的研究结果无疑可为此提供重要的科学依据。互花米草的扩散及其与当地优势种芦苇之间关系的演进历程具有典型的生态学特征,尤其是在上海崇明东滩更是如此[4]。明确种群的空间分布格局是研究其空间利用方式以及种间关系的基础。盐沼植物群落的演替与种群的空间分布格局具有一定的联系[5-6]。由于种群对环境的适应以及种群内个体间的相互作用,在一定的环境条件下,种群会呈现出特定的空间分布格局,这种格局随时间而变化。因而,研究外来种互花米草种群的扩散机制及其与本地优势植物芦苇种群的竞争关系,并了解其各自的空间分布格局及其相互关系,对于丰富现有的生态学理论以及科学指导滩涂保育实践都具有十分重要的科学价值。
李贺鹏等[7]利用遥感技术研究了2003年上海地区互花米草的分布现状;黄华梅等[8]利用GIS手段分析了1998年至2005年崇明东滩盐沼植被的时空演替动态过程。上海崇明东滩植被种类组成虽然简单,但在某些局部区域芦苇与互花米草等优势种密集混生,使得遥感影像中的混合像元相对较多,加之分辨率限制(商用ASTER地面分辨率为15 m)等原因,导致目前还难以在较小范围内(10 m×10 m)完全通过遥感手段精确反映崇明东滩芦苇和互花米草种群斑块的分布格局。
作者采用GIS技术与野外调查相结合的方法,在1 500 m×100 m尺度上对上海崇明东滩芦苇与互花米草种群的镶嵌分布格局进行研究,并通过选取典型环境参数,探讨这2种植物种群的分布格局与生境异质性的关系,以期为深入了解和分析上海崇明东滩互花米草种群的扩散状况及其与本地种芦苇种群的生态关系以及崇明东滩湿地的生物多样性保护和生态系统经营提供一定的理论依据。
1 研究地概况和研究方法
1.1 研究地概况
实验地设在上海崇明东滩湿地潮间带。崇明东滩湿地位于长江入海口,在上海市崇明岛东端,地理位置为东经 121°50'~122°05'、北纬 31°25'~31°38',属东部亚热带季风性气候,四季分明、温和湿润。年平均日照时数2 138 h;年均气温15.3℃;最热月为7、8月份,月平均气温26.8℃ ~26.9℃;最冷月为1月份和2月份,月平均气温3.0℃ ~3.9℃。年平均无霜期约320 d,雨量丰富,年均降水量1 022 mm,空气相对湿度82%。崇明东滩地处亚太候鸟迁徙路线中段,是东亚最大的候鸟保护区之一,2001年被列入国际重要湿地名录(编号1144),2005年经国务院批准成为国家级自然保护区[9]。
1.2 研究方法
1.2.1 种群分布格局研究方法 于2010年7月,在崇明东滩捕鱼港设置1条垂直于堤坝且自西向海走向的样线作为研究样地,详见图1。该样地东西向长1 500 m、南北向宽100 m,在标准坐标纸上绘制芦苇与互花米草种群在自然生境中的分布格局图,并采用Arcgis 9.3软件将其数字化。计算各类斑块的相关景观水平格局指数[10],以刻画它们的镶嵌式分布格局特征。利用Fragstats 3.3软件计算景观指数值,并对芦苇与互花米草种群斑块进行镶嵌分布格局分析。选取的典型景观指数包括:景观空间形态指标(包括斑块密度PD、最大斑块指数LPI、边缘密度ED和景观形状指数LSI)、景观空间关系指标(包括散布与并列指数IJI、聚集度指数AI、连通度指数CONNECT、蔓延度指数CONTAG和景观分裂指数LDI)以及景观组分指标(包括 Shannon多样性指数 SHDI和Shannon均匀性指数SHEI)。
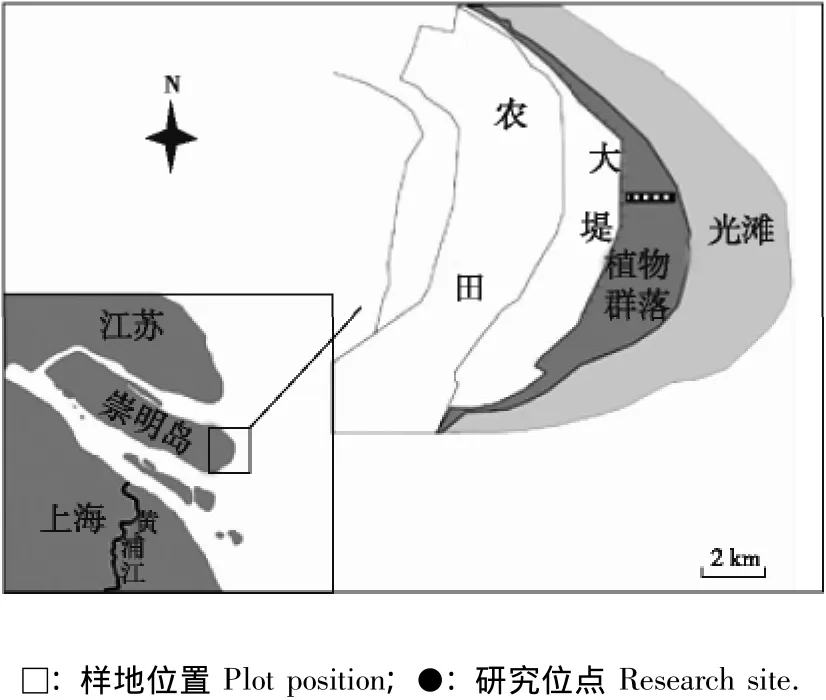
图1 上海崇明东滩湿地研究样地和位点示意图Fig.1 Diagram of research plots and sites in Chongming Dongtan wetland of Shanghai
1.2.2 种群分布特征研究方法 于2010年8月,按照与大堤距离的远近划分高、中高、中、中低和低5个潮位生境,分别在5个潮位生境中各选取1个研究位点,相邻位点间隔300 m(图1),每个研究位点随机划分3个面积10 m×10 m的样方,分别调查样方内芦苇和互花米草种群活体与立枯体植株的数量,并分析其相关性。
1.2.3 环境参数测定 选测4个环境参数,包括相对高程、土壤盐度、土壤中总氮和总磷含量。其中,相对高程用GPS72定位仪(北京合众思壮科技股份有限公司生产)测量。在落潮时期采集各样方的土样,每个样方中每隔100 cm设置1个土壤样点,设5次重复。采用半径5 cm环刀取地表下0~10 cm土壤样品,用自封袋密闭保存并带回实验室。采用烘干法测量土壤含水量。按劳家柽等[11]的方法测定土壤盐度,将干土研磨成细粉,取20 g土样,用100 mL蒸馏水稀释后用DDS-11A型电导率仪(上海精密仪器仪表有限公司生产)测量土壤电导率,并计算土壤盐度。剩余土样过100目筛,使用Skalar Santt流动注射分析仪(荷兰Skalar公司生产)测定总氮和总磷含量。
1.3 数据分析方法
使用Excel 2003软件对原始数据进行初步整理;分别用Arcgis 9.3和Fragstats 3.3软件完成景观分布格局的制图和景观指数的计算;采用SPSS 16.0统计分析软件对数据进行差异显著性分析和Pearson相关性分析。
2 结果和分析
2.1 芦苇和互花米草种群的分布格局分析
2.1.1 斑块分布格局分析 在1 500 m×100 m尺度上,上海崇明东滩芦苇与互花米草种群的分布格局见图2。由图2可见:2个种群呈明显的斑块镶嵌分布格局。芦苇种群主要分布于高潮位和中潮位地带,在低潮位地带仅呈零星散状分布;其分布沿潮位的降低逐渐减少。互花米草种群主要分布于中潮位和低潮位地带,高潮位地带相对较少;其分布沿着潮位的降低逐渐增多。而空白斑块生境则多见于中高潮位。
2.1.2 斑块面积和周长的比较 上海崇明东滩芦苇与互花米草不同类型斑块的特征参数见表1。由表1可以看出:研究区域内共有42个斑块,其中芦苇种群斑块有25个、互花米草种群斑块有13个、空白斑块(对于面积较小的其他植物斑块以及漕沟等无植物分布的斑块均记为空白斑块)有4个。面积最大的斑块属互花米草种群,斑块面积达40 210.60 m2;其次是空白斑块,斑块面积达30 751.28 m2。互花米草种群斑块数量较少但面积较大,13个斑块的平均面积达5 844.82 m2,明显大于芦苇种群斑块的平均面积(1 603.89 m2)。而芦苇种群斑块数量多但面积小,斑块面积2 000 m2以上的仅有9个,最大斑块面积仅为6 861.22 m2;此外,还有7个斑块面积不足10 m2。空白斑块数量最少,仅4个,各斑块面积相差较大。

图2 上海崇明东滩湿地样地内芦苇和互花米草种群斑块分布图Fig.2 Diagram of patch distribution of Phragmites australis(Cav.)Trin.ex Steud.and Spartina alterniflora Loisel.populations in plot of Chongming Dongtan wetland of Shanghai
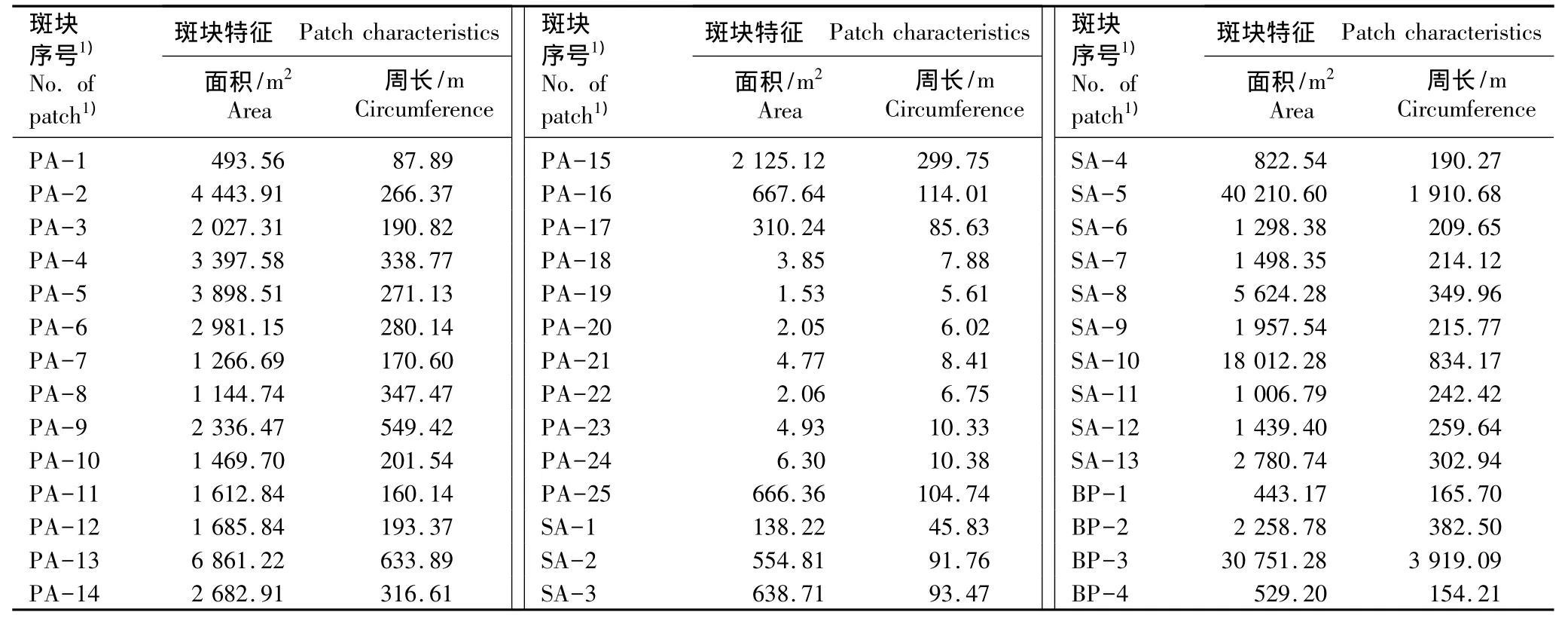
表1 上海崇明东滩湿地内芦苇和互花米草种群斑块的特征Table 1 Patch characteristics of Phragmites australis(Cav.)Trin.ex Steud.and Spartina alterniflora Loisel.populations in Chongming Dongtan wetland of Shanghai
由表1还可见:在各类型斑块中,周长最大的斑块为空白斑块,斑块周长达3 919.09 m;其次为互花米草种群斑块,斑块周长为1 910.68 m。芦苇种群斑块周长为5.61 ~633.89 m,平均周长仅 186.71 m;互花米草种群斑块周长为45.83~1 910.68 m,平均周长为381.59 m,明显大于芦苇种群斑块;而空白斑块周长为154.21~3 919.09 m,景观连通度较大,部分呈集中分布。
上述分析结果表明:3种类型斑块的面积和周长在一定程度上反映了芦苇与互花米草种群在东滩湿地的典型分布特征和生境异质性特点。在中、低潮位,互花米草占据了大面积的生境,较芦苇有明显优势;而在中、高潮位,芦苇种群则占据优势。这一特点说明互花米草种群对东滩湿地低潮位生境有较强的入侵性和适应力。总体而言,崇明东滩芦苇种群生境的破碎化程度大于互花米草种群;互花米草种群斑块间的连通性较高,在中低潮位地带呈大规模集中分布;而空白斑块数量虽少但连通性最高,聚集度最大。
2.1.3 斑块景观参数和格局指数分析 在采用Fragstats 3.3软件计算景观指数时,经检验采用cell size为0.5 m,由此产生的分辨率问题导致矢量图转化为栅格图时产生误差,共识别出37个有效斑块,经多次检验这并不影响研究结果。计算结果(表2)表明:芦苇种群斑块、互花米草种群斑块和空白斑块的分布不均匀。从斑块面积上看,互花米草种群斑块面积最大,占总面积的50.62%,与另2类斑块在面积上差异明显;芦苇种群斑块和空白斑块的面积接近,分别占总面积的26.69%和22.69%。但从斑块数量上看,互花米草种群斑块数量明显少于芦苇种群斑块,表明其扩散分布呈集中化分布特点;而芦苇种群斑块则分割严重,呈现出破碎化特征。从斑块边缘长度上看,芦苇种群斑块边缘长度最大,达5 121.0 m;互花米草种群斑块边缘长度最小,但仍达到4 547.5 m,二者差异不大。
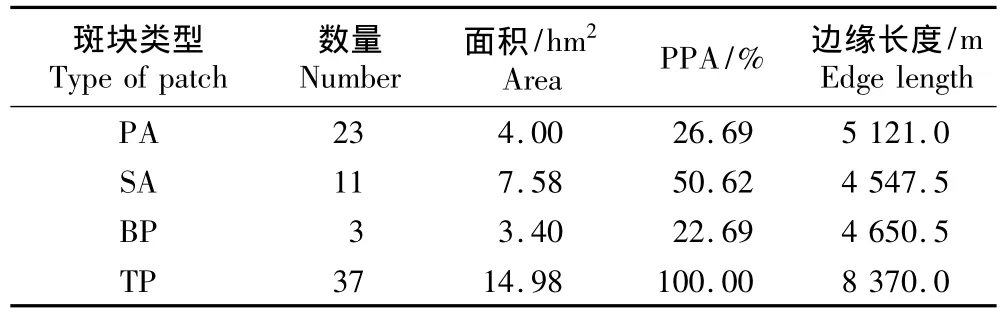
表2 上海崇明东滩湿地内芦苇和互花米草种群斑块的基本景观参数1)Table 2 Basic landscape parameters of patches of Phragmites australis(Cav.)Trin.ex Steud.and Spartina alterniflora Loisel.populations in Chongming Dongtan wetland of Shanghai1)
通常,斑块密度越大,表明一定面积上异质景观要素斑块数量越多、规模越小,同时也意味着景观破碎化程度和空间异质性程度越大;边缘密度则用于揭示景观或类型被边界分割的程度,边界密度越高,说明景观破碎化程度越大。由表3可见:在1 500 m×100 m的尺度内,芦苇与互花米草种群斑块密度较大、斑块规模较小、空间异质性较大。芦苇种群的斑块密度和边缘密度均大于互花米草种群,表明芦苇种群的破碎化程度大于互花米草种群;空白斑块的边缘密度大于互花米草种群,连通性也较高。芦苇种群斑块和互花米草种群斑块以及空白斑块的聚集度指数都较大,连通度指数也较高,均接近100%,表明在1 500 m×100 m尺度上这3类斑块的连通性较强。
对总斑块的分析结果(表3)显示:各景观和斑块类型格局指数数值均较高,较大的边缘密度说明景观的边缘效应显著,斑块边界的分割程度较高、开放性强、易于同周围斑块进行物质和能量交换。景观形状指数仅6.586 8,说明景观形状并不复杂;聚集度指数和连通度指数分别为98.870 1%和99.731 4%,表明该区域景观整体结构较为紧凑且连通性较强;蔓延度指数较高(59.967 1%),说明景观类型较少,连续性良好;Shannon多样性指数(1.033 8)大于1,表明样地已经出现景观破碎化现象;Shannon均匀性指数较大(0.941 0),推测这可能是由于该景观斑块类型和数目相对较少,且各种斑块类型在景观中分布较为均匀所致。
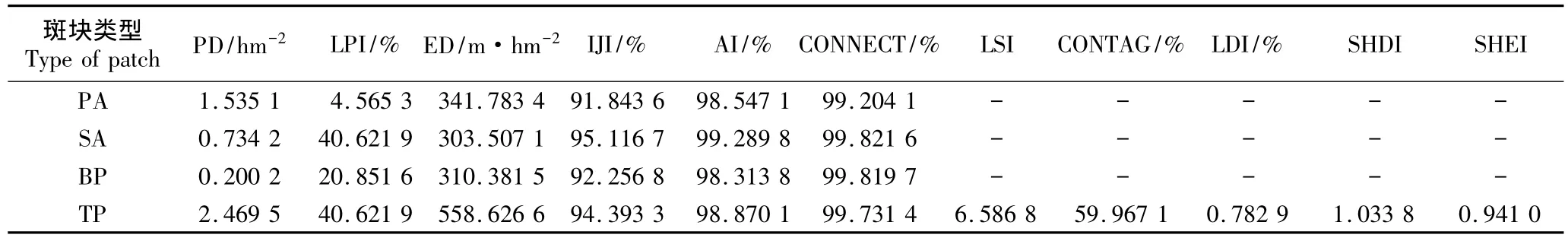
表3 上海崇明东滩湿地内芦苇和互花米草种群斑块的景观及斑块类型格局参数1)Table 3 Pattern parameters of landscape and patch type of Phragmites australis(Cav.)Trin.ex Steud.and Spartina alterniflora Loisel.populations in Chongming Dongtan wetland of Shanghai1)
2.2 芦苇和互花米草种群分布特征分析
2.2.1 沿潮位梯度的数量分布特征分析 植株密度可反映出植物种群适应和利用生境的现状或趋势。上海崇明东滩湿地不同潮位芦苇与互花米草的植株数量见表4。分析结果表明:5个潮位分布的芦苇与互花米草的植株总数无显著差异,表明这2种植物种群对崇明东滩湿地高、中、低各潮位生境的利用程度可能较为相似,且与前述关于不同类型斑块分布格局的分析结果具有一致性。芦苇种群在高潮位和中潮位样方中的分布数量显著高于中低潮位和低潮位样方(P<0.05),倾向于低盐生境,这是由芦苇的基本生态学特性决定的。而中低潮位和低潮位样方中互花米草的植株数量显著高于中高潮位和中潮位样方(P<0.05),反映出互花米草的喜(耐)盐性和喜(耐)水特性。另外,互花米草种群植株总量中活体植株比例较高,而立枯体比例较小,表明互花米草种群在崇明东滩湿地生境中的生长扩散时间较短。加之互花米草立枯体在高潮位样方内的植株数量显著低于低潮位样方,推测互花米草在高潮位样方的扩散时间最短、种群年龄较小。
值得注意的是,高潮位样方中互花米草活体的数量显著高于芦苇活体数量(P<0.05),与中低潮位和低潮位样方中二者的差异相似,其原因与高潮位样方中土壤水分和盐度较高、有利于互花米草种群在局部生境的生长和扩散有关。低潮位样方内互花米草立枯体的株数显著高于芦苇立枯体(P<0.05),反映出在过去时段内互花米草种群在低潮位生境的分布状况及其较芦苇种群的优势地位。而在高潮位、中高潮位以及中潮位样方中,二者立枯体的株数均无显著差异,说明在过去时段内2种植物种群在这一生境范围内对资源的竞争能力差异不大。
表4 上海崇明东滩湿地内不同潮位样方中芦苇和互花米草植株数量的统计结果(±SE)1)Table 4 Statistical result of individual number of Phragmites australis(Cav.)Trin.ex Steud.and Spartina alterniflora Loisel.in quadrats at different tidal levels of Chongming Dongtan wetland of Shanghai(±SE)1)
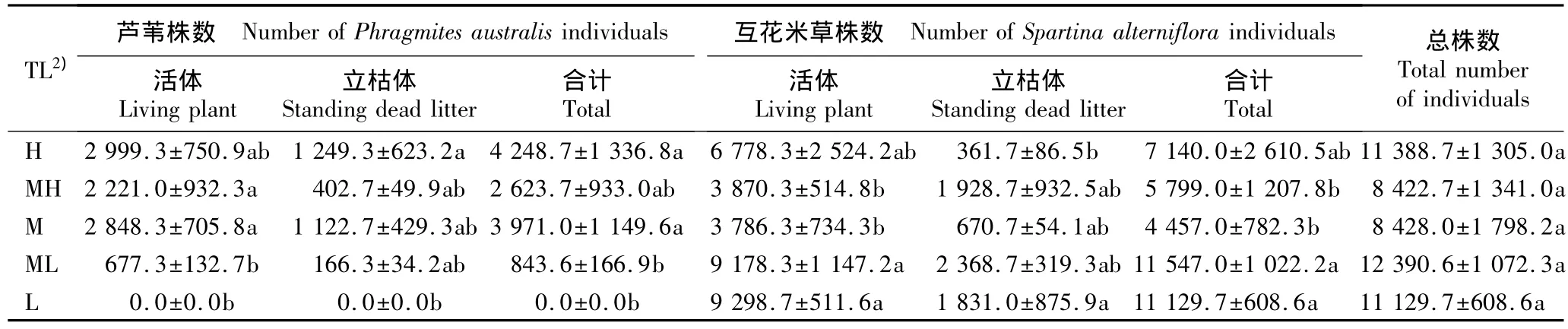
表4 上海崇明东滩湿地内不同潮位样方中芦苇和互花米草植株数量的统计结果(±SE)1)Table 4 Statistical result of individual number of Phragmites australis(Cav.)Trin.ex Steud.and Spartina alterniflora Loisel.in quadrats at different tidal levels of Chongming Dongtan wetland of Shanghai(±SE)1)
1)同列中不同的小写字母表示差异显著(P<0.05)Different small letters in the same column indicate the significant difference(P<0.05).2)TL:潮位Tidal level;H:高潮位High tidal level;MH:中高潮位Middle-high tidal level;M:中潮位Middle tidal level;ML:中低潮位Middle-low tidal level;L:低潮位 Low tidal level.
?
2.2.2 植株数量间的相关性分析 对于地上部每年枯萎的植物种群而言,在生长季内分布于生境中的活体株数是该植物种群当前或近期竞争、扩散结果的直观表现,而立枯体的株数则反映了该种群此前较早阶段的竞争、扩散“迹象”。据观察,芦苇和互花米草的立枯体能够在生境内持续站立多年。因此,可以初步认为目前该生境中的芦苇和互花米草立枯体是经过数年时间积累下来的。相关性分析结果表明(表5):芦苇种群活体和立枯体的株数分别与芦苇的总株数呈极显著正相关(P<0.01),互花米草活体株数与其总株数呈极显著正相关(P<0.01);但互花米草立枯体株数与芦苇立枯体株数呈显著负相关(P<0.05),芦苇活体株数与互花米草总株数也呈显著的负相关(P<0.05),说明在多年的积累过程中,互花米草种群植株总数逐渐增多,对生境面积的利用率越来越高,并以此争夺芦苇种群扩散的潜在空间。
2.3 芦苇和互花米草种群植株数量与部分环境参数的相关性分析
2.3.1 沿潮位梯度各环境参数的变化及其相关性分析 统计结果(表6)表明:崇明东滩湿地中潮位样方的相对高程最高(9.2 m),其次为中高潮位样方,高潮位样方的相对高程明显低于前两者但比中低和低潮位样方高。随潮位的降低,土壤盐度从18.01 ng·L-1降至15.68 ng·L-1,然后逐渐升高,低潮位样方的土壤盐度最高,达到37.17 ng·L-1。随潮位降低,土壤总氮和总磷含量表现出相似的变化趋势,即随潮位的降低,土壤总氮和总磷含量总体上均逐渐下降,但土壤总磷含量的最高值出现在中高潮位样方,而土壤总氮含量的最高值则出现在高潮位样方。
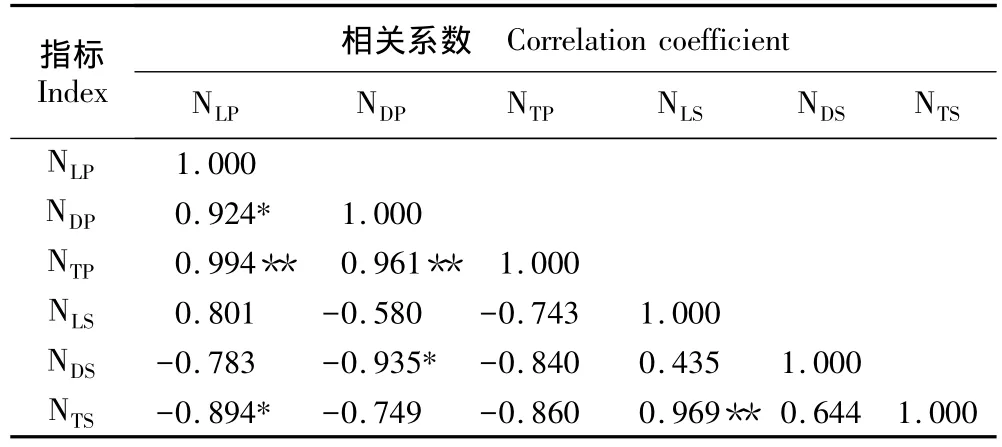
表5 上海崇明东滩湿地内芦苇和互花米草株数的Pearson相关性分析1)Table 5 Pearson correlation analysis of individual number of Phragmites australis(Cav.)Trin.ex Steud.and Spartina alterniflora Loisel.in Chongming Dongtan wetland of Shanghai1)
表6 上海崇明东滩湿地不同潮位样方的部分环境参数测定结果(±SD)1)Table 6 Determination result of some environmental parameters in quadrats at different tidal levels in Chongming Dongtan wetland of Shanghai(±SD)1)
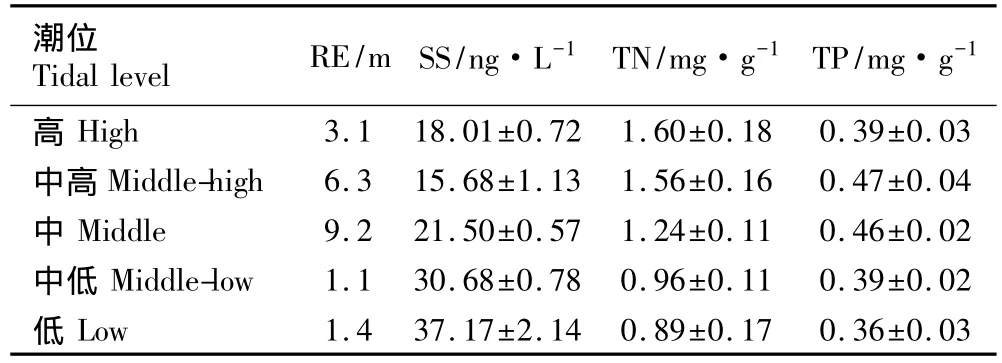
表6 上海崇明东滩湿地不同潮位样方的部分环境参数测定结果(±SD)1)Table 6 Determination result of some environmental parameters in quadrats at different tidal levels in Chongming Dongtan wetland of Shanghai(±SD)1)
1)RE:相对高程Relative elevation;SS:土壤盐度Soil salinity;TN:土壤总氮含量Total nitrogen content in soil;TP:土壤总磷含量Total phosphorus content in soil.
?
一般来说,海岸湿地与海岸距离越近,其相对高程越低。但表7的相关分析结果表明:在上海崇明东滩湿地,相对高程与距堤坝的距离间未表现出明显的线性相关性;但土壤盐度与距堤坝的距离呈显著正相关、与土壤总氮含量呈显著负相关;而土壤总氮含量与距堤坝的距离呈极显著负相关性。另外,土壤总磷含量与相对高程则表现出显著的正相关性。
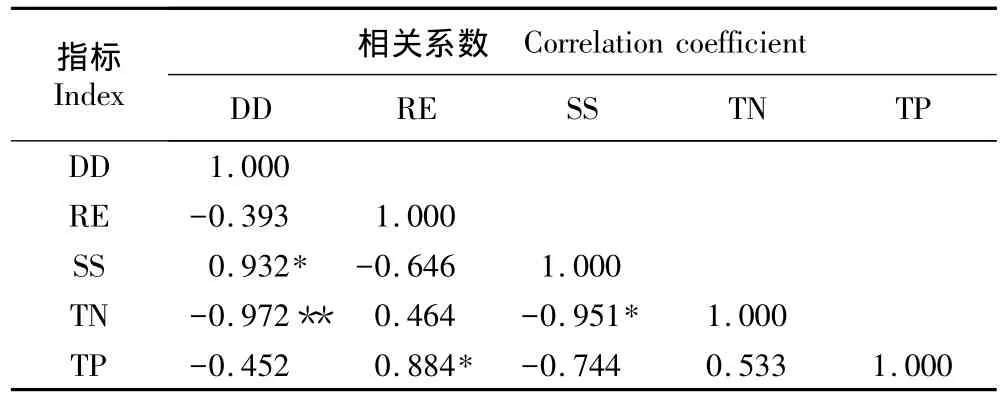
表7 上海崇明东滩湿地内各环境参数的Pearson相关性分析1)Table 7 Pearson correlation analysis of environmental parameters in Chongming Dongtan wetland of Shanghai1)
2.3.2 芦苇和互花米草植株数量与环境参数的相关性分析 上海崇明东滩湿地芦苇和互花米草株数与部分环境因子的相关系数见表8。芦苇种群活体株数与距堤坝的距离及土壤盐度均呈显著负相关(P<0.05),表明随崇明东滩湿地土壤盐度的增大,芦苇生长逐渐受到抑制。互花米草种群活体株数与相对高程和土壤总磷含量均呈显著负相关(P<0.05);互花米草总株数与相对高程呈显著负相关(P<0.05)。一般相对高程较低的生境,淹水时间和频率相应增加,土壤盐度较高,这有利于具有耐淹、耐盐特性的互花米草种群的生长及其与芦苇种群的竞争。
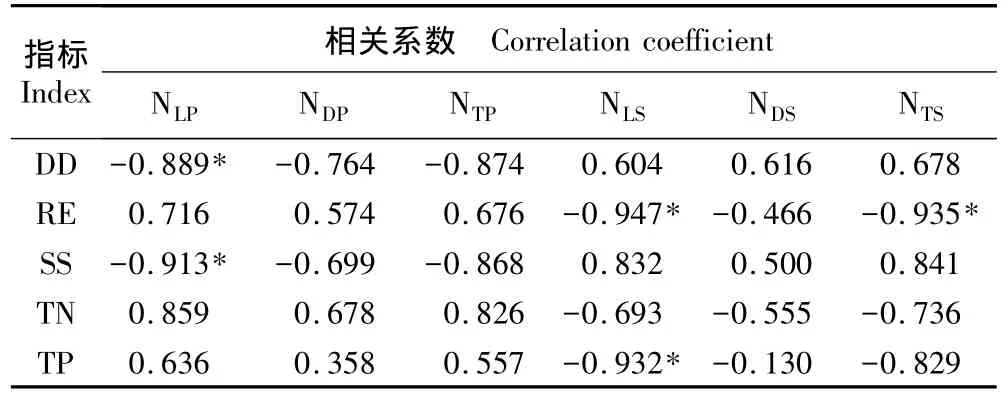
表8 上海崇明东滩湿地内芦苇和互花米草株数与部分环境参数的Pearson相关性分析1)Table 8 Pearson correlation analysis of individual number of Phragmites australis(Cav.)Trin.ex Steud.and Spartina alterniflora Loisel.with some environmental parameters in Chongming Dongtan wetland of Shanghai1)
3 讨论和结论
有研究表明:在淤涨型河口盐沼中,植物群落往往沿高程呈带状分布[12-14]。在属于淤涨型的崇明东滩湿地上,芦苇和互花米草种群的空间分布格局并未呈现典型的带状分布,而是以斑块镶嵌分布为主,但对于涵盖各潮位梯度的整体滩涂而言,中、低潮位生境以互花米草种群为主,中、高潮位生境以芦苇种群为主的格局依然存在。景观及斑块格局分析结果表明:互花米草种群在适宜生境内相对集中分布,具体表现为在低、中潮位分布较多,呈大斑块、集中化特点,斑块间连通性较大,同时有进一步向高潮位适宜生境扩散的趋势;在低、中潮位生境内,互花米草受到的来自芦苇和其他植物的种间竞争压力较小,因而能够快速扩张、占据大面积的生境,并且常以单优群落斑块的形式存在,反映了其在该生境中具有较强的利用资源和占据生境的能力。芦苇种群则基本分布在中、高潮位生境,斑块面积小而数量多,表现出破碎化现象,特别是在中潮位生境中这一现象更为明显。互花米草在中、高潮位的生长和扩散在一定程度上破坏了芦苇种群斑块的连通性,其向芦苇种群分布区的成功入侵是导致这种现象的重要原因。此外,空白斑块多位于高、中潮位生境,在土壤盐度较高的低潮位生境分布较少,这与互花米草在该生境大面积的扩散分布有关,反映了互花米草具有较强的喜(耐)盐、喜(耐)水能力以及较大的竞争优势。空白斑块面积较小,连通性大且开放性强;条件适宜的空白斑块以及目前虽被芦苇或互花米草种群占据但固守不力的斑块将成为互花米草或芦苇种群进一步扩散、竞争的潜在生境。研究结果还表明:样地景观边缘密度总体较高、开放性较强,有利于同周围生境进行物质、能量和信息的交换,这可能为芦苇及互花米草种群的生长和扩散创造一定的有利条件。
在某一生境内何者占据优势,取决于在不同生境内植物的适应性和竞争力的强弱。在低潮位生境内,互花米草较本地种芦苇有明显的竞争优势,随着滩涂的不断淤涨而创造出新的适宜的生境条件,从而为互花米草种群的不断扩散提供更多的生境空间。各潮位生境内芦苇和互花米草活体和立枯体植株的分布特点体现了2种植物分别在空间、时间和资源3个层面上对崇明东滩湿地的适应与选择。从研究结果来看:伴随着滩涂的不断淤涨,目前的总体空间格局基本态势可能不会发生较大的改变,但由于不同潮位的具体位置会发生改变,原来分布的植物种群可能会逐渐发生演替。
由潮汐带来的土壤盐度是河口盐沼植被的典型限制因子之一,土壤中可利用的氮、磷含量也对盐沼植物的生长和扩散起着重要作用。前人的研究结果显示:随着滩涂高程的提升,以土壤盐度、土壤氮和磷含量为代表的盐沼典型环境因子呈现明显的梯度变化[15-16]。而崇明东滩湿地各样方的相对高程、土壤盐度、土壤总氮和总磷含量指标虽基本呈现一定梯度的变化,但与样方所处的潮位并不呈现完全一致的相关性,说明崇明东滩湿地环境因子具有整体梯度明显、局部微生境复杂多变的特征。在样地内,距海岸最远处的高潮位样方的相对高程却有所降低,这与典型的淤涨型滩涂高程规律存在一定差异。这种情况的产生可能与建堤取土有关,也可能与该地段本来的地势有关。在调查中作者发现:高潮位样方内潮沟数量和面积明显增多,推测这里丰富的水流可能会改变局部生境的环境因子,进而影响此处植物种群的分布。高潮位生境中出现的盐分和水分偏高的微生境有利于互花米草生长,进一步导致芦苇活体株数降低、使芦苇的扩散受到抑制。这一现象也为解释在高潮位生境中互花米草的入侵和扩散机制提供了重要线索。局部微生境因子(如土壤盐分和氮、磷含量)的变化对芦苇种群与互花米草种群的扩散和分布格局可能产生复杂的局部性影响,使其分布格局呈现对局部微生境的适应特点。
互花米草种群在上海崇明东滩湿地的成功扩散对本地种甚至整个东滩湿地生态系统带来的威胁至少在目前是显而易见的。陈中义[17]的研究结果表明:互花米草种群在低潮位生境定植后,能迅速地在与本地种海三棱藨草(Scirpus mariqueter Tang et Wang)种群的竞争中取得优势,而且互花米草种群的扩张很有可能将海三棱藨草种群完全排除出该区域。那么,同为本地种的芦苇在与互花米草的竞争过程中将会如何表现呢?前人也曾研究过二者对环境因子的适应和耐受能力[18-19],认为由于土壤盐分等环境因子的限制,芦苇种群的分布主要集中在高程较高、盐度较低的生境。从本研究结果来看:芦苇种群与互花米草种群对空间资源的竞争以中潮位滩涂最为激烈,但并不局限于该区域,在某些高潮位局部区域,互花米草种群也已形成了小规模斑块并且有向部分芦苇种群生境蔓延的趋势。究其成因可能与局部微生境的高程较低导致土壤盐度较高进而不利于芦苇生长却有利于互花米草生长有关,但是否还有其他影响因素,则有待进一步深入研究。此外,从长远来看,局部微生境的环境差异对2种植物动态扩张的速率以及最终分布格局的影响程度尚需进一步研究,对互花米草种群能否在芦苇种群长期固守的高潮位生境内继续扩展也应保持关注。
[1]邓自发,安树青,智颖飙,等.外来种互花米草入侵模式与爆发机制[J].生态学报,2006,26(8):2678-2686.
[2]AYRES D R,STRONG D R.The spartina invasion of San Francisco Bay[J].Aquatic Nuisance Species Digest,2002,4(4):38-40.
[3]徐国万,卓荣宗.我国引种互花米草的初步研究[M]∥南京大学学报编写组.米草研究的进展:22年来的研究成果论文集.南京:南京大学出版社,1985.
[4]王 卿.互花米草在上海崇明东滩的入侵历史、分布现状和扩张趋势的预测[J].长江流域资源与环境,2011,20(6):690-696.
[5]RANWELL D S.Ecology of Salt Marshes and Sand Dunes[M].London:Chapman and Hall,1972:258.
[6]ROZEMA J,VAN DIGGELEN J.A comparative study of growth and photosynthesis of four halophytes in response to salinity[J].Acta Oecologica,1991,12(5):673-681.
[7]李贺鹏,张利权,王东辉.上海地区外来种互花米草的分布现状[J].生物多样性,2006,14(2):114-120.
[8]黄华梅,张利权,袁 琳.崇明东滩自然保护区盐沼植被的时空动态[J].生态学报,2007,27(10):4166-4172.
[9]祝振昌,张利权,肖德荣.上海崇明东滩互花米草种子产量及其萌发对温度的响应[J].生态学报,2011,31(6):1574-1581.
[10]邬建国.景观生态学:格局、过程、尺度与等级[M].2版.北京:高等教育出版社,2007:107-115.
[11]劳家柽,陈小萱,祁明楣,等.土壤农化分析手册[M].北京:农业出版社,1988:365-368.
[12]ADAM P.Saltmarsh Ecology[M].Cambridge:Cambridge University Press,1990.
[13]BERTNESS M D.Interspecific interactions among high marsh perennials in a New England salt marsh[J].Ecology,1991,72(1):125-137.
[14]VAN ANDEL J,BAKKER J P,GROOTJANS A P.Mechanisms of vegetation succession:a review of concepts and perspectives[J].Acta Botanica Neerlandica,1993,42(4):413-433.
[15]EMERY N C,EWANCHUK P J,BERTNESS M D.Competition and salt-marsh plant zonation:stress tolerators may be dominant competitors[J].Ecology,2001,82(9):2471-2485.
[16]SILVESTRI S,DEFINA A,MARANI M.Tidal regime,salinity and salt marsh plant zonation[J].Estuarine,Coastal and Shelf Science,2005,62(1/2):119-130.
[17]陈中义.互花米草入侵国际重要湿地崇明东滩的生态后果[D].上海:复旦大学生命科学学院,2004.
[18]VASQUEZ E A,GLENN E P,GUNTENSPERGEN G R,et al.Salt tolerance and osmotic adjustment of Spartina alterniflora(Poaceae)and the invasive M haplotype of Phragmites australis(Poaceae)along a salinity gradient[J].American Journal of Botany,2006,93(12):1784-1790.
[19]WANG Q,WANG C H,ZHAO B,et al.Effects of growing conditions on the growth of and interactions between salt marsh plants:implications for invasibility of habitats[J].Biological Invasions,2006,8(7):1547-1560.
