面向入侵检测的集成人工免疫系统
2012-10-26陈岳兵冯超张权唐朝京
陈岳兵,冯超,张权,唐朝京
(1.总参第六十一研究所,北京 100141;2.国防科技大学 电子科学与工程学院,湖南 长沙 410073)
1 引言
人体免疫系统保护人体免受各种病原体的入侵,为入侵检测系统提供了丰富的启发。研究者应用免疫学原理来设计智能计算范例,逐渐发展成一个新的研究领域,称为人工免疫系统[1]。研究者分别基于否定选择机制和危险模型[2]提出了 NSA和DCA[3]。2种算法都可以用于入侵检测,但前者可用性欠佳,后者检测性能偏低,在应用方面都有局限性。
在入侵检测研究中,追求高检测率、低误报率[4]和快速的检测速度。根据检测方法不同,分为误用检测和异常检测。误用检测又称为特征检测,它是通过对已知入侵的研究,提取攻击的特征形成特征集合,利用这些特征集合对当前的数据进行各种处理后,再进行特征匹配工作,如果匹配成功,则判定为入侵。该方法能够很好地检测已知的入侵,具有检测率高、误报率低和检测速度快等特点,缺点是它不能检测未知的入侵。当有未知的入侵出现的时候,模式库必须随之更新,给系统的维护管理带来困难。异常检测又称为基于行为的检测,它是利用与系统行为相关的一些统计量来构造正常的行为模式[5],如果待检测行为偏离正常行为,则判定为入侵。该方法存在统计量选取困难、异常阈值确定困难、误报率较高的问题,但是它能对未知的入侵进行检测。
本文对危险模型进行了分析,指出否定选择机制和危险模型两者之间并不矛盾:在危险模型中,否定选择机制和危险性判别机制在不同的位置发挥各自的作用,两者协作完成检测和应答。基于这种新的认知,结合实时DCA和NSA构建了一个集成人工免疫系统(IAIS),用于入侵检测。实时DCA检测行为特征,NSA检测结构特征,构建的 IAIS可以同时利用结构特征和行为特征进行入侵检测。通过入侵检测基准数据集对 IAIS进行验证,并与其他方法进行了比较。IAIS检测性能与经典分类算法相当,且具有良好的可用性。
2 启发
在免疫学中,感染非我模型[6]和危险模型[2]是2种主要的免疫学模型,两者都有各自的支持者和部分证据,都是具有一定合理性的解释。表面上看,2种模型互相矛盾,但是危险模型的提出者Matzinger从未明言感染非我模型是错误的,也没有讲否定选择是不正确的。Matzinger认为否定选择是一种避免自我免疫的前置条件,只是在免疫应答之前还需要进一步的确认机制以消灭有害抗原。危险模型在图 1中给出,图中 APC(antigen presenting cells)、B、Tk和Th分别表示抗原提呈细胞、B细胞、毒性T细胞和辅助T细胞。APC属于固有免疫系统;B细胞、毒性T细胞和辅助T细胞属于适应性免疫系统。图中共有3类信号:抗原和B细胞受体,毒性T细胞以及APC之间的作用产生信号1;辅助T细胞和APC释放信号2;身体组织的正常细胞在受到损害的时候释放信号 0,或危险信号,用于激活本地APC。感染非我模型和危险模型的唯一区别在信号0。在感染非我模型中,APC通过模式识别受体常见的非我分子,产生刺激信号。而在危险模型中,APC感知组织中受损细胞释放的的危险信号。
感染非我模型和危险模型都包括抗原与B细胞以及毒性T细胞之间的结构匹配。虽然2种模型不可能同时正确,这并不表示这两者完全不相容。实际上,否定选择和自我/非我模型之间不能划等号。在图1中,否定选择是作为危险模型中的部分机制存在的,否定选择和危险性判别都是必要的。据此,提出结合危险性判别机制和否定选择机制,建立集成系统。否定选择机制负责检测结构特征,危险性判别机制负责检测行为特征。
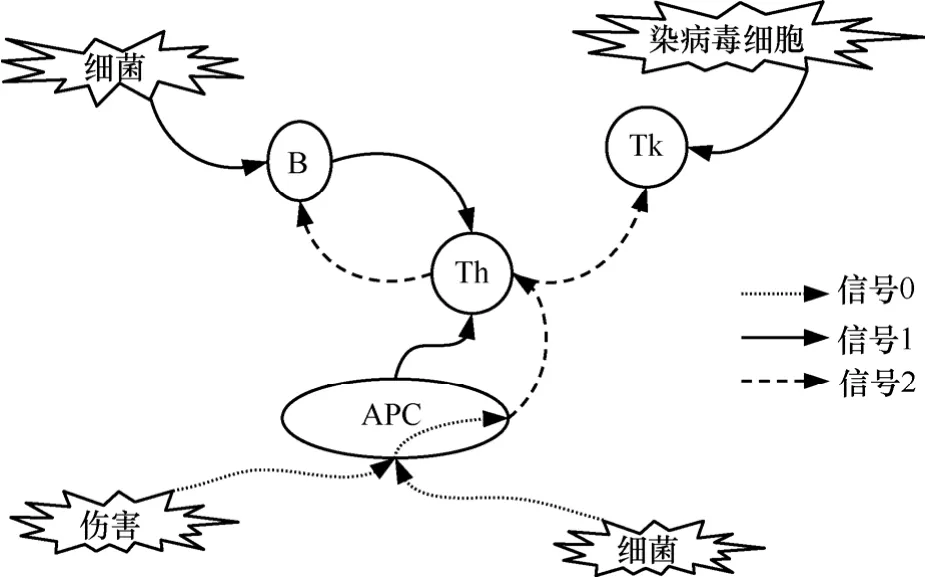
图1 危险模型
3 集成人工免疫系统
本节根据对免疫系统的新的认识,提出结合DCA和NSA建立IAIS,描述了系统结构及其实现,给出了系统的使用方法。
3.1 系统概述
IAIS总体结构如图2所示,系统基本组件及其功能描述如表1所示。IAIS集成的2个算法分别为实时DCA和二进制表示的NSA。
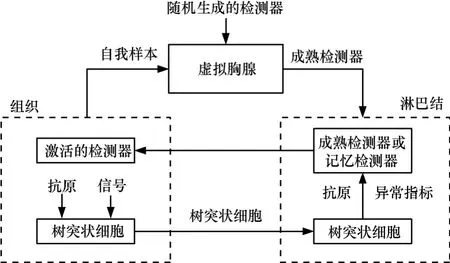
图2 IAIS结构
IAIS的功能是检测目标系统中的异常实体,如图2所示。组织代表目标系统;抗原是组织中运行的实体;信号是与组织运行状态相关的观测值;树突状细胞负责将最新的抗原和信号从组织传送到淋巴结;淋巴结是一个分析中心,提呈的抗原和信号在此经过分析之后得到代表抗原异常程度的指标;虚拟胸腺负责生成成熟检测器。淋巴结中分析得到的异常指标用于辅助成熟检测器和记忆检测器对抗原的检测。

表1 IAIS组件

算法1描述了IAIS的运行过程。系统根据组织的当前状态维持一个动态的自我集合,组织状态异常时出现的抗原被禁止加入自我集合。然后检查NSA中记忆检测器和普通检测器的生命周期,清除过期检测器,生成新的检测器以填补空缺。实时DCA对输入抗原和信号进行处理以得到不同抗原类型的动态异常指标。
在 IAIS中,提出结合抗原动态异常指标计算检测器和抗原之间的匹配阈值,如式(1)所示。通过汉明距离计算检测器和抗原之间的亲和度并与动态匹配阈值进行比较,亲和度在匹配阈值之上的判为异常,反之判为正常。此过程连续运行,不断处理新输入数据。
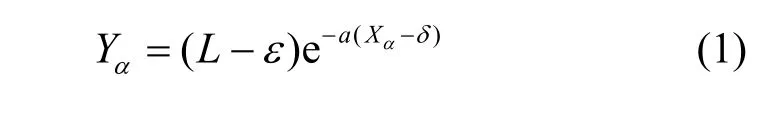
其中,Xα是抗原类型α的动态异常指标,Yα是此类抗原的匹配阈值,a是常数,δ是实时DCA中的异常阈值,L是检测器长度,ε是自我半径[8],它决定检测器在二进制空间的覆盖范围。对记忆检测器取更大的a值以降低器匹配阈值。根据式(1),抗原类型的异常程度越高,匹配阈值越低;反之,抗原类型的异常程度越低,匹配阈值越高。
3.2 系统实现
本节描述了实时DCA和NSA实现。
1) 实时DCA
实时DCA的基本功能是接收输入信号和抗原,输出每一类抗原的动态异常指标。多路信号源作为算法输入,分为 PAMP (pathogen-associated molecular pattern)、危险信号(danger signal)、安全信号(safe signal)和炎性信号(inflammation signal),各信号的含义如下。
PAMP:表明出现异常行为,此信号增强与高置信度异常行为的存在密切相关。
危险信号:表明可能存在异常行为,此信号增强表明异常行为存在的可能性增加。
安全信号:表明有正常行为发生,此信号增强表明正常行为的可能性增加。安全信号用于抵消PAMP和危险信号的影响。
炎性信号:表明总体状态异常。炎性信号用于放大其他信号。
实时DCA通过式(2)处理输入信号得到输出信号。输出信号包括协同刺激信号(csm)、半成熟信号(semi)和成熟信号(mat)。
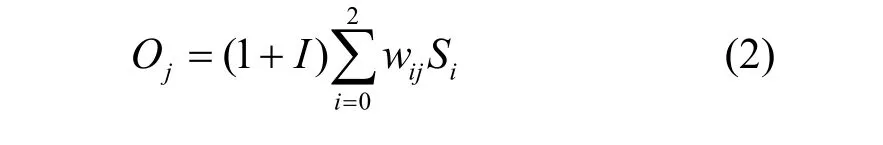
其中,Si是输入信号,Oj是输出信号,I是炎性信号,wij是从Si到Oj的转换权值。
实时DCA是在DCA[9]基础上进行改进提出的,主要包括2点改进。第一,实时DCA中增加了树突状细胞在淋巴结中的生命周期。为简化起见,让树突状细胞在淋巴结中存活一个系统周期,执行抗原提呈;第二,实时 DCA提出了动态异常指标,在 MAC[9]基础上提出动态 MAC(Cα),如式(3)所示。

其中,α是所有具有相同值的抗原结合,mα是抗原类型α被提呈为成熟抗原的数量,Ai是抗原类型i被提呈的总数,A是抗原类型总数。mα和Ai是根据当前淋巴结中存活的树突状细胞统计得到的,过期的树突状细胞不在统计之列。Cα在0到1之间,Cα越靠近1,表明此类抗原异常程度越高。
实时DCA伪代码如算法2所示。组织和T细胞种群被实现为2个串行子系统,树突状细胞负责将信息从组织传送到淋巴结,T细胞种群对信息进行分析。

计算并记录抗原类型及对应的动态异常指标
在组织中,首先初始化树突状细胞种群,为每个树突状细胞赋予一个迁移阈值,用于决定其在组织中的生命周期。迁移阈值是指定范围内的随机数,这使得实时DCA成为一个随机系统,自然地,IAIS也是一个随机系统。当有输入数据的时候,组织处理数据得到信号和抗原,然后组织中每个树突状细胞经历一个累积信号并走向成熟过程[9]。与DCA不同的是,实时DCA中达到迁移阈值的树突状细胞要迁移到淋巴结中,并执行抗原提呈。
T细胞种群对淋巴结中的树突状细胞进行实时分析,并生成异常阈值的时间序列,逐代记录与抗原类型对应的动态异常指标。
2) NSA
实现的 NSA采用二进制表示法,算法包括 2个阶段:检测器生成阶段和检测阶段。检测器生成阶段如算法3所示,随机生成候选检测器,匹配自我样本的候选检测器被清除,否则加入检测器集合,当检测器数目达到预设值的时候算法终止。

在检测阶段,计算输入样本和所有检测器的亲和度,如果样本在某一个检测器的自我半径之内,则将此样本判为异常。
3.3 使用过程
在实验之前,首先介绍如何使用IAIS,其一般过程包括4个部分。
1) 定义抗原:抗原就是目标系统中运行的实体。IAIS需要身份标识来表示这些实体,通常用数字和字符串形式的唯一标识符表示一个抗原或者一类抗原。
2) 抽取信号:通过各种信号表示系统状态。用户需要根据先验知识和分析抽取有用信号来代表整个系统状态,包括PAMP、危险信号、安全信号和炎性信号。
3) 数据预处理:将原始信号转化为无量纲的数字,再对每类信号进行归一化处理,例如,将指定信号区间映射到[0,100],常用的归一化函数包括线性函数,阶梯函数和Sigmoid函数,最后将抗原流和信号流提交给IAIS。
4) 数据处理:IAIS处理提交的抗原流和信号流,并输出每类抗原的动态异常指标。
4 实验测试
在KDD99数据集上进行实验,研究IAIS的属性,并通过与其他方法的比较来评估系统性能。
4.1 测试集
KDD99数据集是一个用于入侵检测领域的基准数据集。实验采用10%子集,它包括494021个数据项(实例或样本)。10%子集与完整数据集具有类似统计特性,保持了类似的正常连接和攻击比。KDD99数据集本身没有时间戳,为了测试 IAIS,每秒采样10个抗原来模拟实际情况,在数据集中加入时间戳,假设数据集以1s为间隔采集。KDD99数据集的数据项是41维(特征或属性)向量。根据文献[10]介绍的数据域含义,对数据集进行预处理,得到抗原和信号。抗原代表结构特征,信号代表行为特征。
1) 抗原
将选择的数据域转换为 24bit的二进制串,用来表示抗原,如表2所示。
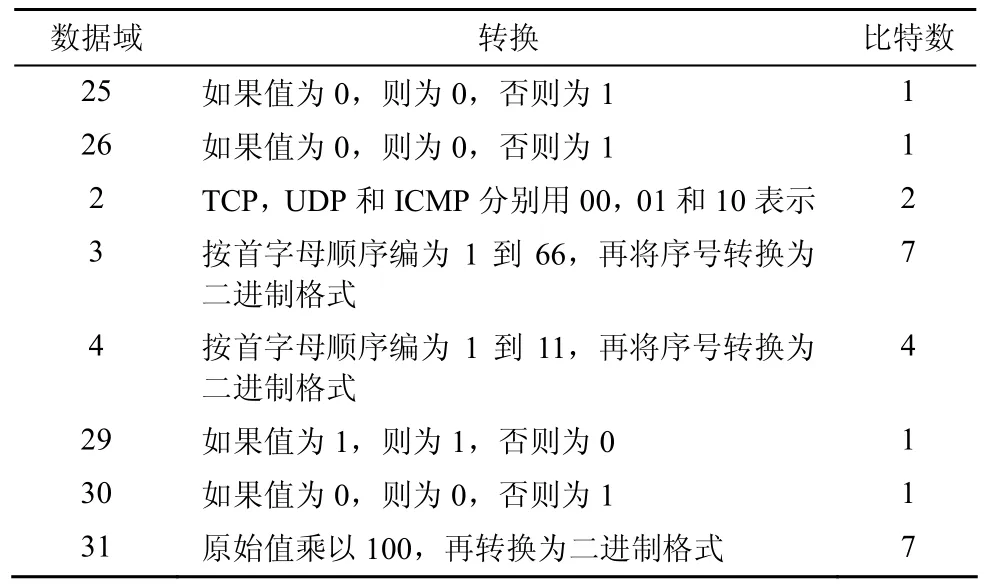
表2 抗原构造
2) 信号
选择作为信号的数据域与文献[11]相同。10个数据域分为3类(不包括炎性信号)。
PAMP:数据域25、26、29、38和40。
危险信号:数据域23和24。
安全信号:数据域12、31和32。
设x为数据域的值,如果x∈[m,n]时表现异常,这个域为PAMP或者危险信号;如果表现正常,则为正常信号。将这些数据域的指定区间按照式(4)归一化到[0,100]。
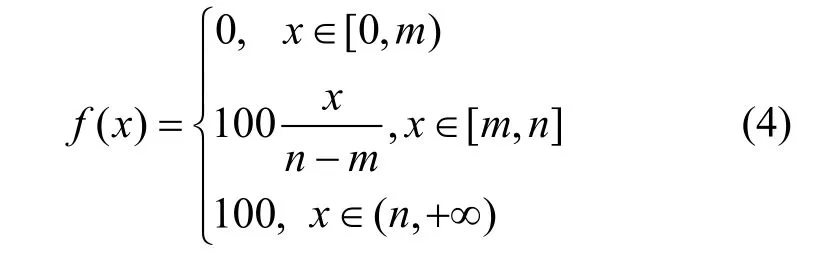
对数据域12,感兴趣的指定区间为[0, 0.99],其他数据域的指定区间按照[min, max]构造。每类信号的均值作为此类信号的值。
4.2 实验设置
IAIS代码通过MATLAB R2009a实现,所有实验在Windows 7(Intel Pentium Dual CPU T2330,4GB RAM)平台下运行。除非特别指出,实验使用表3中给出的参数。树突状细胞数量的选择是精度和代价之间的平衡,考虑计算开销,设为10;细胞周期率(cell cycle rate)根据数据集中信号采样率设置,设为1采样/s;检测器长度(L)为抗原位数,L=24。实验对系统性能有显著影响的主要参数进行了敏感性分析。信号处理通过式(2)进行,权值与DCA[3]中使用的一样,如表4所示。为了模拟实时运行环境,通过一段代码读取数据集并提交给IAIS。因为IAIS是随机系统,每个实验运行10次,结果取平均值。利用ROC曲线来评估IAIS的性能。
实验分为3部分进行。
E1:自我半径实验。
E2:异常阈值实验。
E3:与其他方法的比较。
E1,E2使用10%子集的前10000个项,记为S1;E3使用完整10%子集,记为SC。
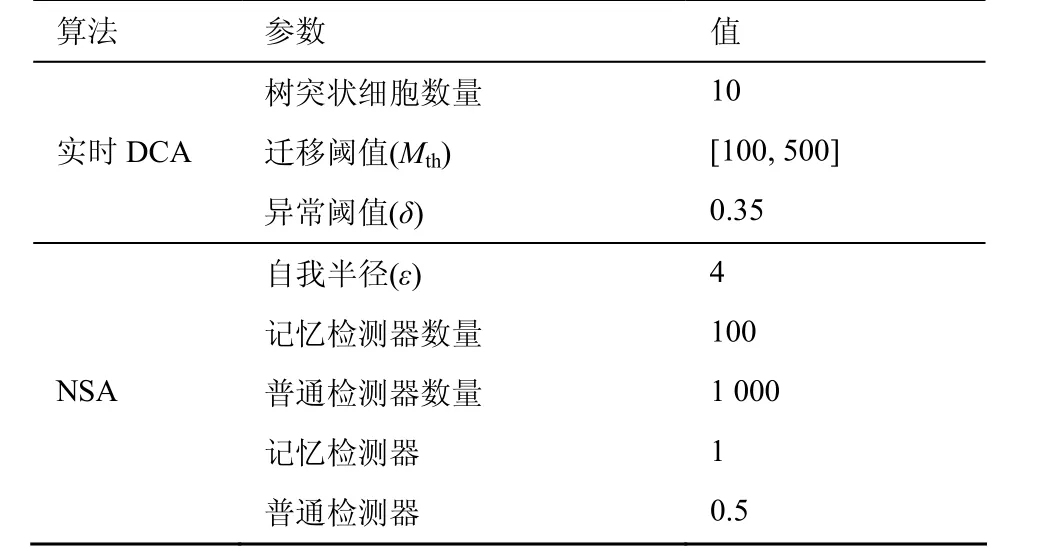
表3 IAIS参数

表4 式(2)推荐权值
5 结果和分析
5.1 E1:自我半径实验
自我半径(ε)决定单个检测器的覆盖范围,如果ε=0,则要求检测器和抗原完美匹配,即结构完全相同。当检测器和抗原之间的亲和度大于L−ε时,认为两者匹配。为研究自我半径对 IAIS性能的影响,在子集S1上测试了如下自我半径ε = 0, 1, 2, …,12。图3给出改变自我半径时的ROC曲线。
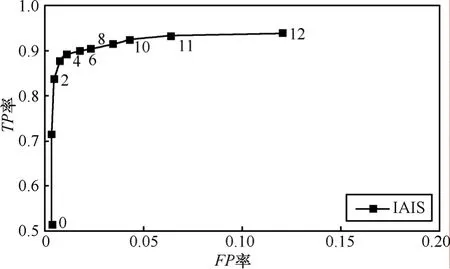
图3 E1的ROC曲线
由图可知,自我半径的取值对TP率和FP率有影响,随着自我半径增加,TP率和FP率都随之增加。ROC曲线上越靠近左上角表示性能越好,即TP率高,FP率低。在TP率和FP率之间进行平衡,下面的实验将自我半径设为ε=4。
5.2 E2:异常阈值实验
异常阈值(δ)用于决定抗原是正常还是异常,动态MAC值高于异常阈值的抗原认为是异常抗原。在子集 S1上应用不同的异常阈值来计算检测率和误报率,测试的异常阈值包括δ=0.1,0.2,…,1,实验结果如表5所示。
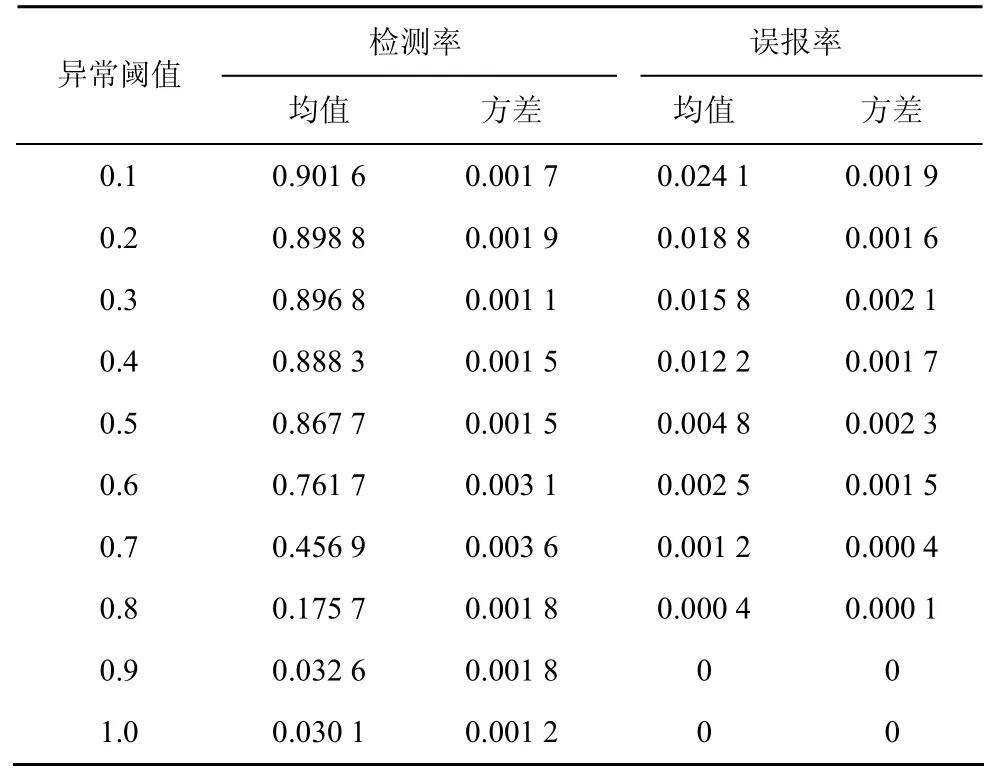
表5 E2的检测率和误报率
表中给出的检测率和误报率的方差相对较小,表明算法性能稳定。从均值来看,异常阈值的取值对 IAIS的性能具有显著的影响,高异常阈值导致低检测率和低误报率。实际应用的时候要在检测率和误报率之间进行平衡。当异常阈值介于0.3和0.4之间时,检测率介于0.8968和0.8883之间,误报率介于0.0158和0.0122之间。下面的实验将异常阈值设为δ=0.35。
5.3 E3:比较
在数据集SC上进行实验,测试IAIS的性能,并与其他方法进行比较,如表6所示。IAIS的实验结果是10次实验的平均值。表中包括NSA和DCA的实验结果,此外还列出了其他智能方法的实验结果,包括人工神经网络,进化计算,支持向量机等。表中各种方法具有各自的特性,实验设置不尽相同,具体细节可参见各种方法的相关文献。
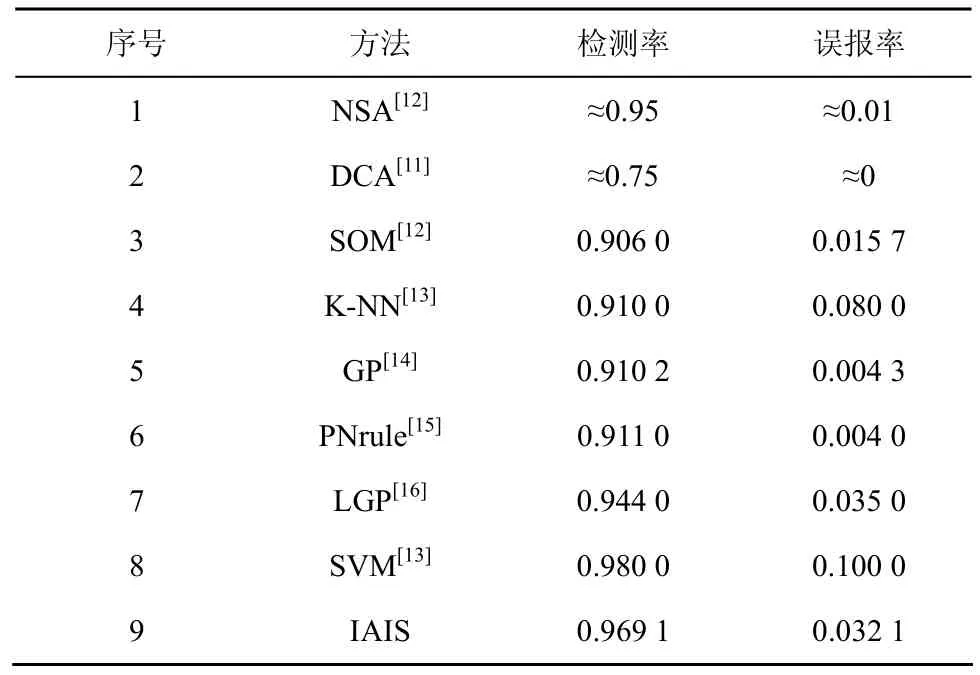
表6 KDD99数据集上的性能比较
首先,将IAIS与NSA和DCA进行比较。与DCA进行比较,IAIS 的检测率比 DCA有明显提高,但与此同时,误报率也提高了,但是幅度不大。相对而言,DCA的检测率太低,因此IAIS更可取。与 NSA进行比较,IAIS的检测率和误报率都比NSA高,但幅度有限。需要指出的是,文献[12]中NSA有个训练过程,需随机选择80%的正常样本进行训练,剩余20%的正常样本和异常样本一起作为测试集。而 IAIS 定义各种信号之后,不需要训练集来训练检测器,训练检测器的自我集合是根据危险信号在线生成的,也就是说,IAIS 可以以实时或近实时的模式运行。
其次,将IAIS 与其他方法进行比较。IAIS 具有次于SVM 的检测率,但是SVM误报率太高,综合检测率和误报率,IAIS性能更好。IAIS误报率与 LGP 相当,检测率比后者高。其他方法虽然误报率低,检测率比IAIS 要低超过5个百分点。通过比较可知,IAIS具有良好的综合性能。
6 结束语
本文对危险模型进行了分析,根据新的关于危险模型和否定选择机制之间关系的理解,提出结合实时DCA和NSA来建立IAIS,用于执行实时入侵检测。系统中实时DCA利用行为特征,NSA利用结构特征。在KDD99数据集上对IAIS进行验证,并与其他方法进行比较。IAIS检测性能与经典分类算法相当,其特点包括:可结合结构特征和行为特征进行检测,不依赖明确标识的数据集来训练检测器,以实时或者近实时的模式运行。
当前 IAIS参数较多,文中仅对主要参数进行了敏感性分析。为了简化起见,部分参数设为常数。人工免疫系统的优点之一就是它能够动态地维持细胞种群数量,因此有待改进,后续工作中可以引入反馈机制让参数根据外部环境的变化进行自适应调整。
[1]DASGUPTA D.Advances in artificial immune systems[J].Theoretical Computer Science, 2006, 403(1):11-32.
[2]MATZINGER P.Tolerance, danger and the extended family[J].Annual Review of Immunology, 1994, 12:991-1045.
[3]GREENSMITH J, AICKELIN U, CAYZER S.Introducing dendritic cells as a novel immune-inspired algorithm for anomaly detection[A].International Conference on Artificial Immune Systems[C].Banff,Canada, 2005.153-167.
[4]LI M.An approach to reliably identifying signs of DDOS flood attacks based on LRD traffic pattern recognition[J].Computers & Security,2004, 23(7):549-558.
[5]LI M.Change trend of averaged hurst parameter of traffic under DDOS flood attacks[J].Computers & Security, 2006, 25 (3):213-220.
[6]JANEWAY C.The immune system evolved to discriminate infectious nonself from non-infectious self[J].Immunology Today, 1992,13:11-16.
[7]SARAFIJANOVIĆ S, BOUDEC J L.An artificial immune system for misbehavior detection in mobile ad-hoc networks with virtual thymus,clustering, danger signal, and memory detectors[A].International Conference on Artificial Immune Systems[C].Catania, Italy,2004.342-356.
[8]CHEN Y B, FENG C, ZHANG Q, et al.Negative selection algorithm with variable-sized r-contiguous matching rule[A].International Conference on Progress in Informatics and Computing[C].Shanghai,China, 2010.150-154.
[9]GREENSMITH J.The Dendritic Cell Algorithm[D].Nottingham, UK:School of Computer Science, University of Nottingham, 2007.
[10]KAYACIK H G, ZINCIR-HEYWOOD A N, HEYWOOD M I.Selecting features for intrusion detection:a feature relevance analysis on KDD 99 intrusion detection datasets[A].Third Annual Conference on Privacy, Security and Trust[C].New Brunswick,Canada, 2005.
[11]GU F, GREENSMITH J, AICKELIN U.Further exploration of the dendritic cell algorithm[A].International Conference on Artificial Immune Systems[C].Phuket, Thailand, 2008.142-153.
[12]GONZÁLEZ F A, DASGUPTA D.Anomaly detection using real-valued negative selection[J].Genetic Programming and Evolvable Machine, 2003, 4(4):383-403.
[13]ESKIN E, ARNOLD A, PRERAU M, et al.Applications of Data Mining in Computer Security[M].Boston, Kluwer, 2002.
[14]FOLINO G, PIZZUTI C, SPEZZANO G.GP ensemble for distributed intrusion detection systems[A].Third International Conference on Advances in Pattern Recognition[C].Bath, UK, 2005.54-62.
[15]AGARWAL R, JOSHI M V.PNrule:a new framework for learning classifier models in data mining (a case-study in network intrusion detection)[A].First SIAM Conference on Data Mining[C].Chicago,2001.1-17.
[16]SONG D, HEYWOOD M I, ZINCIR-HEYWOOD A N.Training genetic programming on half a million patterns:an example from anomaly detection[J].IEEE Trans on Evolutionary Computation,2005,9(3):225-239.
