食物营养与机体健康的关系
——肠胃始动论
2012-10-27邓泽元伍国耀印遇龙
阮 征,邓泽元,伍国耀,印遇龙,3,*
(1.南昌大学生命科学与食品工程学院,江西 南昌 330031;2.德克萨斯农工大学农业与生命科学学院,美国 德克萨斯州77843-2471;3.中国科学院亚热带农业生态研究所,湖南 长沙 410125)
食物营养与机体健康的关系
——肠胃始动论
阮 征1,邓泽元1,伍国耀2,印遇龙1,3,*
(1.南昌大学生命科学与食品工程学院,江西 南昌 330031;2.德克萨斯农工大学农业与生命科学学院,美国 德克萨斯州77843-2471;3.中国科学院亚热带农业生态研究所,湖南 长沙 410125)
随着饮食结构与环境的改变,健康问题将成为21世纪人们面临的最大挑战之一。通过食物来促进身体健康,逆转亚健康状态,甚至干预或改善慢性病是调整健康状态的一条重要途径。本文综述近10年来食物成分、微生物及其代谢物、肠道结构功能三者的相互作用,以及食物与肠胃对机体其他系统的影响。认为肠胃在承载食物成分对机体健康的调控方面具有非常重要的作用:一是起始阶段的桥梁与引发作用,二是中间过程中的双向调节作用。
食品;营养;健康;肠胃
据WHO统计,目前全世界20岁及以上的成年人中有15亿人超重,其中2亿多男性和近3亿女性为肥胖。中国同样也面临着严重的健康问题。Reynolds等[1]在2000—2001年对15540名中国人进行流行病调查,以WHO标准判断男性超重率为24.1%,肥胖率为2.8%,女性超重率26.1%,肥胖率为5%[1]。肥胖成为影响健康的重大隐患,以及肥胖等因素引起的代谢综合征,如高血压与糖尿病,严重威胁人类的健康。据统计,目前我国已成为仅次于印度的全球第二大糖尿病国家,预测至2025年中国的糖尿病患者将超过3700万[2]。这些亚健康或疾病的发生不仅仅严重影响患者的身体健康和生活质量,同时也给各国造成了沉重的经济负担,因此预防或治疗慢性疾病已经成为21世纪全世界面临的最大公共卫生挑战之一,健康问题已经在全球范围内引起了广泛关注。
亚健康状态或疾病发生的机理复杂,通常认为与环境、遗传和饮食密切相关。在众多因素中,通过食物干预或改善亚健康或慢性病正逐渐引起了人们的重视。2010年,比利时科学家Szic等[3]提出食物可调节慢性病的观点。Zhao Liping等[4]提出“慢性病的肠源性假说”,认为肠道菌群是人体的第二基因组,通过改善肠道菌群,有利于慢性炎症的消除并使宿主的代谢机能逐步恢复。
消化系统(主要是肠胃)是与食物直接接触人体的第一个系统,是连接机体内环境和外部环境的桥梁,具有消化吸收功能、屏障功能、以及免疫与信号转导功能[5-7]。当前食物成分调节肠道结构和功能(如肠上皮细胞屏障[8]与黏膜免疫[9])、调节肠道微生物[10]的作用机制方面的研究较少。
本文对食物成分、微生物及其代谢物,以及肠道结构功能的三者之间的联系作用(图1),以及对机体其他组织与器官造成的影响进行总结归纳,试图找出其中的内在联系。对全面地认识食物成分、微生物和肠道对健康的影响,对食物调节与促进人体健康,逆转亚健康状态以及干预慢性病具有重要借鉴意义。
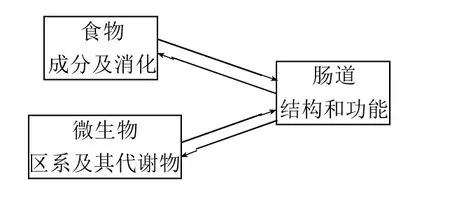
图1 肠腔内的食物、微生物和肠道三者的相互联系与作用Fig.1 Interactions of intestinal structure and function with food ingredients, microbes or their metabolites
1 食物对消化系统(肠胃)的影响
消化系统由消化道和消化腺两部分组成[11]。消化道是一条起自口腔延续为咽、食道、胃、小肠(十二指肠、空肠、回肠)、大肠(盲肠、结肠、直肠),止于肛门的一段很长的肌性管道,其中肠道是物质消化和吸收的主要场所。消化腺包括唾液腺、胰腺、肝脏、胃腺、肠腺,其主要功能是分泌消化液。
1.1 肠道的基本结构和功能
1.1.1 肠道的基本结构
典型的肠道组织结构分为黏膜层、黏膜下层、肌肉层以及外膜。黏膜层主要由上皮层、固有层和黏膜基层组成[12]。上皮层由单层柱状细胞组成(肠上皮细胞),主要包括吸收细胞、杯状细胞、潘氏细胞、内分泌细胞及未分化细胞。吸收细胞数量最多,它是由密集而规则排列的微绒毛构成,每个细胞上微绒毛约有1000根,使细胞游离面面积扩大了约20倍;杯状细胞散布于吸收细胞间,能分泌黏液,具有润滑和保护作用,从十二指肠至回肠末端其数量逐渐增多;潘氏细胞位于绒毛底部,内含溶菌酶,同时具有分泌α-防御素的能力,具有一定的灭菌作用;内分泌细胞多集中在十二指肠上部,能分泌多种激素,如L细胞广泛分布于各肠段,分泌肠高血糖素;I细胞分布多见于十二指肠和空肠,分泌缩胆囊素-促胰酶素等;未分化细胞也称隐窝干细胞,胞体较小,细胞不断的增殖、分化、向上迁移,以补充绒毛顶端脱落的吸收细胞和杯状细胞,同时也具有分化成潘氏细胞和内分泌细胞的能力;固有层中富含免疫细胞,如T细胞、B细胞、巨噬细胞,除了含有大量分散的淋巴细胞外,还含有淋巴小结,在十二指肠和空肠多为孤立淋巴小结,回肠多为集合淋巴小结。黏膜基层由内环行和外纵行两层平滑肌组成。
黏膜下层为疏松结缔组织,含有较多的血管和淋巴管;肌肉层包含内环行和外纵行两层平滑肌;外膜除十二指肠厚壁为纤维膜外,小肠其余部分的外膜均为浆膜。
1.1.2 肠道功能
肠道功能主要包括消化吸收功能、屏障和免疫作用,如图2所示[13]。目前,对肠上皮细胞的消化转运功能研究较早也较为清楚,后两者逐渐成为研究热点。肠道屏障功能和免疫作用对维持肠道稳态具有重要的意义,肠道功能正常与否与肠道黏膜层密切相关,尤其是上皮层和固有层。
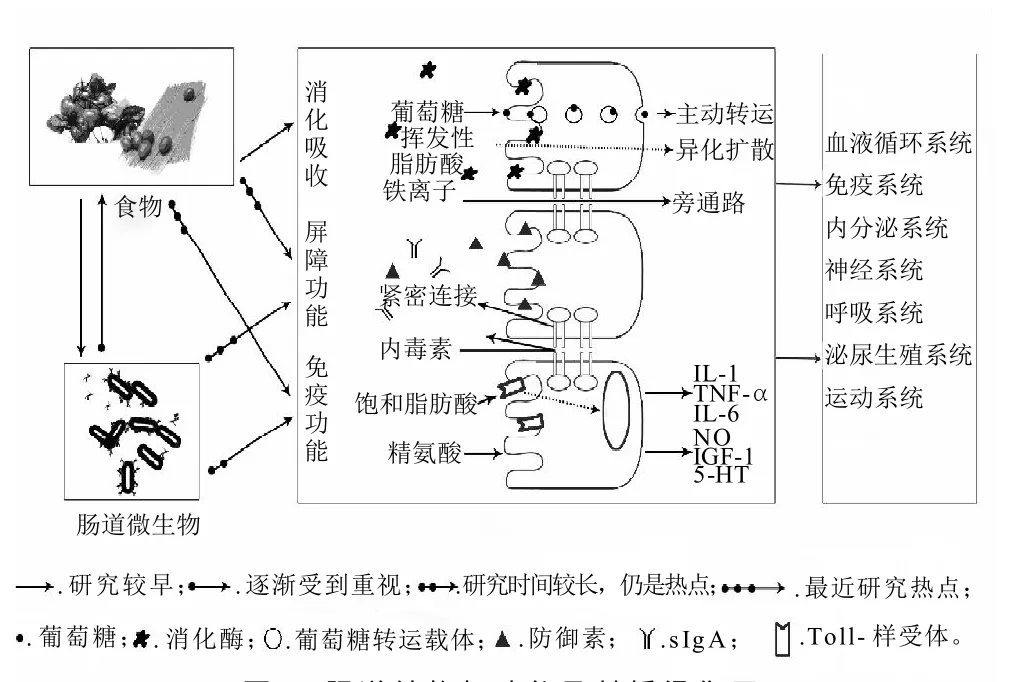
图2 肠道结构与功能及其桥梁作用Fig.2 The role of intestinal structure and function
肠道消化与吸收功能,肠上皮细胞在发挥这一功能中起着主要作用。在肠上皮细胞表面微绒毛的刷状缘上分布着大量的酶,如α-糊精酶、麦芽糖酶、蔗糖酶以及寡肽酶。胰淀粉酶的消化产物麦芽寡糖、麦芽糖以及从食物中摄入的蔗糖、乳糖等进一步在这些酶的作用下水解生成单糖,包括葡萄糖、果糖和半乳糖[14]。肠上皮细胞对营养物质的吸收可以通过多种途径:主动转运,单纯扩散,易化作用,胞吞作用[15]。
肠道的屏障功能,主要包括机械屏障、生物屏障和免疫屏障功能[16]。机械屏障主要由肠上皮细胞及其紧密连接蛋白构成,机械屏障可阻止肠腔中的大分子物质(如微生物、抗原)进入机体内部[17]。生物屏障是指与肠道相互依赖又相互作用的微生态系统平衡。肠道免疫屏障功能主要由分泌型IgA和肠上皮内淋巴细胞免疫监视构成。它能抵抗外来和自身抗原诱导免疫应答反应。这三者一起构成一个完整的肠道屏障,保护肠道免受外来的抗原和免遭异常的免疫应答反应[18]。
肠道的免疫功能由肠道黏膜免疫系统体现,它主要包括肠道黏膜上皮及固有层内呈弥散分布的淋巴细胞,以及组织化的淋巴组织(如派氏结、肠系膜淋巴结及较小的孤立淋巴滤泡)[19]。成人肠道黏膜面积至少有200m2,肠道内环境非常复杂,使肠道黏膜免疫系统长期曝露于各种病原体、食物和肠道共生菌群,因此肠道黏膜免疫系统形成了一套严格的调节机制区分各种抗原刺激并进行分类并迅速做出反应:对无害抗原(食物营养成分和有益菌种)能下调免疫应答,具有免疫耐受特性;而对有害抗原则产生体液和细胞免疫并排除抗原[20]。肠道黏膜免疫在调节肠道免疫耐受和免疫反应平衡扮演重要角色。
此外,肠上皮细胞还具有信号识别与转导的功能,部分参与免疫作用。肠上皮细胞表面表达Toll样受体(TLRs),识别肠腔内的抗原[21]。在正常情况下,细胞表面TLRs低水平表达,只有在疾病状态(炎症性肠病)下其表达显著增加,这在一定程度上增加了对肠道共栖微生物的耐受性。同时肠上皮细胞通过分泌的胸腺基质淋巴细胞生成素(TSLP)与T细胞上相应的受体结合来调节辅助型T细胞的分化,TSLP还能通过影响VA的代谢调节DCs细胞的功能[22],揭示肠上皮细胞可参与肠道黏膜免疫的调控[23]。
1.2 食物成分与肠道结构和功能的相互作用
1.2.1 食物成分对肠道结构和功能的影响
食物成分及其代谢物可影响肠道消化和吸收功能。日粮中添加谷氨酰胺可以缓解肠道中乳糖酶的下降,显著增加回肠中亮氨酰氨基肽酶,降低十二指肠中碱性磷酸酶[24],提高肠道对木糖的吸收功能[25]。丁酸盐能提高回肠蔗糖酶的活性[26]。
食物成分及其代谢物亦可影响肠道屏障功能。1)膳食成分可调节肠道屏障。Li Nan等[27]研究成果表明谷氨酰胺可通过活化磷酸肌醇3激酶(phosphatidylinositol 3-kinase,PI3K),增加肠上皮细胞Caco-2屏障功能。精氨酸是哺乳期的一种必需氨基酸,在维持和保护肠道黏膜、促进肠道损伤修复等方面发挥重要作用,精氨酸干预LPS诱导的猪肠上皮细胞损伤,发现精氨酸能促进猪肠上皮细胞的增殖,揭示精氨酸能促进肠道损伤的修复[28]。2)食物成分转化物及其消化吸收过程影响肠道屏障功能的完整。在营养物质的消化吸收过程中伴随着能量代谢,能量代谢过程产生的自由基能损伤肠道屏障。王艳艳[29]研究显示高脂膳食喂养的小鼠回肠自由基增加了1倍,同时伴随抗氧化酶活力的降低。自由基诱导酪氨酸的磷酸化能降低紧密连接蛋白的表达,增加肠道通透性[30]。3)参与物质代谢的酶可调节肠道屏障功能。腺苷酸活化蛋白激酶(AMP-kinase,AMPK)在脂质代谢,能量代谢及葡萄糖代谢中发挥着重要作用,近年研究显示AMPK的活化能调节肠上皮细胞紧密连接蛋白的分布,增强肠上皮细胞屏障功能[31]。
食物成分及其代谢物可调控肠道免疫功能。Lindsay等[32]对中度结肠炎患者给予低聚果糖干预3周,患者结肠中IL-10阳性树突状细胞的比例由30%增加到53%,同时表达TLR2和TLR4树突细胞显著增加。多糖类物质在肠道菌群的作用下代谢产物丁酸对肠道具有免疫调节作用,Segain等[33]用丁酸与来自结肠炎患者肠道固有层淋巴细胞和外周血单核细胞共同培养,发现丁酸能显著抑制NF-κB的活化和IκBα的降解,抑制肿瘤坏死因子-α,白介素-6、白介素-1β促炎基因表达以及肿瘤坏死因子-α的分泌。VA代谢产物维甲酸可通过T淋巴细胞表面得维甲酸孤儿素受体调节T淋巴细胞的分化[34]。
除此之外,食物成分及其代谢物对肠道的抗氧化系统也造成影响。Lee等[35]将大豆皂苷干预结肠炎小鼠发现,大豆皂苷I能显著提高结肠谷胱甘肽的含量和超氧化物歧化酶、过氧化氢酶的活力,并降低丙二醛的含量。Jaganath等[36]将芦丁与人粪便在厌氧条件下共同孵育,经过肠道菌群代谢作用后,发现芦丁的代谢产物的抗氧化能力和发酵时间和代谢物的积累量相关,同时不同个体来源的粪便显示的对芦丁的代谢能力不同,抗氧化性也不同,其代谢产物槲皮素和3,4-二羟基苯乙酸对抗氧化能力显著强于芦丁,而3-羟基苯乙酸、4-羟基苯甲酸抗氧化能力几乎为零。这表明不同个体间菌群的差异能导致营养成分代谢的差异,同时营养成分在肠道内代谢物的种类影响肠道内抗氧化能力。
综上,食物成分及其代谢物对肠道结构和功能具有重要的影响,对肠道形态、肠道消化酶的活力、肠上皮细胞间的紧密连接、肠道免疫功能和抗氧化能力均能产生影响。
1.2.2 肠道功能和结构对食物成分及其代谢物的影响
肠上皮细胞表面含有丰富的酶类以及转运载体参与营养物质的消化和吸收。机体对葡萄糖的吸收可以通过肠上皮细胞膜上葡萄糖转运载体(GLUT)[37];黄酮苷元 (如槲皮素) 可通过转运载体如钠离子依赖葡萄糖转运载体而以原型形式被吸收[38];钙、铁等一些矿物质主要通过肠黏膜上的钙结合蛋白而主动转运,部分通过上皮细胞间紧密连接而被吸收[39]。
1.3 食物成分和微生物的相互作用
成人肠道内定殖的细菌细胞数量多达1014个,种类约500~1000种,其数量约为人体细胞的10倍,所编码的基因数至少是人体自身基因的100倍[40]。这些种类与数量繁多的微生物组成一个复杂的微生态系统,直接参与人体的消化、物质代谢以及免疫调节。同时食物成分对微生物菌群具有调节作用。
1.3.1 食物成分对微生物菌群的影响
由于食物成分是肠道细菌发酵的主要底物,因此膳食结构和食物的成分和含量在很大程度上能影响肠道菌群的组成及其代谢。Finegold等[41]比较传统日式饮食和西方饮食的日裔美国人肠道菌群的组成时发现,虽然他们具有相似的遗传背景,但肠道菌群组成却显著差异,日式饮食下除肠杆菌以外的兼性厌氧菌和需氧菌的数量较西方组显著增多。喂养高脂的小鼠肠道内产丁酸盐的细菌增多,小鼠容易肥胖[42]。Cani等[43]发现高脂饮食使革兰氏阴性菌的数目增加,双歧杆菌减少。Bouhnik等[44]对20个健康人每天分别用12.5g果寡糖和蔗糖(每组10人),果寡糖组中人肠道双歧杆菌明显提高。壳聚糖干预糖尿病大鼠能显著降低大肠肝菌和肠球菌的数量,对双歧杆菌和乳酸杆菌的增殖作用具有显著增殖作用[45]。Lee等[46]对肠道菌群代谢茶多酚研究发现,茶多酚及其代谢物对致病菌的生长具有抑制作用,如3-甲氧基没食子酸和没食子酸对产气荚膜梭菌的生长,而对有益菌的生长影响不大。
1.3.2 微生物对食物成分的代谢作用的影响
肠道菌群是人体不可分割的组成部分,它通过自身的酶直接参与宿主的代谢过程。肠道细菌可以代谢机体自身不能代谢的化合物。研究表明至少能产生的156种碳水化合物活化酶,其中77种糖苷水解酶、35种糖基转移酶、12种多糖裂解酶、11种糖酯酶。它们降解机体自身不能降解的多糖,比如木聚糖、果胶、阿拉伯糖等,其代谢终产物为短链脂肪酸[47]。1)微生物作用于不同食物成分产生的短链脂肪酸的种类不同。支链氨基酸(如缬氨酸、亮氨酸)发酵产生支异丁酸、异戊酸等之链脂肪酸[48],而芳香族氨基酸(如苯丙氨酸、色氨酸)发酵则产生酚类和吲哚类物质[49]。2)微生物作用于不同的食物成分产生短链脂肪酸的量不同。抗性淀粉比淀粉产生的乙酸多,而淀粉发酵则产生较多的丁酸[50]。Moreau等[51]用低聚乳果糖和抗性淀粉分别喂葡聚糖硫酸钠诱导结肠炎小鼠,低聚乳果糖组丁酸的产量要优于抗性淀粉。此外,肠道菌群还参与宿主的代谢,在肠道细菌的作用下将牛磺酸和甘氨酸结合的结合型胆汁酸降解,释放出胆汁酸以及牛磺酸[52]。
同时肠道细菌对食物营养成分的转化作用将影响到其生物功能的发挥,既能发挥有益作用又能产生有害影响。越来越多的研究表明食物成分经过肠道菌群后的代谢产物是其发挥生理活性的主要物质。无菌小鼠体内不能正常代谢多酚类物质,表明肠道菌群在黄酮类物质的代谢过程中起着重要的作用[53]。Rwoland等[54]将大豆黄酮与人粪便在厌氧环境下共同发酵培养,结果发现大豆黄酮被人体肠道细菌转化为生物活性更高的牛尿酚。人参皂苷被认为是人参的主要活性成分,研究发现人体肠道内菌群转化人参皂苷的代谢产物Compound K 是非天然的人参皂苷(由二醇型人参皂苷转化而来),被认为是真正被人体吸收和发挥活性作用的实体[55]。Tsuchihashi等[56]将葛花苷与人肠道内中的不同菌株共同培养后发现葛花苷的代谢物不同,与链状双歧杆菌和假链状双歧杆菌菌株共同培养后,主要代谢产物为尼泊尔莺尾异黄酮;而与消化链球菌属菌株共同培养后,主要代谢产物则为6-羟基鹰嘴豆素A。白雪[57]发现葛花苷在大鼠体内代谢产物尼泊尔莺尾异黄酮对降低小鼠醉酒率和延长睡眠潜伏期时间优于野葛花水提物和葛花苷。与之相反,肠道微生物对食物成分的代谢作用还可能不利于宿主健康,如糖精(cyclamate,环己氨基磺酸盐)本身对人体无害,可在尿液中却发现了一种可能的潜在致癌物环己胺(cyclohexylamine),研究发现宿主细胞不能实现这种转化,当其与粪便共同孵育时即可发生这种转化[58]。合成维生素也是肠道菌群对物质代谢的影响之一。研究表明肠道菌群可以合成多种人体所需的维生素,包括VB1、VB2、VB12、VK、尼克酸和叶酸等[59-60]。
综上所述,肠道微生物在机体的代谢过程中发挥着重要作用,首先它能代谢机体(肠胃)不能代谢的食物成分进行辅助机体代谢。其次还可参与机体代谢,其对营养物质的转化作用对宿主将产生有益或有害的作用,比如Nicholson等[61]认为人体的代谢是由自身基因和肠道共生的微生物共同作用的结果。
1.4 微生物对肠道结构和功能的影响
肠道菌群结构失衡会影响肠道机械屏障的变化。抗生素能破坏肠道菌群平衡,机会致病菌C.difficile相对比例升高,而其产生的毒素增加肠上皮细胞通透性而导致肠道细菌移位[62]。Brun等[63]发现肥胖小鼠肠道通透性比正常小鼠高,且血液中炎症因子显著增高,并认为肠道通透性的增加导致内毒素血症,最终发生炎症性损伤。与此相反,有益菌的增加对肠道屏障功能具有保护作用。Cani等[43]给高脂膳食诱导的糖尿病小鼠饲喂果聚糖,肠道双歧杆菌的数量增加,血中内毒素降低,通过对肠道菌群与血内毒素进行相关性分析发现双歧杆菌与内毒素的含量呈现显著负相关,因此揭示双歧杆菌数量的增加能保护肠道的通透性。将益生菌用于结肠炎动物模型与结肠炎患者,发现益生菌对促进疾病的改善有较好的效果[64-66],Zocco等[67]用Lactobacillus GG干预溃疡性结肠炎患者,通过临床观察,内窥镜以及肠道组织学观察发现该乳酸菌对疾病的缓解具有显著作用,与临床用药5-氨基水杨酸效果无显著差异。
肠道菌群不仅仅能影响肠道的屏障功能,同时还被认为是肠道免疫功能的“塑造者”[68]。在无菌动物体内肠道免疫功能受到严重影响,如肠相关淋巴组织发育受损,派伊尔淋巴结和肠系膜淋巴结数量减少,孤立的淋巴滤泡的成熟受阻,同时免疫细胞(如C D 4、 CD8 T淋巴细胞数量减少,免疫细胞表达的受体减少(MHC-Ⅱ、TLR9)表达减少,抗菌活性物质(如IgA)分泌减少,这揭示了肠道菌群对肠道免疫功能的形成产生重要的影响[69-71]。Toll样受体是一类重要的细胞表面模式识别受体,能广泛的识别肠道菌群不管是有益菌还是有害菌携带的病原体相关分子模式,因此Abreu[72]认为肠道功能的建立与肠上皮细胞表达的TLRs对肠道菌群的识别有关。
在宿主肠上皮细胞的黏膜上有黏附素受体,其中大部分受体具有脂肪酸结合位点的蛋白质或糖脂;同时在肠道细菌表面有不同黏附结构的配体,这为肠道细菌在肠上皮细胞黏膜上黏附和定殖提供条件。研究表明双歧杆菌菌体表面的脂磷壁酸是一种黏附素,能介导双歧杆菌在肠上皮细胞的黏膜上黏附[73]。一般认为只有活菌才具有黏附能力,Fourniat等[74]发现死乳杆菌也具有黏附能力。有益菌在肠道的定殖具有重要的意义,定殖有助于形成稳定的膜菌群,抑制有害菌在肠上皮细胞表面的黏附。
综上所述,肠道中食物成分、微生物以及肠道三者间的相互影响,共同维持着肠道健康。食物成分可直接影响肠道结构和功能,也可影响肠道微生物区系及其代谢产物,并可经由肠道菌群来调控肠道结构和功能,因而食物在这三者中的地位显得尤为重要。近年来随着对肠道功能研究的不断深入,肠-肝轴、肠-胰岛轴、肠-脑轴等概念的相继提出,对肠道功能的认识有了进一步的认识,同时它在维持人体健康方面起到的重要作用引起了广泛的关注和研究热情。本文将进一步探讨食物成分对机体各个系统的影响,如图3所示,旨在揭示肠胃结构功能在维持机体健康中所起的重要作用。
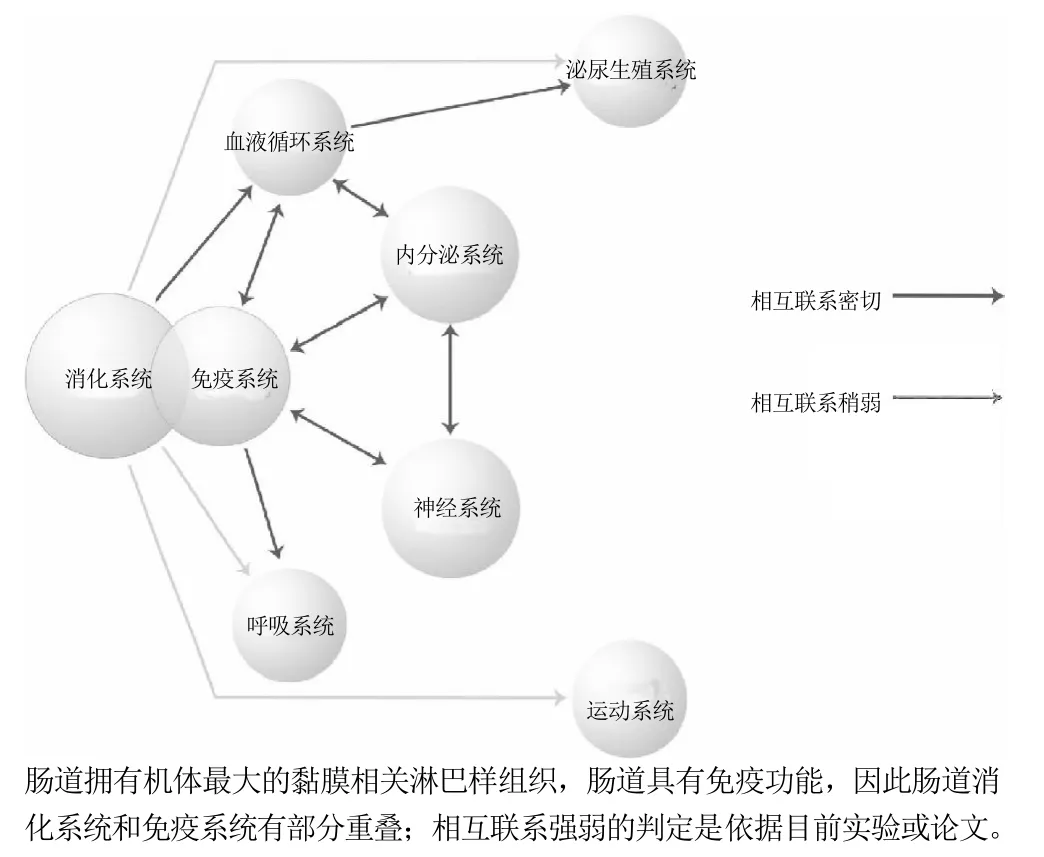
图3 消化系统与其他系统的影响Fig.3 Effect of gastrointestine (digestive system) on other organs and tissues
2 肠胃与免疫的相互作用
在肥胖,糖尿病与高血压等慢性疾病的患者体内发现一个共同特征[75-76]:体内持续的、低水平的慢性炎症,因此近年来,机体的代谢与免疫的关系受到广泛的关注,《Trends in Immunology》[77]、《Nature Reviews immunology》[78]、《Nature》[79-80]等杂志上均发表多篇文章[81-82],试图理清两者之间的关系。
食物中的膳食成分如脂质(脂肪酸、胆固醇、脂溶性维生素等)、葡萄糖、微量元素(锌、铜、铁等)都对免疫系统有重大的影响。同时参与免疫过程的模式识别受体和细胞因子等又能影响食物成分的吸收代谢过程[78,81]。深入了解两者之间的关系为通过膳食干预调理人体生理状态,维持内环境的稳定,提高免疫力提供理论支持。
2.1 食物成分及其代谢物对免疫的调控
食物过敏是食物引起免疫反应最典型的症状之一。如花生、牛奶引起由IgE介导的免疫反应;小麦、黑麦等谷物能引起对醇溶谷蛋白及其相关蛋白不耐受的个体产生乳糜泻,当患者进食这些致敏性谷物或蛋白后引起小肠黏膜的损伤,阻碍营养物质的吸收并产生炎症[83]。
脂肪是人体的六大营养素之一,近年研究表明它还参与到免疫调节的过程。高脂膳食引起肠上皮细胞通透性增加,从而导致血内毒素含量的增加[30,43,84],而内毒素在血液循环过程中的增加是导致系统性的慢性炎症的重要原因,也是引起肥胖及相关的代谢综合征产生的主要因素[85]。Ghanim等[86]研究发现食用高脂高碳水化合物的膳食相对于高碳水化合物的膳食能显著增加血单核细胞TLR4、TLR2的表达以及核转录因子κB(NF-κB)的活化,提示高脂是导致TLRs在单核细胞表面增加的原因。同时研究表明,饱和脂肪酸能诱导TLR4、TLR2基因的表达,并可通过Myd88依赖途径和非依赖途径活化NF-κB,从而分泌大量的炎性因子;而不饱和脂肪酸却能抑制TLRs基因的表达及其信号通路的活化[87]。
氨基酸也同样被认为具有免疫调节作用。其中对精氨酸的研究比较多,精氨酸是幼年动物必须氨基酸之一,对维持肠道健康具有重要作用。研究表明,补充精氨酸能调节T淋巴细胞活性、增加自然杀伤细胞和巨噬细胞的活力,从而增强机体免疫力功能[88]。对断奶仔猪灌喂精氨酸,能增加血清IgM、IgG的含量,增强脾脏IL-8、TNF-α基因的表达,从而增强了仔猪的免疫力[89]。
VA的代谢物维甲酸对免疫系统的具有重要的影响[34,90],它不仅对T淋巴细胞和B淋巴细胞的功能具有调节作用,同时还影响树突状细胞的功能。Mucida等[91]揭示维甲酸能抑制T细胞上的维甲酸孤儿素受体的表达,有利于诱导Th0细胞向Treg细胞极化。维甲酸还能通过DCs表面相应的受体途径调节细胞因子的产生,能促进Th2型细胞因子:IL-4、IL-5、IL-6[92]。Hsu等[93]将大豆和绿茶联合使用干预患有前列腺癌的大鼠,实验结果表明它们的联合使用能抑制前列腺组织NF-κB的活化,降低炎性因子浸润和促炎因子的分泌,同时实验结果还表明,绿茶和大豆蛋白单独使用效果不明显。
2.2 免疫对食物吸收和代谢的影响
免疫细胞的活化及其功能的维持需要大量的能量,而葡萄糖是主要的能量来源。免疫细胞被抗原激活后,对葡萄糖的需求量激增,此时可通过两个途径增加对葡萄糖的吸收。首先,增加免疫细胞细胞膜上葡萄糖转运载体蛋白(GLUT)的表达;其次,免疫细胞激活后引起的信号通路也能增加对葡萄糖的摄入,如T细胞活化后,其表面的协同刺激分子受体CD28刺激PI3K-AKT信号通路的活化,增加对葡萄糖的摄入[77,94]。Healy等[95]认为胞内增加葡萄糖浓度可以抑制白细胞凋亡,这种保护性作用与细胞利用葡萄糖的程度有关。CD28被认为是具有免疫和代谢双重作用的代表性分子之一。
除此之外,免疫过程中分泌的细胞因子也参与到了物质的代谢过程。高水平的IL-6增加血清甘油三酯以及血糖的浓度,而IL-6基因缺失的小鼠对葡萄糖的耐受下降[96];Plomgaard等[97]研究显示TNF-α在血液中的激增,使脂解作用增强,同时增加游了离脂肪酸的含量。然而细胞因子对代谢的具体作用机制还不明确。除细胞因子外,黏附因子也被认为参与到机体的代谢过程。Dong等[98]通过对ICAM-1基因缺失的小鼠研究发现,在正常饮食的情况下ICAM-1基因缺失的小鼠体质量显著高于正常小鼠,白色脂肪(P<0.01)与褐色脂肪(P<0.05)占体质量比例显著升高;在高脂膳食饲喂2周后,基因缺失小鼠与正常小鼠采食量无显著差异,但体质量却显著高于正常小鼠,同时也影响到脂肪沉积。
3 肠胃与血液循环系统的相互作用
由于小肠是食物消化和营养物质吸收的主要场所,因此小肠肠腔内物质的组分以及肠上皮细胞完整性将直接影响到血液循环系统。
3.1 食物成分及其代谢物对血液生物化学组分的影响
Toufektsian等[99]发现花青苷增加大鼠血清中n-3多不饱和脂肪酸(如EPA与DHA)的含量。苦瓜具有多种生物活性物质,其中苦瓜多糖被称为植物胰岛素,董英等[100]通过对糖尿病小鼠饲喂水提取和碱提取的苦瓜多糖,实验结果表明苦瓜多糖能显著降低血中葡萄糖的含量。
当肠上皮细胞的屏障功能受到损害时,血液中脂多糖的含量的显著升高[76],而脂多糖是革兰氏阴性菌细胞壁的主要成分,能刺激免疫细胞活化并引起促炎因子分泌增加,促炎因子通过循环在全身蔓延将引起全身性的炎症[85],因此肠道屏障功能完善与否与糖尿病等慢性疾病密切相关[101]。由脂多糖入血引起的疾病最典型的是脓毒素血症,张晗等[102]对脓毒血症大鼠研究中发现血碱性磷酸酶的含量与疾病的严重程度负相关。大量研究表明高脂膳食是增加肠道通透性的主要原因之一。Cani等[43]用高脂膳食喂养小鼠,肠道的通透性显著增加,导致血液中脂多糖的浓度显著升高,同时血液中促炎因子IL-1α、IL-1β、IL-6的含量也显著高于普通小鼠。Li Qiurong等[103]也证实,n-3多不饱和脂肪酸在促炎因子诱导的通透性缺陷和上皮屏障功能紊乱中,能通过调整紧密连接的膜微观区域,增加肠道上皮细胞的紧密性。刘玉江等[104]用膳食纤维干预乙酸灌肠法制作大鼠炎症性肠病疾病模型时也证实,添加膳食纤维的肠内营养能降低门静脉血中的脂多糖水平、促进肠道黏膜的修复、加强肠黏膜屏障的保护作用。
3.2 食物成分及其代谢物对血管壁的影响
大豆中含有丰富的大豆肽,经酶解后能产生血管紧张素转化酶抑制剂(ACEⅠ),能够降低血压。汤健等[105]发现大豆肽对自发性高血压降压效果明显。丁志山等[106]对姜黄素抗肿瘤的研究过程中,发现姜黄素能抑制血管生成。增加膳食中脂肪的摄入量,容易引起脂肪在血管壁上沉积,引起动脉粥样硬化等疾病。大量证据表明炎症在动脉粥样硬化发生、发展和演变过程中起着重要作用,炎症是动脉粥样硬化性疾病的基本特征,同时亦是动脉粥样硬化性疾病的始动因子。
4 肠胃对呼吸系统的影响
通过对炎症性肠病的研究发现,约超过50%的肠炎患者有不同程度的呼吸系统症状,如支气管炎、支气管扩张、肺炎等[107-108],对于肠炎患者会患呼吸系统疾病的原因知之甚少。肠缺血再灌注损伤启动炎症应答可导致远隔器官的损害,甚至多器官功能衰竭,其中以急性肺损伤(acute lung injury,ALI)引起的呼吸窘迫综合征(acute respiratory distress syndrome,ARDS)最为突出。发病机制错综复杂,迄今尚未完全阐明,但本质是炎症反应的失控[109]。
刘辉[110]用VA干预难治性哮喘取得了一定的效果,其对疾病的缓解作用可能依赖于VA代谢产物维甲酸对免疫的调控作用。姜黄素是广泛存在于香辛料中了多酚类物质,在小肠黏膜中被代谢,具有多种生理功能,如提高抗氧化能力,抑制NF-κB的活化等,同时还可以通过调节组蛋白去酰基化酶调控肺上皮细胞促炎因子基因的活化[111]。芬兰的一项对超过10000人群调查,发现多酚的摄入量与哮喘的发病率呈负相关[112],Walda等[113]一项超过13000个志愿者对慢性梗阻性肺疾发病率的研究结果也显示出相关的变化。
5 肠胃对神经-内分泌系统的影响
食物成分及其代谢产物对神经-内分泌系统的影响主要有两条途径:一是直接影响,另一个是通过免疫系统间接影响。大豆是一种优良的植物蛋白,动物实验研究结果表明,饲喂大豆蛋白后能改变动物体内激素浓度,特别是甲状腺素、胰岛素、升血糖激素等激素与胆固醇的代谢密切相关。Meja等[114]研究认为姜黄素通过上调组蛋白去酰基化酶的表达来恢复糖皮质激素的功能。胃肠道黏膜中的嗜铬细胞,在进食或者病理性刺激的时候刺激嗜铬细胞分泌5-羟色胺,5-羟色胺是一种神经递质,在肠至脑的信息传递过程中起着重要作用[115]。
同时鉴于食物成分对免疫的重要调节作用,免疫-神经-内分泌系统间的紧密的相互联系与作用,因此,肠胃结构与功能对维持正常的生理功能和内环境稳定起着重要作用。IL-1是由免疫细胞分泌的细胞因子,研究表明,它能刺激神经肽的分泌,比如促进促肾上腺激素释放激素(CRH)、黑皮质素[116]。TNF-α在众多疾病中均发现显著增加,它在肥胖诱导的糖尿病的发展过程中起着重要作用,研究显示它能降低胰岛素受体对胰岛素的敏感性[117-118]。α-黑素细胞刺激素(α-MSH)由下丘脑神经元前阿片黑素细胞皮质激素产生,与黑皮质素4受体(MC4R)结合后,抑制食欲,增加基础代谢率[119]。
6 肠道始动论
近几十年来随着生活环境、生活方式与食物结构的改变,全球肥胖、Ⅱ-型糖尿病、心血管疾病的发病率呈现急剧上升趋势。美国华盛顿大学Gordon研究组提出将“肠道菌群作为一种环境因素条件脂肪贮存”的观点[120],提倡人们应该关注肠道,关注肠道微生物。英国帝国大学Nicholson教授研究组对中国、美国、日本和英国的17个不同地区的4360名志愿者尿液代谢组学分析,发现高血压与肠道菌群的组成有密切关系[121]。揭示肠道在亚健康或疾病的发生与发展过程中具有举足轻重的地位。
肠胃不仅作为机体对外来营养物质消化和吸收的主要场所,还具有屏蔽或清除外来有害物质与微生物的功能。肠胃为机体的健康输送有益的物质,体现了作为食物与机体健康之间的桥梁作用,也体现了作为机体系统中的起始位置,谓之始。食物对肠道微生物的调节作用、肠道菌群对食物成分的代谢作用、食物与肠道微生物调节肠道结构与功能以及肠道屏障功能是否完整、肠道黏膜免疫是否平衡等均能影响机体的健康。肠胃在调节机体健康过程中体现了推动(正向调节)与拉动(负向调节)的作用,谓之动。
肠道始动论的提出旨在更加关注在肠胃空间中食物成分、微生物和肠道结构功能三者之间的作用机制和物质基础变化,以及肠道结构功能调控机体健康的作用途经。多角度、多层次的深入研究将为通过食物干预调控亚健康状态和预防疾病供理论依据,将对人类健康与生活质量产生重要影响。
[1]REYNOLDS K, GU D F, WHELTON P K, et al. Prevalence and risk factors of overweight and obesity in China[J]. Obesity(Silver Spring), 2007, 15(1): 10-18.
[2]KING H, AUBERT R E, HERMAN W H. Global burden of diabetes, 1995—2025: prevalence, numerical estimates, and projections[J]. Diabetes Care, 1998, 21(9): 1414-1431.
[3]SZIC K S V, NDLOVU N M, HAEGEMAN G, et al. Nature or nurture: let food be your epigenetic medicine in chronic inflammatory disorders [J]. Biochemical Pharmacology, 2010, 80(12): 1816-1832.
[4]ZHAO Liping, SHEN Jian. Whole-body systems approaches for gut microbiota-targeted, preventive healthcare[J]. J Biotechnology, 2010, 149(3): 183-190.
[5]BRANDTZAEG P. Development and basic mechanisms of human gut immunity[J]. Nutrition Reviews, 1998, 56(Suppl 1): 5-18.
[6]GROSCHWITZ K R, HOGAN S P. Intestinal barrier function: molecular regulation and disease pathogenesis[J]. J Allergy Clin Immunol, 2009, 124(1): 3-20.
[7]UEMATSU S, FUJIMOTO K, JANG M H, et al. Regulation of humoral and cellular gut immunity by lamina propria dendritic cells expressing Toll-like receptor 5[J]. Nature Immunology, 2008, 9(7): 769-776.
[8]刘琳娜. 大黄多糖对肠上皮细胞损伤的修复作用及其机制研究[D].西安: 第四军医大学, 2005.
[9]HINO K, KUROSE M, SAKURAI T, et al. Effect of dietary lactosucrose (4G-β-D-galactosylsucrose) on the intestinal immune functions in mice [J]. Journal of Applied Glycoscience, 2007, 54(3): 169-172.
[10]LAPARRA J M, SANZ Y. Interactions of gut microbiota with functional food components and nutraceuticals[J]. Pharmacological Research, 2010, 61(3): 219-225.
[11]王芬, 左明雪. 人体及动物生理学[M]. 2版. 北京: 高等教育出版社, 2001: 277-314.
[12]王兴鹏. 肠道屏障功能障碍: 基础与临床[M]. 上海: 第二军医大学出版社, 2006: 8-11
[13]SHIMIZU M. Interaction between food substances and the intestinal epithelium[J]. Biosci Biotechnol Biochem, 2010, 74(2): 232-241.
[14]秦环龙. 肠道屏障功能的基础与临床[M]. 上海: 上海交通大学出版社, 2007: 9-10.
[15]WOUDSTRA T, THOMSON A B R. Nutrient absorption and intestinal adaptation with aging[J]. Beat Practice &Research Clinical Gastroenterology, 2002, 16(1): 1-25.
[16]段建华, 晏春根. 肠黏膜屏障研究进展[J]. 实用医学杂志, 2004, 20 (2): 212-214.
[17]TSUKITA S, FURUSE M, ITOH M. Multifunctional strands in tight junctions[J]. Molecular Cell Biology, 2001, 2: 285-293.
[18]李伟, 陈庆森. 肠道黏膜免疫屏障及其菌群与机体健康关系的研究进展[J]. 食品科学, 2008, 29(10): 649-655.
[19]艾国平. 肠道粘膜免疫的构成与功能[J]. 免疫学杂志, 2000, 16(增刊4): 82-84.
[20]MOWAT A M. Anatomical basis of tolerance and immunity to intestinal antigens[J]. Nature Reviews Immunology, 2003, 3(4): 331-341.
[21]LEE J, MO J H, SHEN C, et al. Toll-like receptor signaling in intestinal epithelial cells contributes to colonic homoeostasis[J]. Current Opinion in Gastroenterology, 2007, 23(1): 27-31
[22]ARTIS D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut[J]. Nature Reviews Immunology, 2008, 8(6): 411-420.
[23]RIMOLDI M, CHIEPPA M, SALUCCI V, et al. Intestinal immune homeostasis is regulated by the crosstalk between epithelial cells and dendritic cells[J]. Nature Immunology, 2005, 6(5): 507-514.
[24]张军民. 谷氨酰胺对早期断奶仔猪肠道的保护作用及其机理研究[D].北京: 中国农业科学院, 2000.
[25]HSU C B, HUANG H J, WANG C H, et al. The effect of glutamine supplement on small intestinal morphology and xylose absorptive ability of weaned piglets[J]. African Journal of Biotechnology, 2010, 9(41): 7003-7008.
[26]LAUS R, GUNTHNER D, LETZGU H. Effects of feeding fat-coated butyrate on mucosal morphology and function in the small intestine of the pig[J]. J of Animal Physiology and Animal Nutrition, 2007, 91(7/8): 312-318.
[27]LI Nan, NEU J. Glutamine deprivation alters intestinal tight junctions via a PI3-K/Akt mediated pathway in Caco-2 Cells[J]. Journal of Nutrition, 2009, 139(4): 710-714.
[28]TAN Bie, YIN Yulong, KONG Xiangfeng, et al. L-arginine stimulates proliferation and prevents endotoxin-induced death of intestinal cells[J]. Amino Acids, 2010, 38(4): 1227-1235.
[29]王艳艳. 高脂膳食诱导小鼠氧化应激及脂代谢相关基因表达的影响[D]. 无锡: 江南大学, 2008.
[30]RAO R K, BASUROY S, RAO V U, KARNAKY K J, et al. Tyrosine phosphorylation and dissociation of occludin-ZO-1 and E-cadherin-βcatenin complexes from the cytoskeleton by oxidative stress[J]. Biochemistry Journal, 2002, 368(Pt2): 471-481.
[31]ZHENG Bin, CANTLEY L C. Regulation of epithelial tight junction assembly and disassembly by AMP-activated protein kinase[J]. PNAS, 2007, 104(3): 819-822.
[32]LINDSAY J O, WHELAN K, STAGG A J, et al. Clinical, microbiological, and immunological effects of fructo-oligosaccharide in patients with Crohn,s disease[J]. Gut, 2006, 55(3): 348-355.
[33]SEGAIN J P, de la BLETIERE D R, BOURREILLE A, et al. Butyrate inhibits inflammatory responses through NF-κB inhibition: implications for Crohn,s disease[J]. Gut, 2000, 47(3): 397-403.
[34]MORA J R, IWATA M, von ANDRIAN H U. Vitamin effects on the immune system: vitamins A and D take centre stage[J]. Nature Reviews Immunology, 2008, 8(9): 685-698.
[35]LEE I A, PARK Y J, YEO H K, et al. Soyasaponin I attenuates TNBS-induced colitis in mice by inhibiting NF-κB pathway[J]. J Agric Food Chem, 2010, 58(20): 10929-10934.
[36]JAGANATH I B, WILLIAM M, LEAN M E J, et al. in vitro catabolism of rutin by human fecal bacteria and the antioxidant capacity of its catabolites[J]. Free Radical Biology & Medicine, 2009, 47(8): 1180-1189.
[37]高磊, 刘绛光. 对葡萄糖转运蛋白的讨论[J]. 国外医学: 临床生物化学与检验学分册, 1999(6): 274-275.
[38]WOLFFRAM S, BLO..CK M, ADER P. Quercetin-3-glucoside is transported by the glucose carrier SGLT1 across the brush border membrane of rat small intestine[J]. J Nutr, 2002, 132(4): 630-635.
[39]KARBACH U. Paracellular calcium transport across the small intestine [J]. J Nutr, 1992, 122(Suppl 3): 672-677.
[40]GILL S R, POP M, DEBOY R T, et al. Metagenomic analysis of the human distal gut microbiome[J]. Science, 2006, 312: 1355-1359.
[41]FINEGOLD S M, ATTEBERY H R, SUTTER V L. Effect of diet on human fecal flora: comparison of Japanese and American diets[J]. Am J Clin Nutr, 1974, 27(12): 1456-1469.
[42]TURNBAUGH P J, LEY R E, MAHOWALD M A, et al. An obesityassociated gut microbiome with increased capacity for energy harvest[J]. Nature, 2006, 444: 1027-1031.
[43]CANI P D, NEYRINCK A M, FAVA F, et al. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia[J]. Diabetologia, 2007, 50(11): 2374-2383.
[44]BOUHNIK Y, de la FLOURIE B, RIOTTO M, et al, Effects of fructooligosaccharides ingestion on fecal bifidobacteria and selected metabolic indexes of colon carcinogenesis in healthy humans[J]. Nutr Cancer, 1996, 26(1): 21-29.
[45]阎春玲, 刘兵, 刘万顺. 壳寡糖及其衍生物对糖尿病大鼠糖耐量及肠道微生态平衡的影响[J]. 世界华人消化杂志, 2007, 15(11): 1202-1207.
[46]LEE H C, JENNER A M, LOW C S, et al. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota[J]. Research in Microbiology, 2006, 157(9): 876-884.
[47]TURNBAUGH P J, HAMADY M, YATSUNENKO T, et al. A core gut microbiome in obese and lean twins[J]. Nature, 2009, 457: 480-484.
[48]MACHFARLANE G T, GIBSON G R, BEATTY E, et al. Estimation of short-chain fatty acid production from protein by human intestinal bacteria based on branched-chain fatty acid measurements[J]. FEMS Microbiology Letters, 1992, 101(2): 81-88.
[49]MACHFARLANE G T, ALLISON C. Utilisation of protein by human gut bacteria[J]. FEMS Microbiol Ecol, 1986, 38(1): 19-24.
[50]CASTERLINE J L J, OLES C J, KU Y. in vitro fermentation of various food fiber fractions[J]. Journal of Agricultural and Food Chemistry, 1997, 45(7): 2463-2467.
[51]MOREAU N M, MARTIN L J, TOQUET C S, et al. Restoration of the integrity of rat caeco-colonic mucosa by resistant starch, but not by fructo-oligosaccharides, in dextran sulfate sodium: induced experimental colitis[J]. British Journal of Nutrition, 2003, 90(1): 75-85.
[52]MATRIN F P, DUMAS M E, WANG Yulan, et al. A top-down systems biology view of microbiome-mammalian metabolic interactions in a mouse model[J]. Mol Syst Biol, 2007, 3(112): 112.
[53]GRIFFITHS L A, BARROW A. Metabolism of flavonoids compounds in germ-free rats[J]. Biochem J, 1972, 130(4): 1161-1162.
[54]ROWLAND I R, H WISEMAN H, SANDERS T A B, et al. Interindividual variation in metabolism of soy isoflavones and lignans: influence of habitual diet on equol production by the gut microflora[J]. Nutr Cancer, 2000, 36(1): 27-32.
[55]KARIKURA M, MIYASE T, TANIZAWA H, et al. Studies on absorption, distribution, excretion and metabolism of ginseng saponinis. Ⅶ. Comparison of decomposition models of ginsenoside-Rbl and -Rb2 in the digestive tract of rats[J]. Chem Pharm Bull, 1991, 39(9): 2357-2361.
[56]TSUCHIHASHI R, KONDERA M, SAKAMOTO S, et al. Microbial transformation and bioaetivation of isoflavones from pueraria flowers by human intestinal bacterial strains[J]. J Nat Med, 2009, 63(3): 254-260.
[57]白雪. 葛花苷大鼠体内代谢的研究[D]. 沈阳: 沈阳药科大学, 2010.
[58]DRASAR B S, RENWICK A G, WILLIAMS R T. The role of the gut flora in the metabolism of cyclamate[J]. Biochem J, 1972, 129(4): 881-890.
[59]HILL M J. Intestinal flora and endogenous vitamin synthesis[J]. Eur J Cancer Prev, 1997, 6(Suppl 2): 43-45.
[60]BHATTACHARYYA D K, KWON O, MEGANATHAN R. Vitamin K2(menaquinone) biosynthesis in Escherichia coli: evidence for the presence of an essential histidine residuein O-succinylbenzoyl coenzyme a synthetase[J]. J Bacteriol, 1997, 179(19): 6061-6065.
[61]NICHOLSON J K, HOLMES E, WILSON I D. Gut microorganisms, mammalian metabolism and personalized health care[J]. Nat Rev Microbiol, 2005, 3(5): 431-438.
[62]HEYMAN M, CORTHIER G, LUCAS F, et al, Evolution of the caecal epithelial barrier during Clostridium difficile infection in the mouse[J]. Gut, 1989, 30(8): 1087-1093.
[63]BRUN P, CASTAGLIUOLO I, LEO V D, et al. Increased intestinal permeability in obese mice: new evidence in the pathogenesis of nonalcoholic steatohepatitis[J]. Am J Physiol Gastrointest Liver Physio, 2007, 292(2): G518-G525.
[64]LEE I A, BAE E A, LEE J H, et al. Bifidobacterium longum HY8004 attenuates TNBS-induced colitis by inhibiting lipid peroxidation in mice[J]. Inflammation Research, 2010, 59(5): 359-368.
[65]MATSUMOTO S, HARA T, HORI T, et al. Probiotic Lactobacillusinduced improvement in murine chronic inflammatory bowel disease is associated with the down: regulation of pro-inflammatory cytokines in lamina propria mononuclear cells[J]. Clinical and Experimental Immunology, 2005,140(3): 417-426.
[66]DIELEMAN L A, GOERRES M S, ARENDS A, et al. Lactobacillus GG prevents recurrence of colitis in HLA-B27 transgenic rats after antibiotic treatment[J]. Gut, 2003, 52(3): 370-376.
[67]ZOCCO M A, dal VERME L Z, CREMONINI F, et al. Efficacy of Lactobacillus GG in maintaining remission of ulcerative colitis[J]. Aliment Pharmacol Ther, 2006, 23(11): 1567-1574.
[68]ROUND J L, MAZMANIAN S K. The gut microbiota shapes intestinal immune responses during health and disease[J]. Nat Rev Immunol, 2009, 9(5): 313-324.
[69]MACPHERSON A J, HARRIS N L. Interactions between commensal intestinal bacteria and the immune system[J]. Nature Rev Immunol, 2004, 4(6): 478-485.
[70]FALK P G, HOOPER L V, MIDTVEDT T, et al. Creating and maintaining the gastrointestinal ecosystem: what we know and need to know from gnotobiology[J]. Microbiol Mol Biol Rev, 1998, 62(4): 1157-1170.
[71]BOUSKRA D, BRZILLON C, BRARD M, et al. Lymphoid tissue genesis induced by commensals through NOD1 regulates intestinal homeostasis[J]. Nature, 2008, 456: 507-510.
[72]ABREU M T. Toll-like receptor signaling in the intestinal epithelium: how bacterial recognition shapes intestinal function[J]. Nat Rev Immunol, 2010, 10(2): 131-143.
[73]BEMET M F, BRASSART D, NEESER J R, et al. Adhesion of human bifidobacterial strains to cultured hurnan intestinal epithelial cells and inhibition of enteropathogen-cell interactions[J]. Appl Euviron Mierobiol, 1993, 59(12): 4124-4128.
[74]FOURNIAT J, COLOMBAN C, LIME C, et al. Heat-killed Lactobacillus acidophilus inhibits adhesion of Eseherichia coli B41 to HeLa cells [J]. Ann Rech Vet, 1992, 23(4): 361-370.
[75]NISKANEN L, LAAKSONEN D E, KRISTIINA NYYSSONEN K, et al. Inflammation, abdominal obesity and smoking as predictors of hypertension[J]. Hypertension, 2004, 44(6): 859.
[76]MONTEIRO R, AZEVEDO I. Chronic inflammation in obesity and the metabolic syndrome[J]. Mediators of Inflammation, 2010, 2010: 1-10.
[77]MATARESE G, CAVA L A. The Intricate Interface between immune system and metabolism[J]. Trends in Immunology, 2004, 25(4): 193-200.
[78]HOTAMISLIGIl G S, ERBAY E. Nutrient sensing and inflammation in metabolic diseases[J]. Nat Rev Immunol, 2008, 8(12): 923.
[79]BENSINGER S J, TONTONOZ P. Integration of metabolism and inflammation by lipid-activated nuclear receptors[J]. Nature, 2008, 454: 470-477.
[80]HOTAMISLIGIl G S. Inflammation and metabolic disorders[J]. Nature, 2006, 444: 860-867.
[81]KONNER A C, BRUNING J C. Toll-like receptors: linking inflammation to metabolism[J]. Trends in Endocrinology and Metabolism, 2011, 22(1): 16-23.
[82]WOLOWCZUK I, VERWAERDE C, VILTART O. Feeding our immune system: impact on metabolism[J]. Clinical and Developmental Immunology, 2008, 2008: 639803.
[83]毛炜翔, 高金燕, 陈红兵. 小麦过敏研究进展[J]. 食品科学, 2007, 28 (8): 559-562.
[84]de la SERRE C B, ELLIS C L, LEE J, et al. Propensity to high-fat dietinduced obesity in rats is associated with changes in the gut microbiota and gut inflammation[J]. American Journal of Physiology Gastrointest Liver Physiology, 2010, 299(2): G440-G448.
[85]ANDREASEN A S, KRABBE K S, KROGH-MADSEN R, et al. Human endotoxemia as a model of systemic inflammation[J]. Current Medicinal Chemistry, 2008, 15(17): 1697-1705.
[86]GHANIM H, ABUAYSHEH S, SIA C L, et al. Increase in plasma endotoxin concentrations and the expression of Toll-like receptors and suppressor of cytokine signaling-3 in mononuclear cells after a high-fat, high-carbohydrate meal[J]. Diabetes Care, 2009, 32(12): 2281-2287.
[87]LEEL J Y, HWANG D H. The modulation of inflammatory egne expression by lipids: mediation through Toll-like receptors[J]. Mol Cells, 21, 21(2): 174-185.
[88]TAN Bie, LI Xinguo, KONG Xiangfeng, et al. Dietary L-arginine supplementation enhances the immune status in early-weaned piglets[J]. Amino Acids, 2009, 37(2): 323-331.
[89]LI Peng, YIN Yulong, LI Defa, et al. Amino acids and immune function [J]. British Journal of Nutrition, 2007, 98(2): 237-252.
[90]郑丽坤, 张磊, 陈乃耀. 维甲酸和肠道免疫稳态[J]. 世界华人消化杂志, 2008, 16(8): 885-891.
[91]MUCIDA D, PARK Y, KIM G, et al. Reciprocal TH17 and regulatory T cell differentiation mediated by retinoic acid[J]. Science, 2007, 317: 256-260.
[92]陶月红. 维生素A对树突状细胞的调节及其在黏膜免疫中的作用[D].上海: 复旦大学, 2006.
[93]HSU A, BRUNO S R, LO..HR C V, et al. Dietary soy and tea mitigate chronic inflammation and prostate cancer via NF-κB pathway in the noble rat model[J]. Journal of Nutritional Biochemistry, 2010, 22(5): 502-510.
[94]FRAUWIRTH K A, RILEY J L, HARRIS M H, et al. The CD28 signaling pathway regulates glucose metabolism[J]. Immunity, 2002, 16(6): 769-777.
[95]HEALY D A, WATSON R W, NEWSHOLME P. Glucose, but not glutamine, protects against spontaneous and anti-fasantibody-induced apoptosis in human neutrophils[J]. Clinical Science, 2002, 103(2): 179-189.
[96]NOWELL M A, RICHARDS P J, HORIUCHI S, et al. Soluble IL-6 receptor governs IL-6 activity in experimental arthritis: blockade of arthritis severity by soluble glycoprotein 130[J]. Journal of Immunology, 2003, 171(6): 3202-3209.
[97]PLOMGAARD P, FISCHER C P, IBFELT T, et al. Tumor necrosis factor-α modulates human in vivo lipolysis[J]. Journal of Clinical Endocrinology & Metabolism, 2008, 93(2): 543-549.
[98]DONG Z M, GUTIERREZRAMOS J C, COXON A, et al. A new class of obesity genes encodes leukocyte adhesion receptors[J]. Proc Natl Acad Sci USA, 1997, 94(14): 7526-7530.
[99]TOUFEKTSIAN M C, SALEN P, LAPORTE F, et al. Dietary flavonoids increase plasma very long-chain (n-3) fatty acids in rats[J]. The Journal of Nutrition, 2010, 11(3): 1-5.
[100]董英, 张慧慧. 苦瓜多糖降血糖活性成分的研究[J]. 营养学报, 2008, 30(1): 54-56.
[101]BOSI E, MOLTENI L, RADAELLI M G, et al. Increased intestinal permeability precedes clinical onset of type 1 diabetes[J]. Diabetologia, 2006, 49(12): 2824-2827.
[102]张晗, 寿松涛, 崔书章. 碱性磷酸酶在脓毒症中的临床意义[J]. 世界急危重病医学杂志, 2007, 4(5): 2071-2073.
[103]LI Qiurong, ZHANG Qiang, WANG Meng, et al. n-3 polyunsaturated fatty acids prevent disruption of epithelial barrier function induced by proinflammatory cytokines[J]. Molecular Immunology, 2008, 45(5): 1356-1365
[104]刘玉江. 膳食纤维对实验性结肠炎大鼠肠黏膜屏障的影响[D]. 太原:山西医科大学, 2010.
[105]汤健, 温绍君, 王佐广, 等. 动物与临床试验: 评价大豆低聚肽的降血压效果[J]. 食品与发酵工业, 2006, 32(3): 11-13.
[106]丁志山, 高承贤, 陈铌铍, 等. 姜黄素具有抑制血管生成与诱导肿瘤细胞凋亡双重作用[J]. 中国药理学通报, 2003, 19(2): 171-173.
[107]SIVAGNANAM P, KOUTSOUMPAS A, FORBES A. Respiratory symptoms in patients with inflammatory bowel disease and the impact of dietary salicylates[J]. Digestive and Liver Disease, 2007, 39(3): 232-239.
[108]MAHADEVA R, WALSH G, FLOWER C D, et al. Clinical and radiological characteristics of lung disease in inflammatory bowel disease[J]. Eur Respir, 2000, 15(1): 41-48.
[109]TARHAN O R, CEYLAN B G, KAPUCUOGLU N, et al. Activated protein C and normal saline infusion might prevent deleterious effects of remote acute lung injury caused by intestinal ischemia-reperfusion: an experimental study in the rat model[J]. Journal of Surgical Research. 2011, 168(1): e81-e86.
[110]刘辉. 吸入维生素A与布地奈德对难治性哮喘大鼠肺部病理及TSLP表达的影响[D]. 合肥: 安徽医科大学, 2008.
[111]BISWAS S, RAHMAN I. Modulation of steroid activity in chronic inflammation: a novel anti-inflammatory role for curcumin[J]. Mol Nutr Food Res, 2008, 52(9): 987-994.
[112]KNEKT P, KUMPULAINEN J, JARVINEN R, et al. Flavonoid intake and risk of chronic diseases[J]. Am J Clin Nutr, 2002, 76(3): 560-568.
[113]WALDA I C, TABAK C, SMIT H A, et al. Diet and 20-year chronic obstructive pulmonary disease mortality in middle-aged men from three european countries[J]. Eur J Clin Nutr, 2002, 56(7): 638-643.
[114]MEJA K K, RAJENDRASOZHAN S, ADENUGA D, et al. Curcumin restores corticosteroid function in monocytes exposed to oxidants by maintaining HDAC2[J]. Respir Cell Mol Biol, 2008, 39(3): 312-323.
[115]GRIDER J R, KUEMMERLE J F, JIN J G. 5-HT released by mucosal stimuli initiates peristalsis by activating 5-HT4/5-HT1p receptors on sensory CGRP neurons[J]. Am J Physiol, 1996, 270(5 Pt 1): G778-G782.
[116]SCHMIDT E D, JANSZEN A W, WOUTERLOOD F G, et al. Interleukin-1-induced long-lasting changes in hypothalamic corticotropinreleasing hormone (CRH): neurons and hyperresponsiveness of the hypothalamus-pituitary-adrenal axis[J]. J Neurosci, 1995, 15(11): 7417-7426.
[117]UYSAL K T, WIESBROCK S M, MARINO M W, et al. Protection from obesity-induced insulin resistance in mice lacking TNF-alpha function [J]. Nature, 1997, 389: 610-614.
[118]PLOMGAARD P, NIELSEN R A, FISCHER P C, et al. Associations between insulin resistance and TNF-α in plasma, skeletal muscle and adipose tissue in humans with and without type 2 diabetes[J]. Diabetologia, 2007, 50(12): 2562-2571.
[119]FAROOQI I S, KEOGH J M, GSH Y, et al. Clinical spectrum of obesity and mutations in the melanocortin 4 receptor gene[J]. N Engl J Med, 2003, 348(12): 1085-1095.
[120]BACKHED F, DING Hao, WANG Ting, et al. The gut microbiota as an environmental factor that regulates fat storage[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101 (44): 15718-15723.
[121]HOLMES E, LOO R L, STAMLER J, et al. Human metabolic phenotype diversity and its association with diet and blood pressure[J]. Nature, 2008, 453: 396-401.
The Role of the Gastrointestinal Tract: A Bridge between Food Nutrition and Health
RUAN Zheng1,DENG Ze-yuan1,WU Guo-yao2,YIN Yu-long1,3,*
(1. College of Life Science and Food Engineering, Nanchang University, Nanchang 330031, China;
2. College of Agriculture and Life Sciences, Texas A&M University, Texas 77843-2471, USA;
3. Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China)
Health improvement will become one of the most important challenges for countries in the 21st century. The consumption of healthy foods is the best way for people to prevent chronic disease. In this paper, the interactions among food compositions, microbes and their metabolites in the intestine as well as intestinal structure and functions are summarized. The gastrointestinal tract plays a vital role in whole-body homeostasis, whereas foods can provide nutrients and regulate biological activity to affect the initial events of a phenomenon and the mid-term bidirectional effects.
food;nutrition;health;gastrointestinal tract
TS201.4
A
1002-6630(2012)11-0273-10
2011-05-26
阮征(1978—),男,副教授,博士,研究方向为功能食品与营养代谢调控。E-mail:ezruan@yahoo.com
*通信作者:印遇龙(1956—),男,教授,博士,研究方向为分子营养。E-mail:yyulong2003@yahoo.com.cn
