夏天无注射液对脑缺血再灌注大鼠海马Ang-2和VEGF mRNA表达的影响
2012-11-01旷明丽余丽梅
旷明丽,余丽梅,张 骏*,吴 芹
(1.遵义医学院附属医院神经内科,贵州 遵义 563003;2.遵义医学院附属医院贵州省细胞工程重点实验室,贵州 遵义 563003;3.遵义医学院贵州省基础药理重点实验室,贵州 遵义 563003)
缺血性脑卒中是临床常见的急性脑血管疾病,其发病率、病死率及致残率很高,故对其防治十分重要。在脑梗死中心坏死区周围存在缺血半暗带,其大量的脑细胞处于休眠或半休眠状态,如在一定的治疗时间窗 (1~3 h)内通过有效的治疗,使原来闭塞的脑动脉血管重新通畅或建立新的侧枝循环,则可使它们的神经功能得到逐渐恢复。
夏天无注射液是从中药中提取的中药注射剂,主要含有普鲁托品等有效成分,临床已用于治疗脑血管病后遗症、坐骨神经痛、风湿性关节炎等疾病。研究表明夏天无中提取的夏天无总碱可抑制大鼠血栓形成,减轻脑栓塞引起的脑水肿[1];对大鼠脑缺血再灌注损伤具有保护作用,可减轻氧化损伤[2]。夏天无注射液对脑缺血损伤也具有保护作用,并可抑制神经元凋亡等[3],还可增加血管性痴呆大鼠海马血管生成素-1的表达[4]。本研究制备大鼠脑缺血再灌注模型,观察夏天无注射液的保护作用,同时分析对大鼠海马血管生成相关的血管生成素-2(angiopoietin-2,Ang-2)和血管内皮细胞生长因子 (Vascular endothelial growth factor,VEGF)表达的影响,以探讨夏天无注射液的可能的作用机制。
1 材料
1.1 药品与试剂 夏天无注射液 (批号:06102001,2 mL含阿片碱0.4 mg)系江西天施康中药股份有限公司产品;尼莫地平注射液 (批号:0203011,2 mg/10 mL)为山东新华制药股份有限公司产品;TTC购自上海山蒲化工有限公司;组织细胞RNA抽提试剂盒购自上海华舜生物工程公司;RT试剂盒、Trizol试剂购自TaKaRa生物工程公司(试剂型号:D312);Ang-2 mRNA引物由美国Sigma公司合成;SYBR GREEN PCR Master Mix为美国ABI公司产品;0.1%DEPC(焦碳酸二乙酯)灭菌水、乙醇、异丙醇、三氯甲烷等试剂为国产分析纯。
1.2 主要仪器 SHZ-88A往复式水浴恒温器 (江苏太仓市实验设备厂);电子天平 (德国赛多利斯公司);TU-1810紫外分光光度计 (北京普析通用分析仪器公司);XW-80A旋涡混合器 (宁波新芝科器研究所);Eppendorf Centrifuge 5417R离心机及逆转录仪 (德国 Eppendorf);BeckMAN CoulTER离心机 (美国贝克曼公司);iCycler荧光定量PCR仪 (美国BIO-RAD);-80℃低温冰箱 (日本三洋公司)。
1.3 动物 雄性SD大鼠,体质量250~300 g,由第三军医大坪医院动物中心提供,均为清洁级动物[生产许可证号:SCXK(渝)20020004],手术前大鼠分组饲养,自由饮食。
2 方法
2.1 MCAO模型的制备和分组 20% 水合氯醛腹腔注射麻醉SD大鼠 (0.18 mL/kg),分离右侧颈总动脉、颈外动脉及颈内动脉等,参照文献 [5],线栓法制备大脑中动脉阻塞 (middle cerebral artery occlusion,MCAO)模型,阻断大脑中动脉血液供应,结扎固定线栓,缝合皮肤切口,栓线外留10 mm。待缺血2 h后再轻拉线头,使栓线头端退至颈总动脉切口处,实现再灌注22 h,术中室温保持在25℃。假手术组栓线插入深度为10 mm。
实验动物分为7组,分别为:正常组、假手术组、模型组、夏天无注射液高 (0.05 mL/kg)、中 (0.025 mL/kg)、低 (0.01 mL/kg)剂量组和尼莫地平 (0.01 mL/kg)对照组,给药组缺血2 h再灌注前30 min肌肉注射相应药物各1次,正常组、假手术组及模型组肌肉注射等体积生理盐水。
2.2 神经功能评价 如表1所示,参考Menzies SA[6]的标准给予评分。缺血2 h后评分在3分及以上者视为模型成功。

表1 脑缺血大鼠神经功能评分
2.3 脑组织梗死面积测定 模型成功者随机每组取大鼠8只,术后24 h断头取脑,去除嗅球、小脑及低位脑干,以冷生理盐水去表面血迹,置-80℃低温冰箱中7~8 min,冻成半硬状,冠状切片分成脑组织5片,然后迅速将脑片置5 mL含有1%氯化三苯四氮唑 (triphenyltetrazolium chloride,TTC)及0.1 mL含有1 mol/L K2HPO4溶液中,避光37℃温孵30 min,其间每隔7~8 min翻动一次,染色后,正常脑组织呈玫瑰红色,梗死组织呈白色;用比重法测定出梗死比重,即梗死脑组织占全脑质量的百分比 (%)=梗死脑组织质量/全脑质量×100%(均为湿质量)。
2.4 RT real-time PCR 按试剂盒说明进行操作,用Trizol试剂和组织细胞RNA抽提试剂盒提取大鼠海马组织RNA,紫外分光光度计测定D(260)和D(280),计算D(260)/D(280)比值和RNA浓度,在1.8~2.0范围内者,按TaKaRa公司的逆转录试剂盒操作说明书,在 Mastercycler Gradient PCR仪逆转录获得cDNA。逆转录条件为:37℃15 min,85℃ 5 s。按SYBR GREEN PCR Master MIX(美国ABI公司)操作说明,进行定量 PCR检测。反应条件:95℃8 min,1 Cycle;95℃ 15 s;60℃ 1 min,40 Cycles;β-actin的退火温度为60.2℃,VEGF的退火温度则为55℃。以βactin的mRNA表达水平为100%,计算Ang-2和VEGF mRNA表达水平。所测基因引物序列见表2。
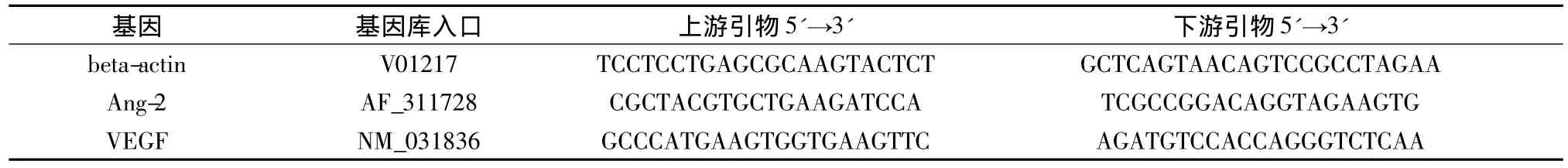
表2 扩增基因引物序列
2.5 统计分析 实验数据用SPSS 12.0统计软件统计,所有数据以均数±标准差 ()表示。组间比较 mRNA水平差异大于或等于2倍以上者,t检验,P<0.05认为有统计学意义。
3 结果
3.1 对脑缺血/再灌注大鼠神经功能评分的影响 大鼠脑缺血2 h再灌注22 h后,假手术组和正常组大鼠神经功能评分均为0分;与模型组比较,夏天无注射液中、高剂量组和尼莫地平组均可降低神经功能学评分 (P<0.05),而夏天无注射液低剂量组对神经功能评分无显著改善。且夏天无注射液中、高剂量组降低神经功能学评分作用较夏天无注射液低剂量组显著 (见表3,P<0.05)。
表3 对脑缺血/再灌注大鼠神经功能评分的影响(,n=8)
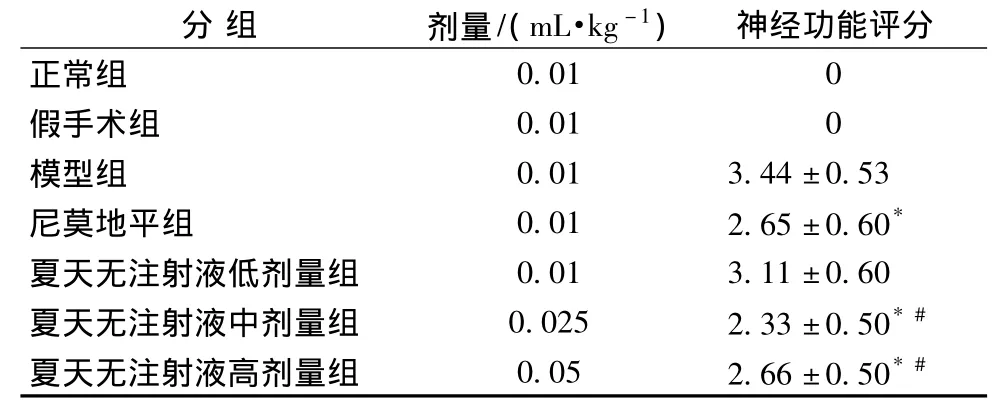
表3 对脑缺血/再灌注大鼠神经功能评分的影响(,n=8)
注:与模型组比较,*P<0.05;与夏天无注射液低剂量组比较,#P<0.05。
?
3.2 对脑缺血/再灌注大鼠脑梗死范围的影响 大鼠脑缺血2 h/再灌注22 h后,TTC染色结果显示,假手术组与正常组一样,未出现梗死灶。与模型组比较,夏天无注射液中、高剂量组和尼莫地平组均可降低脑梗死面积 (分别为P<0.05,P<0.01,P<0.05),而夏天无注射液低剂量组对脑梗死面积无显著改善。且夏天无注射液中、高剂量组降低脑梗死范围作用较夏天无注射液低剂量组显著 (见表4、图1,P<0.05)。
表4 对脑缺血/再灌注大鼠脑梗死范围的影响 (,n=8)
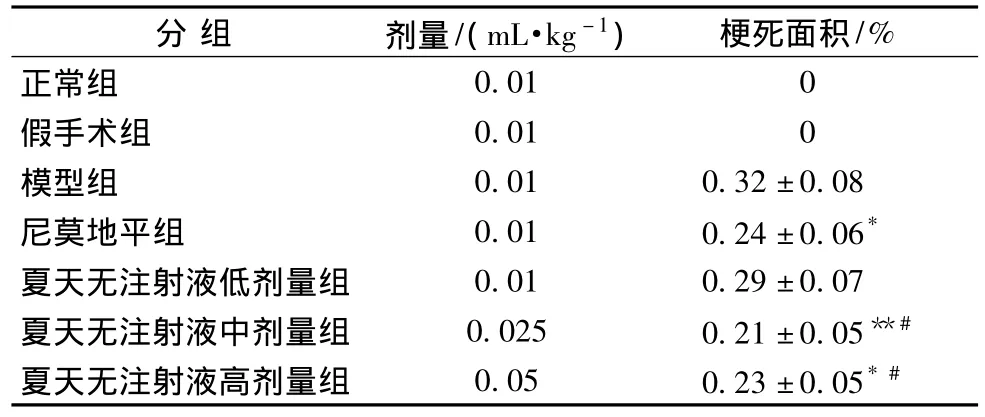
表4 对脑缺血/再灌注大鼠脑梗死范围的影响 (,n=8)
注:与模型组比较,*P<0.05,**P<0.01;与夏天无注射液低剂量组比较,#P<0.05。
分组 剂量/(mL·kg-1) 梗死面积/%正常组0.01 0假手术组 0.01 0模型组 0.01 0.32±0.08尼莫地平组 0.01 0.24±0.06*夏天无注射液低剂量组 0.01 0.29±0.07夏天无注射液中剂量组 0.025 0.21±0.05**#夏天无注射液高剂量组 0.05 0.23±0.05*#
3.3 对脑缺血2 h/再灌注22 h大鼠海马内Ang-2和VEGF mRNA水平的影响 大鼠脑缺血2 h/再灌注22 h后,与正常组及假手术组比较,模型组及夏天无注射液中、高剂量组海马组织Ang-2和VEGF mRNA均显著增高 (P<0.05)。夏天无注射液中、高剂量组与模型组比较,海马组织Ang-2 mRNA的表达进一步显著增加 (见表5,P<0.05)。

图1 夏天无注射液对脑缺血/再灌注大鼠脑梗死范围的影响 (TTC染色)
表5 对脑缺血2 h/再灌注22 h大鼠海马内Ang-2和VEGF mRNA水平的影响(,n=3)
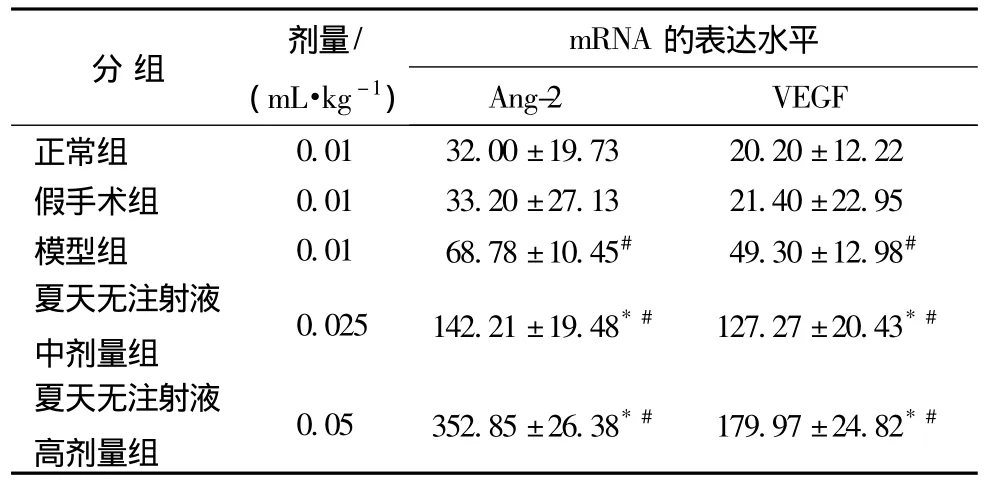
表5 对脑缺血2 h/再灌注22 h大鼠海马内Ang-2和VEGF mRNA水平的影响(,n=3)
注:与模型组比较,*P<0.05,与正常组、假手术组比较,#P<0.05。
分组 剂量/(mL·kg-1)mRNA 的表达水平Ang-2 VEGF正常组0.01 32.00±19.73 20.20±12.22假手术组 0.01 33.20±27.13 21.40±22.95模型组 0.01 68.78±10.45# 49.30±12.98#夏天无注射液中剂量组0.025 142.21±19.48*# 127.27±20.43*#夏天无注射液高剂量组0.05 352.85±26.38*# 179.97±24.82*#
4 讨论
血管的生成是一个复杂过程,受到许多正负向调节关键分子如血管生长因子和抑制血管生长因子的调控,正常生理状态下,机体内的血管一经新生即保持高度的稳定性。缺血性脑损伤后是否有血管生成是其康复的关键之一,涉及原有血管破坏、血管内皮细胞增生、出芽、新生血管形成和构建稳定的血管网络的过程。VEGF是最常见的血管生长因子之一,可作用于血管内皮细胞的特异性多功能因子,参与脑缺血后脑组织的病理修复过程[7]。脑缺血后,缺血区特别是缺血半暗带VEGF表达上调刺激了内皮细胞增生,从而形成新的血管,且VEGF贯串血管新生的整个过程,并能在内皮细胞内诱导抗凋亡蛋白,从而维持新生血管中内皮细胞的生存[8],VEGF还具有减少脑梗死面积,减轻脑水肿的作用[9-10]。
血管生成素/血管生成素受体 (Tie)通路是调节血管生成的另一重要信息途径,在血管的发生、成熟和维持血管的完整性与稳定性等方面扮演着重要角色[11-12],可减轻炎症反应和降低血管通透性等。Ang-2作为血管生成素家族中的一员,由血管内皮细胞合成,参与引发内膜去稳定、打破血管的稳定信号、导致血管消退、启动血管形成信号、促进血管形成等作用[13]。Ang-2对血管生成的影响与局部微环境有关,当VEGF存在时Ang-2则破坏血管的稳定,迅速重塑血管基底膜,激活内皮细胞增殖、侵袭、迁移、血管出芽、周细胞和平滑肌细胞聚集黏附,形成新生血管,并使血管管径增粗,通透性增加[14-15],即参与血管消退、血管形成和血管网的重建;无VEGF时,激活的内皮细胞则发生凋亡,血管发生退行性变,血管的新生受到抑制。
本研究结果显示,脑缺血/再灌注损伤后,大鼠的神经功能评分明显增高,出现较大面积的脑梗死,同时发现,海马组织的Ang-2和VEGF mRNA表达明显增高,这与文献报道一致[16-18],进一步证实在破坏血管稳定和启动新生血管形成中的Ang-2和VEGF的一致升高。值得注意的是与模型组比较,夏天无注射液治疗后,脑缺血2 h/再灌注22 h大鼠的神经功能评分明显改善,且脑梗死面积明显缩小,表明夏天无注射液对脑缺血/再灌注损伤具有明显的保护作用。此外,与模型组比较,夏天无注射液治疗组 Ang-2、VEGF mRNA的表达进一步升高,提示:夏天无注射液在促进梗死区血管消退,增强血管新生信号表达,加速血管内皮细胞增生,启动血管新生方面可能发挥着重要作用。但夏天无注射液在脑缺血/再灌注损伤修复期中是否通过血管生成从而促进损伤后修复值得进一步探讨。
[1]高 健,顾振纶,王天佑.夏天无总碱对大鼠实验性脑血栓形成的影响[J].中国血液流变学杂志,2003,13(4):325-326.
[2]胡雪勇,孙安盛,余丽梅,等.夏天无总碱抗实验性脑缺血的作用[J].中西医结合学报,2005,3(1):46-49.
[3]余丽梅,文国容,邓 江,等.夏天无注射液对小鼠脑缺血和神经元保护作用的实验研究[J].上海中医药杂志,2006,40(9):70-72.
[4]余丽梅,于 爽,龚其海,等.夏天无注射液对血管性痴呆大鼠海马血管生成素-1表达的影响 [J].中成药,2006,28(6):839-842.
[5]Yang Y R,Wang R Y,Wang P S.Early and late treadmill training after focal Brain ischemia in rats[J].Neurosci Lett,2003,339(2):91-94.
[6]Wen T C,Regido M,Gressons P,et al.A reproducible experimental model of focal cerebral ischemia in the neonatal rat[J].Brain Res Brain Res Protoc,2004,13(2):76-83.
[7]Szpak G M,Lechowicz W,Lewandowska E,et al.Border zone neovascularization in cerebral ischemic infarct[J].Folia Neuropathol,1999,37(4):264-268.
[8]Yancopoulos G D,Davis S,Gate N W,et al.Vascular-specific growth factors and blood vessel formation[J].Nature,2000,407(6801):242-248.
[9]王海东,王明荣,景 涛,等.血管内皮生长因子及其受体在食管癌中的表达和临床意义[J].中国现代医学杂志,2000,10(10):19-20.
[10]Man S M,Zhang D P,Zhang P Y,et al.Investigation of relation between plasma vascular endothelial growth factor(VEGF)and brain vascular disease[J].China J Mod Med,2004,14(10):36-39.Chinese.
[11]Sundberg C,Kowanetz M,Brown L F,et al.Stable expression of angiopoietin-l and other markers by cultured pericytes:phenotypic similarities to a subpopulation of cells in maturing vessels during later stages of anginge-nesis in vivo[J].Lab Invest.2002,82(4):387-401.
[12]Ardelt A A,McCullough L D,Korach K S,et al.Estradiol regulates angiopoietin-1 mRNA expression through estrogen receptor-alpha in a rodent experimental stroke model[J].Stroke,2005,36(2):337-341.
[13]Maisonpierne P C,Suri C,Jones P F,et al.Angiopoietin-2,a natural antagonist for Tie-2 that disrupts in vivo angiogenesis[J].Science,1997,277(5322):55-60.
[14]Lobov I B,Brooks P C,Lang R A.Angiopoietin-2 displays VEGF-dependent modulation of capillary structure and endothelial cell survival in vivo[J].Proc Natl Acad Sci USA,2002,99(17):11205-11210.
[15]Cai M,Zhang H,Hui R.Single chain Fv antibody against angiopoietin-2 inhibits VEGF-induced endothelial cell proliferation and migration in vitro[J].Biochem Biophys Res Commun,2003,309(4):946-951.
[16]Croll S D,Wiegand S J,Vascular growth factors in cerebral ischemia[J].Mol Neurobiol,2001,23(2-3):121-135.
[17]Zhang Z G,Zhang L,Tsang W,et al.Correlation of VEGF and angiopoietin expression with disruption of blood-brain barrier and Angiogenesis after focal cerebral schemia[J].J Cereb Blood Flow Metab,2002,22(4):379-392.
[18]Beck H,Acker T,Wiessner C,et al.Expression of angiopoietin-1,angiopoietin-2,and tie receptors after middle cerebral artery occlusion in the rat[J].Am J Pathol,2000,157(5):1473-1483.
