仁寿血防疫区人工种植益母草构型分支与生物量配置
2012-11-28何亚平蒋俊明费世民周金星孙启祥彭镇华
何亚平,蒋俊明,费世民**,周金星,孙启祥,彭镇华
(1.四川林业科学研究院,四川成都 610081;2.中国林业学研究院,北京 100093)
与木本植物不同,草本植物具有极短的生殖年龄,其分枝能直接形成生殖枝,或者说草本植物生殖枝形成的过程就是构型分支过程。在个体结构发育与分支过程中,总是存在顶端优先的问题,即在无人为干扰且资源适宜或不足时,个体总是通过减少分支来增加高生长,从而达到开花结实的状态。在资源充足时,往往内部不同方向的生长势都较强,利于基部分支的细胞分裂素合成和运输。据此推理,植物可能在资源水平不同时能形成不同的个体构型,如基部明显型或主干明显型,或谓之顶端优先或基部优先(余茂德等译,1993;Lauri,2007)。植物体这种分支与成体结构形成的过程就是生殖枝内部结构分化,进而形成不同的资源配置与生殖格局,该过程与生殖枝位置与生境紧密相关(刘左军等,2003;操国兴等,2007;Lundgren& Sultan,2005)。位置效应是用来解释这种生殖现象的假说,如露蕊乌头(樊宝丽等,2008)、黄帚橐吾(刘左军等,2003),即中间头状花序最先形成,维管组织优先建立。实际上,这种位置或节点效应根源是结构差异,一则不同位置的结构大小或与先发育维管组织的连通性造成的物质输导能力差异;二则韧皮部厚度、面积驱动的器官承载力大小。
植物构型分支过程也是光合作用不断积累有机物质,并蓄积在体内,生物量配置或生产力配置是构型形成的功能表现,也是结构异质性下非生物生境资源利用的产物。个体大小不同,形成了各器官资源配置格局,并形成了构型适应生境的物质基础。1 a生草本植物能迅速反应构型分支过程,并短时间内完成生活史过程,很直观地反应了构型发育。
益母草(Leonurus heterophyllus Sweet.)是唇形科植物,生物碱类成分由于含量较高,活性较强,一直被认为是益母草属植物的有效成分。益母草全草含生物碱0.11% ~2.09%,其中益母草碱(Leonurine)0.02% ~0.12%,水苏碱(Stachdrine)0.59% ~1.72%(厚毅清等,2010)。在血吸虫病中间宿主钉螺防控上,益母草生物碱的化感作用能抑制钉螺发育(唐万鹏等,2006;胡兴宜等,2007;唐万鹏等,2007;胡兴宜等,2005;宋从文等,2005;唐万鹏,王万贤,2006),并具有良好的经济效益。益母草在长江下游疫区有栽植试验,在长江上游疫区仁寿示范区也有小规模实验性栽植,基于钉螺孽生生境改造的生态位替代理论,分别选择山丘区富集的田埂与旱地间作系统来栽植益母草,增加抑螺防病林的经济性。益母草生长发育过程研究相对较多,如施肥、经营模式、不同种源等益母草的生长观测(盛束军等,1998;徐建中等,2007;张燕等,2007;张丽梅等,2009;姜华年,赵鹏,2011),对构型分支过程中结实特性与生物量配置,及其生境适应性问题缺乏认识。
本文基于植物生长与发育过程中构型分支、生殖枝特性与生物量配置的研究现状与实践需要,对益母草经济性状形成过程中的关键问题进行探讨,并从稳定群落建立的要求来重点认识种子形成的结构限制,为优化栽培与产量提高提供理论依据与科技支撑。四川仁寿县有益母草野生分布,栽培中还未成功的报道,结合疫区益母草试验栽培,初步报道了益母草栽培的基本生长指标。
1 材料与方法
四川仁寿县属于低山丘陵地貌类型,该区血吸虫病流行严重,是四川报道最早的地方。该地为中亚热带季风性湿润气候,冷热四季分明,干湿季明显,多低温寒潮;年平均气温为17.4℃,≥10℃年积温为5 532.8℃,无霜期为310 d,年平均降水量为1 009.3 mm,全年日照为1 196.6 h,全年太阳辐射为894.184 kJ·cm-2。浅丘、平坝区土壤类型以老冲积黄壤和紫色土为主,土层深厚,土壤微酸;中丘和深丘土壤类型以棕色土为主,兼有黄沙土、冷沙黄泥土等类型分布;低山区土壤以紫色石骨土、红砂土、黄泥土为主,土层较薄,水土流失严重。
1.1 示范区概况
示范区建立在仁寿县洪峰乡新建村3队,以水系为中心,对人居、水岸、路渠、坡耕地、田埂、下湿田进行血防林培育,从整体上优化水资源配置,并系统整体防控与局部防控相结合,形成水系廊道、道路廊道格局,将整体环境改造与局部抑螺相结合。示范区自然植被为马尾松、柏木、苦楝、青冈、桉树、慈竹等,栽植树种以桉树、樟树、慈竹、香椿、苦楝、五角枫、湿地松、枫杨、核桃等,还有小叶女贞林带、香根草草带。对药用价值经济草本植物试验栽植有益母草,在示范区下部田埂上和上部旱地农林间作桉树-钓樟林下进行带状栽植。
1.2 试验设置
选择仁寿山丘区血防林用地中两种常见的土地类型,一种是新造林林下,一种是田埂。前者是旱地综合高效利用,后者是坡坎用地的高效利用。从生境资源上看,下部田埂土壤肥厚,水分充足,而山上旱地土壤瘠薄,水分相对不足。这两种生境的资源富集程度不同,代表了益母草栽培的两种土地利用类型。
1.3 调查方法
益母草一级分支是从基部发出,二级分支从一级分支上发出,但二级枝、三级枝都是果枝,直接开花结实。2011年7月~8月,对2010年在下部田埂与间作生境中栽植的益母草进行高度测量,统计一级分支数、二级分支数、三级分支数,两类生境各选择37株和38株。同时选择壮枝收集果序,并保存完好,避免种子散布,在室内对果序上不同分节部位的果实数量、节点临近枝径、相应节间长度进行测量,测量样本分别为6枝和6枝。用筛子将果序分解为上部、中部和下部3个部分,并收集种子若干,以200粒为单位,测定千粒重,重复6次。在间作型生境中选择不同大小个体36株,挖取全株,在室内清洗根际泥土,并测定根系、茎杆、叶片、果实生物量,并取各器官样品用烘箱测定水分含量。
1.4 数据分析方法
将数据录入计算机,并对成对数量进行散点图拟合曲线,并进行差异性、相关性检验。
2 结果与分析
2.1 个体构型分支
与间作生境相比,埂边生境资源相对充足,个体高度明显较高(f=73.252,p=0.000),一级分支数量多(f=1.287,p=0.260),二级枝、三级枝数量大(f=43.567,p=0.000;f=6.915,p=0.010),分支率较高(f=12.512,p=0.001)。新建村3队两种生境条件下益母草构型分支特性如表1所示。
两种生境下益母草高度与二级生殖枝分支数量散点图拟合曲线如图1所示。埂边生境益母草高度与二级生殖枝数量之间拟合曲线符合方程:y=21.199x2-46.987x+49.429(R2=0.3617);表明高度较低时生殖分支数量较低,而在高度增加时生殖枝分支数量增加缓慢,但分支数量起点较高,曲线靠近高度高值区。相比下,间作生境益母草高度与二级生殖枝数量之间拟合曲线符合方程:y=54.588x2-78.365x+31.811(R2=0.6101),也表明间作时高度起点较低,且高度增加时二级枝生殖枝数量增加迅速。
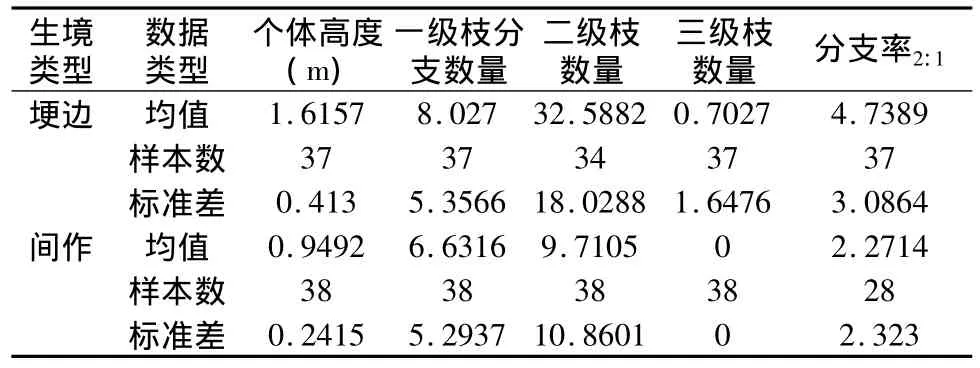
表1 新建村3队两种生境条件下益母草构型分支特性

图1 两种生境下益母草二级生殖枝分支数量随着高度递变拟合曲线
2.2 二级枝的生殖特性
两种生境下益母草二级枝生殖特性如表2所示。总体上,埂边生境的平均枝径、间距长度、果实数量与间作生境无明显差异(f=0.78,p=0.386;f=0.38,p=543;f=365,p=552)。暗示,草本益母草资源配置存在差异时,其通过生殖构型分支引起了树体结构的差异,形成对生境的适应性策略,而在二级生殖枝诸多特性,如枝径、节点间距、果实数量上并不具有可塑性。
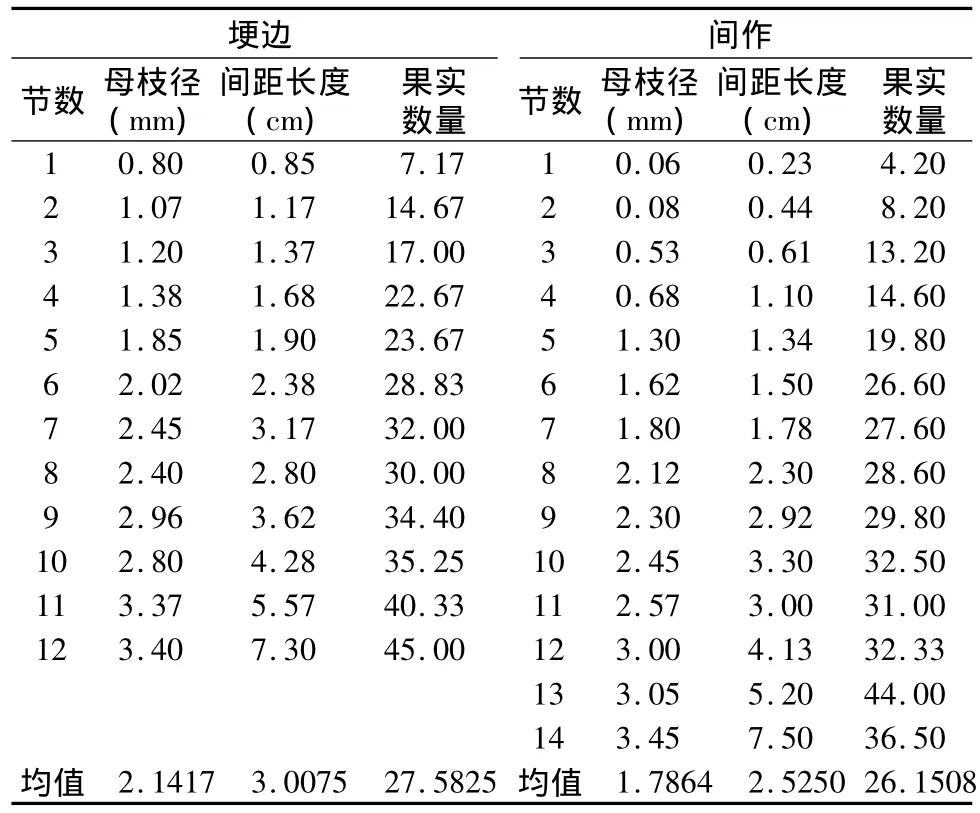
表2 两种生境下益母草二级枝生殖特性
两种生境下益母草节数、果序临近枝径、间隔距离与果实数量相互拟合曲线如图2所示。二级生殖枝节数与果实数量之间拟合曲线符合方程:y=-0.0897x2+4.1575x+5.4161(R2=0.9683,埂边生境)、y=14.049Ln(x)-0.3559(R2=0.9186,间作生境),果实数量在二级生殖枝节数增加中持续增加,且间作生境增加相对较快。二级生殖枝节数与枝径之间拟合曲线符合方程:y=0.2464x+0.5398(R2=0.9788,埂边生境),y=0.2644x-0.1967(R2=0.9803,间作生境),表明节数增加导致果序临近枝径持续增加;从曲线位置可知,埂边生境枝径不明显高于间作生境。节数与节间距离之间拟合曲线符合方程:y=0.7832e0.1782x(R2=0.982,埂边生境)、y=0.3186e0.2274x(R2=0.9398,间作生境),表明节数增加导致果序间距增加,且埂边生境不明显偏高而曲线位于上方。果序临近枝径与果实数量、果序间距与果实数量之间分别呈现直线方程:y=9.7737x+7.4635(R2=0.926,埂边生境)、y=11.96x+1.9691(R2=0.9521,间作生境),与对数方程:y=16.872Ln(x)+12.164(R2=0.9873,埂边生境)、y=11.106Ln(x)+18.685(R2=0.9224,间作生境),表明,两种生境下果实数量与枝径直线增长,而随着间距长度对数增长。从结构上分析,生殖枝下部枝径与上部枝径递变与间距长度递变未能呈现很好的直线增长关系,二者也是曲线关系(y=1.0378x1.316,R2=0.9452;y=0.38e0.856x,R2=0.9499),这是两种生境下果实数量与枝径、与间距长度之间呈现两种曲线的原因。
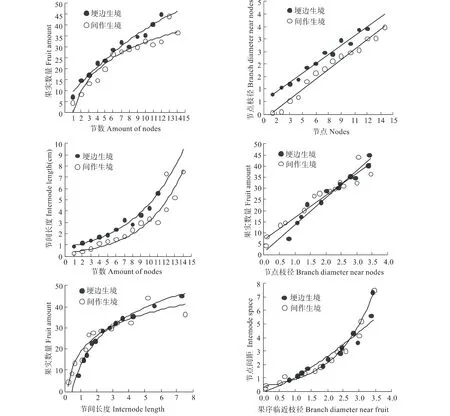
图2 两种生境下益母草节数、果序临近枝径、间隔距离与果实数量相互拟合曲线
2.3 间作益母草生物量及其器官配置
间作生境个体生物量及其器官配置受到个体大小的影响。间作生境下不同生物量级下个体益母草生物量的器官配置如表3所示。分别以5 g为单位,划分生物量等级,则从1级~10级,生物量及其器官配置均呈现增长状态,其个体生物量浮动在1.1 g~108.47 g之间,叶片生物量为0.03 g~12.31 g之间,花果生物量为0.24 g~18.95 g,枝生物量为0.42~52.93 g,根系生物量在0.41 g~24.29 g。益母草生物量的器官相对配置:叶片配置为6.97%、花果配置为16.15%、枝条配置为42.84%、根系配置为34.05%。
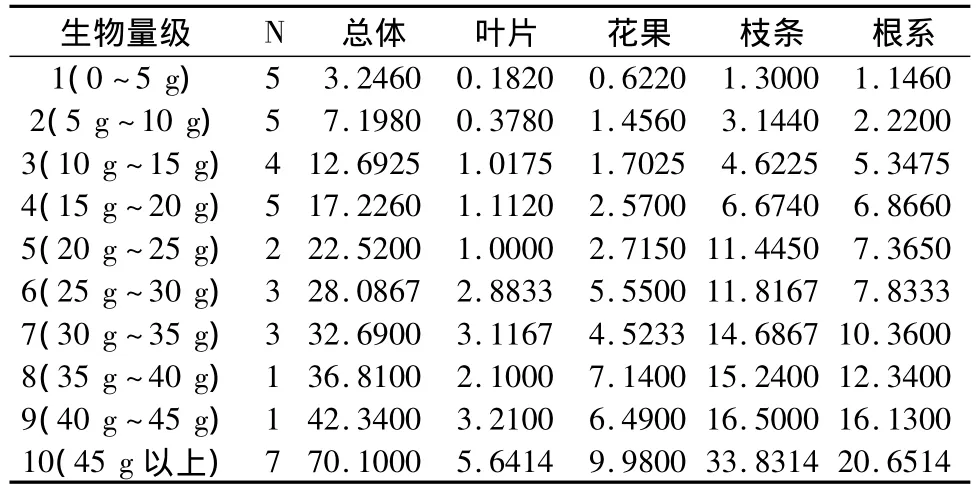
表3 间作生境下不同生物量级下个体益母草生物量的器官配置
对间作生境益母草个体生物量与器官配置之间散点图进行线性拟合(如图3所示),叶片、花果、枝条、根系符合直线方程:y=0.088x-0.2645(R2=0.7077)、y=0.1448x+0.1776(R2=0.87)、y=0.2769x+1.1617(R2=0.874)、y=0.4905x-1.0759(R2=0.9651)。
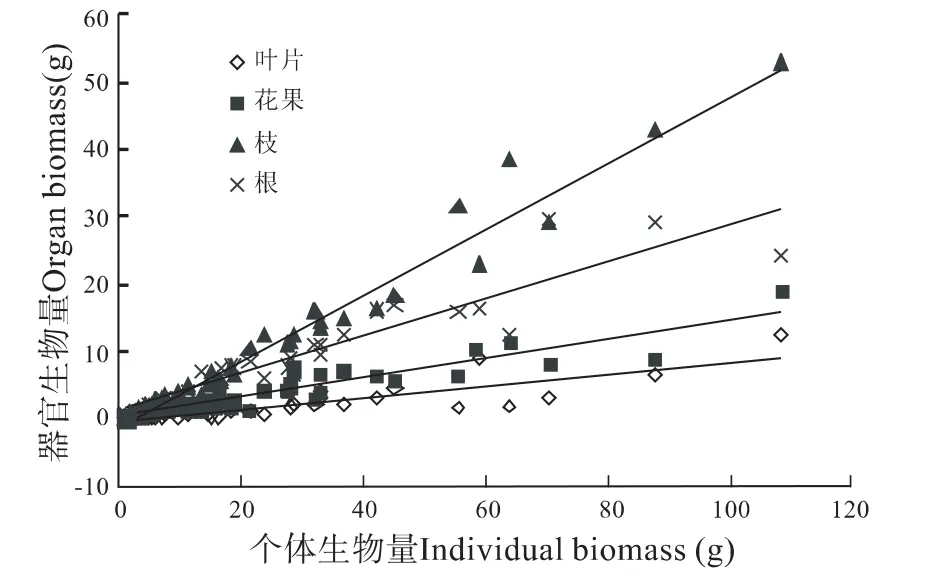
图3 益母草个体生物量与叶片、花果、枝、根生物量之间拟合曲线的散点图
2.4 生殖枝不同部位种子千粒重
益母草二级生殖枝上部种子千粒重为0.7252 g±0.08556 g,中部为种子千粒重为 0.8729 g±0.06789 g,下部种子千粒重为0.8794 g±0.05299 g。方差分析表明,生殖枝不同部位种子千粒重之间存在极显著差异(df=1,f=9.286,p=0.002),且上部与中部、下部保持显著差异,但中、下部之间没有差异。益母草生殖枝不同部位的千粒重的柱状误差图如图4所示。

图4 益母草生殖枝不同部位的千粒重
3 讨论
益母草一级枝从基部发出,其分枝才具有生殖能力,这种分支依照木本植物构型枝级划分方法(何亚平等,2010),称之为二级枝。益母草一级枝发生一般基于水分充足或主干枝受到干扰,调查中存在两种构型,一种是主干优先型,一种是基部优先型(余茂德等译,1993;Lauri,2007)。前者是干旱瘠薄生境中生殖构型,后者是水肥充足或主干受到结构干扰时的生殖构型。显然,无论是埂边生境,还是间作生境,二级生殖枝果序临近枝径、间距长度、果实数量都表现为随着节点位置增加而增大。在水肥充足时益母草头年建成幼苗基部分支明显而造成一级枝明显增加;在缺乏顶芽受损时,基部围绕主干分支与主干枝一起生长。在水肥相对不足时,基部分支能力相对减弱,尤其是密度效应存在时无分支发生,偶有伴随主干与基部分支并存的构型。
理论上分析,在一级分支数量较多时,可能会降低个体高度,实际却是高度与一级枝分支数量呈现不显著正相关(间作生境:r=0.219,p=0.186;埂边生境:r=0.280,p=0.093)。暗示,在特定生境中,高度生长充足时一级分支增加概率相对增加。钉螺孽生草本群落中,生态位替代要求在自然或人为经营条件下能占据优势生态位,形成优势群落,替代钉螺孽生的草本群落。益母草缺乏克隆生殖特性,无法长久占据生态位,只能在人工播种、除草、施肥条件下形成优势群落。其高生长在优势群落形成中意义明显,而土壤水分张力较大时高生长明显;而在水分张力相对较小时,肥力补充也能增加高生长潜力,从而形成群落中较大的高度。埂边生境有渠水供应,且土层深厚,土壤水分张力相对较大,成熟个体高度较大(1.6157±0.413,n=37),而间作生境位于丘顶坡面上,岩层较浅,土层较薄,土壤水分张力相对较小,成熟个体较小(0.9492±0.2415,n=38)。
在肥沃的埂边生境中,生殖枝本身果序着生节点临近的枝径、节点间距与各节点果实数量并无明显差异,生境驱动的结实特性差异主要体现在构型可塑性响应上,即通过一级枝、二级枝,甚至是三级枝数量来适应于土壤资源条件。在生殖枝水平上,单一节点上枝径、节点间距与结实数量都未能反映这种差异;生境差异对构件器官的影响除了构件分支大小有关的生殖枝长度外,可能不是表现在果实数量上,而是在果实和种子大小上还存在不同。统计分析也表明,生殖枝上部、中部和下部种子千粒重存在极显著差异,表明其生殖器官水平上对生境的适应可能主要基于质量而不是数量。
益母草自身高度分化直接影响了居群内生物量的分异,进而形成了大小依赖的生物量器官配置模式。在无密度制约时,这种个体大小差异反映了居群内资源异质性,也是生境差异所致。益母草各器官绝对生物量配置都与个体总生物量呈现明显的直线正相关(如图3所示),表明随着个体大小增加,茎杆、根系、花果、叶片生物量增加速度依次降低,暗示个体增加时营养结构性维持资源配置增加最大。考虑相对生物量时,随着个体大小增加,叶片、花果、根系相对配置未表现明显的大小依赖性,即个体大小增加并未引起生物量配置的增加,二者呈现均不显著直线关系(r=0.256,p=0.132;r=-0.233,p=0.172;r=-0.306,p=0.069)。仅有茎杆生物量随着个体大小呈现显著正相关(r=0.37,p=0.027),表明个体增加引起了茎杆生物量积累,配置比例增加,拟合为线性方程y=0.001x+0.4009(R2=0.1366)。在收获器官不同时,这种维持性结构资源配置随着个体大小的无条件增加降低了定向培育目标产量,尽管益母草为全草入药,但根系部分难以清洗,可能并不适宜利用。从培育上考虑,增加肥料与人工管理,利于提高生物量,并提高其品质,这一点从肥沃埂边生境相对较大构型与分支率看出;从生态适应上考虑,益母草在肥沃生境中个体较大,构型从分支数量、分支率上作出可塑性调节,且具有较大的茎杆配置,以增加高大个体的物理支撑作用。
在两性花植物中,适合度成分常随种群、同一种群内的不同个体或同一花序的不同部位而发生变化(操国兴等,2007);在同一生殖枝上,不同位置结实量和种子大小也不同。益母草器官结实受到生殖枝自身位置特性的影响,在构件水平上表现明显的结构决定性。益母草生殖枝各节点果实数量与节点临近生殖枝枝径、间距长度有关,较粗较长的枝条能在节点上形成较大数量的果实。从顶部节点到基部节点,果实数量逐渐增加,并且枝径、节点距离不同而形成不同的增长模式,曲线或直线(如图2所示):生殖枝果实数量与节点位置、间距之间呈现抛物线、对数曲线,与枝径呈现直线方程。这种生殖枝节点间长度从基部向顶部递减是结构与生长速率共同造成的,是资源权衡配置的结果。位置效应现象实质上是结构限制问题。这种结构限制不但影响了果序上从基部到顶部的果实数量,还影响了种子大小,即除了果实数量上响应于结构递变,还在果实大小上呈现递减趋势。测定表明,上部种子大小小于中部和下部,就表明这种大小适应性(如图4所示)。Diggle(1995)认为果实与种子发育模式的解释与解剖构型有关。由于开花顺序不同,黄帚橐吾顶部种子生产大于基部(刘左军等,2003)。兰玉簪结籽基部种子重量大而顶部种子重量低(操国兴等,2007)。Polygonum hydropiper杂交后代的可塑性依赖于子代在母体上的构型位置(Lundgren&Sultan,2005)。生殖枝基部与顶部之间种子千粒重差异实质上是基于开花与维管束形成早晚,而黄帚橐吾与蓝玉簪二者相反。益母草与蓝玉簪类似,暗示了结构限制实质上是发育上资源配置早晚的问题,与枝条解剖学(Diggle,1997)有关。
益母草传统上以开花的地上部分入药,其有效成分为益母草碱和水苏碱,具有明显的抑螺效果(唐万鹏等,2006;胡兴宜等,2007;唐万鹏等,2007;胡兴宜等,2005;宋从文等,2005;唐万鹏,王万贤,2006)。人工措施(如施肥)能调节益母草生境资源水平,进而塑造生长高度与结实状况,进而提高了生产量(盛束军等,1998;徐建中等,2007;张燕等,2007)与水苏碱含量(徐建中等,2007;张燕等,2007)。经营模式不同,益母草营养价值不同,果园套种与露天栽培氨基酸总量和Vc含量不同(张丽梅等,2009)。这种经营模式可能也影响到生境资源水平。仁寿县兴林抑螺示范区中,两种生境下益母草资源充足样地高度大,而分支数量多,生殖枝多,具有较大的生产量,其经济生产与抑螺成分总量都明显优于间作生境。益母草种子千粒重0.66 g~1.31 g,可化为3级,实验室内发芽率在25% ~91%之间(胡旋等,2011),仁寿人工种植益母草千粒重为0.72 g~0.87 g。试验发现,适宜的种子萌发温度为25℃ ~30℃(张丽梅等,2009;厚毅清等,2010)或30℃ ~35℃,光照无影响(芦站根等,2010),属于中温萌发型。仁寿县益母草种子成熟在6月~7月份,地表具备了种子萌发的温度光照水分条件,但调查时未发现种子萌发而成的幼苗,表明草本盘结度、地表干扰程度、郁闭度等可能影响入土和幼苗建成,自然更新困难。作为钉螺孽生地生态位替代物种,其本身需要良好的人工措施来促进更新并维持其群落优势状态。这一点与克隆植物不同,克隆植物能无性繁殖,自动建立稳定优势群落,形成自调控系统,如仁寿示范区香根草的栽植。
[1]Diggle P K.Architectural effects and the interpretation of patterns of fruit and seed development[J].Annual reviews of ecology and systematics,1995,26:531 ~552.
[2]Diggle P K.Ontogenetic contingency and floral morphology:the effect of architecture and resource limitation[J].Internation Journal of Plant Sciences,1997,158(suppl.):99 ~ 107.
[3]Kenicer G J,Kajita T,Pennington R T,Murata J.Systematics and biogeography of Lathyrus(Leguminosae)based on internal transcribed spacer and cpDNA sequence data[J].American Journal of Botany,2005,92(7):1199 ~1209.
[4]Lundgren M R,Sultan S E.Seedling expression of cross-generational plasticity depends on reproductive architecture[J].American Journal of Botany,2005,92(2):377 ~381.
[5]操国兴,严娟,等.玉簪花序水平上雌性适合度成分变化及种子数量与大小权衡关系研究[J].西南大学学报(自然科学版),2007,29(6):147 ~150.
[6]樊宝丽,赵志刚,等.露蕊乌头(Aconitum gymnandrum Maxim)花序内位置依赖的性分配[J].生态学报,2008,28(6):2909~2915.
[7]厚毅清,王乃亮,等.益母草属植物种子发芽、细胞培养及其化学成分研究进展[J].中国农学通报,2010,26(18):75 ~78.
[8]胡兴宜,唐万鹏,刘立德,等.益母草抑制钉螺生长的初步研究[J].云南农业大学学报,2005,20(6):875 ~877.
[9]胡兴宜,唐万鹏,等.益母草不同组分的抑螺效果及钉螺酯酶同工酶的影响[J].生态学杂志,2007,26(5):728 ~731.
[10]胡旋,李卫东,等.益母草种子质量分级标准研究[J].种子,2011,30(4):83 ~85.
[11]姜华年,赵鹏.不同种源地益母草生物学特性观测评价[J].江苏农业科学,2011,39(2):356 ~359.
[12]刘左军,杜国祯同,等.影响黄帚橐吾种子生产的因素Ⅰ.生境和花序结构[J].植物生态学学报,2003,27(5):677 ~683.
[13]芦站根,周文杰,等.光照温度和NaCl对益母草种子萌发的影响[J].北方园艺,2010,(20):181~183.
[14]盛束军,郑俊波,俞旭平,等.不同施肥水平对益母草生长的影响[J].植物资源与环境,1998,7(1):31 ~34.
[15]宋丛文,刘立德,等.益母草抑制钉螺的研究[J].湖北林业科技,2005,(5,总135期):7~9.
[16]唐万鹏,王万贤.益母草对钉螺化感作用的试验研究[J].湿地科学与管理,2006,2(4):47 ~53.
[17]唐万鹏,张旭东,等.益母草水苏碱对钉螺生理生化的影响[J].湿地科学与管理,2006,2(4):54 ~56.
[18]唐万鹏,张旭东,等.益母草水苏碱浸杀法对湖北钉螺糖原和蛋白质含量的影响[J].安徽农业大学学报,2007,34(1):82~84.
[19]徐建中,王志安,等.氮肥对益母草产量和药材品质的影响[J].中国中药杂志,2007,32(15):1587 ~1588.
[20]余茂德,何旭东,等.木本植物中顶端优势,顶端支配,梢端优先,基部优先与生长、树形形成的关系[J].蚕学通讯,1993,(3):49~52.
[21]张丽梅,陈熹,陈钟佃.果园套种对益母草生物学特性及品质的影响[J].现代中药研究与实践,2009,22(6):17~19.
[22]张燕,李国雷,等.施肥对益母草生长、土壤酶活性及水苏碱含量的影响[J].南京农业大学学报,2007,30(2):78 ~82.
